苦瓜葉片葉綠素響應白粉病菌侵染的遺傳分析
2022-10-17 05:48:30田麗波陳虹容范晨偉郭雪松
熱帶作物學報 2022年9期
關鍵詞:模型
楊 怡,田麗波*,商 桑,陳虹容,范晨偉,郭雪松
苦瓜葉片葉綠素響應白粉病菌侵染的遺傳分析
楊 怡1,田麗波1*,商 桑2*,陳虹容1,范晨偉1,郭雪松1
1. 海南大學園藝學院/海南省熱帶園藝作物品質調控重點實驗室,海南海口 570228;2. 海南大學生命科學學院,海南海口 570228
為了探索苦瓜苗期葉片葉綠素a、葉綠素b以及總葉綠素響應白粉病菌侵染的遺傳規律,以苦瓜高代自交系09-a與09-b為親本構建的170個F2個單株和140個F2:3家系作為材料,利用植物數量遺傳的主基因+多基因混合遺傳模型分別對2個地點的2個群體進行基于單個分離世代群體的遺傳分析。結果表明,白粉病菌脅迫下葉綠素a、葉綠素b以及總葉綠素含量均會降低,感病品系降低幅度顯著大于抗病品系,降解量與病情指數呈正相關,且降解量在2個群體中均呈正態分布。F2和F2:3群體中葉綠素a和葉綠素b的降解量均由一對加性主基因決定,2個群體控制葉綠素a降解量的1對主基因加性效應分別為0.3151、0.1670,控制葉綠素b降解量的1對主基因加性效應分別為0.1333、0.1670。總葉綠素降解量受2對加性-顯性主基因控制,2對主基因在F2群體中加性效應分別為0.2601、0.3184,顯性效應為?0.3452、?0.2221,在F2:3群體加性效應分別為0.1892、0.1624,顯性效應分別為?0.0465、?0.0897,2對主基因的加性和顯性效應共同發揮著重要作用。3個性狀的加性效應均為正值,且控制總葉綠素降解量的2對主基因均以加性效應為主。F2和F2:3群體中葉綠素a降解量主基因遺傳率為76.40%、62.68%。葉綠素b降解量主基因遺傳率分別為72.99%、62.86%,控制葉綠素降解量的2對主基因遺傳率為65.25%、95.36%,遺傳率均大于60%,表明白粉菌脅迫下葉綠素a、葉綠素b及總葉綠素的降解主要受主基因的控制,但也受部分環境的影響。在苦瓜白粉病抗性育種實踐中,可將葉綠素相關性狀的降低量結合病情指數在早期世代進行選擇。本研究為探索苦瓜白粉病抗病機理和抗病育種提供了理論依據。
苦瓜;葉綠素;遺傳分析;主基因+多基因模型
苦瓜(L.)俗名癩瓜、涼瓜,是葫蘆科具有重要經濟價值的蔬菜。起源于印度,在亞洲和非洲國家廣泛種植[1]。其果實富含β-胡蘿卜素、維生素C、葉酸、鎂、磷和鉀等營養物質[2],具有抗氧化、消炎、抗癌、抗糖尿病、抗菌、抗肥胖和免疫調節的功效[3]。因其含有預防疾病和促進健康的植物化合物,已被用于各種中醫治療中。近年來,隨著人們對苦瓜營養價值及諸多藥用功效的深刻認識,栽培面積也逐年擴大。而白粉病是苦瓜設施與露地栽培過程中普遍發生的真菌性病害之一,多發生在亞熱帶和熱帶地區[4-8]。主要為害葉片、葉柄以及莖蔓等,具有潛育期短、傳播速度快、周年可發生等特點[9-11],嚴重影響其產量和品質。殺菌劑是防治白粉病的主要措施之一。然而,藥劑防控不僅危害健康還污染環境,因此,培育抗或耐白粉病品種是一種理想的病害控制策略。
病原菌入侵會使植物寄主發生一系列復雜的生理生化變化,許多病原菌尤其是危害植物葉片的病原菌侵染植物后通常會與葉綠體發生互作,從而導致葉綠體的降解,葉片出現褪綠、黃化、早衰等癥狀,進而影響葉片的光合能力,造成大量減產[11-12]。因此葉片葉綠素含量的高低往往能客觀反映植物抗病性的強弱[13]。MORADI等[14]通過對黃瓜抗感白粉病品種的葉片葉綠素熒光發射光譜分析表明,感病品種受侵染后葉片光合器官受損,但對抗病品種的影響不顯著,葉綠素熒光發射分析可為白粉病的早期檢測和植物種質資源的篩選和分類提供依據。此外,AZMAT等[15]認為豌豆葉表面氣孔密度、葉綠素a和b、總葉綠素濃度與感病嚴重程度顯著相關,可作為豌豆抗白粉病篩選的標準。因此掌握感病后葉綠素含量的遺傳規律,有利于抗病基因的定位和抗病品種的選育,實現農藥減施,對綠色、高效、安全的農業可持續發展具有重要意義。
目前葉綠素突變體的遺傳機理研究較為廣泛,大都屬于質量性狀,如大豆黃綠葉突變體、水稻黃葉突變體、以及擬南芥黃化突變體均受1對隱性核基因控制[16-19]。而大部分研究表明葉綠素含量屬于數量性狀,主要受核基因控制,具有較高的遺傳力[20]。傳統的質量性狀分析法存在把控制性狀遺傳的基因視為一個整體,且無法闡明單個基因的行為和效應等缺點。數量性狀遺傳的基本模型為主基因與多基因共同控制,即可采用主基因+多基因混合遺傳分析方法[21]。該方法把孟德爾分離分析法合理地融合在數量性狀遺傳分析研究中,給出了數量性狀基因體系及其效應的最佳估計,將2對或2對以上的包括上位性和連鎖等情況考慮在內,使植物數量性狀的遺傳分析更深入、準確[22]。目前該方法已經在大豆[23]、玉米[24]、不結球白菜[25]、烤煙[26]以及苦瓜[27]等多種植物葉綠素含量遺傳分析上廣泛應用。然而在葉綠素響應逆境脅迫的遺傳機理方面研究較少,在苦瓜上的研究則更少。本研究利用主基因+多基因混合遺傳分析法對白粉病菌侵染苦瓜后葉綠素相關性狀降解量進行遺傳分析,探討葉綠素響應白粉病的遺傳規律,為提高作物產量和探索抗病機理提供理論依據。
1 材料與方法
1.1 材料
供試材料:以純合的苦瓜高代自交系09-a(P1)和09-b(P2)為親本,均在2015年鑒定其對白粉病的抗性[28]。09-a為斯里蘭卡的野生苦瓜,對白粉病的抗性表現為抗病,在接種白粉菌后感病葉葉綠素降解量低,09-b從廣東收集獲取,對白粉病的抗性表現為感病,在接種白粉菌后感病葉葉綠素降解量高。利用母本自交系(09-a)和父本自交系(09-b)組合配置雜交一代F1及自交二代F2,同時F2通過單粒傳獲得F2:3家系。供試白粉病菌為單囊殼白粉菌(),生理小種2F,由海南大學熱帶瓜類團隊分離純化并鑒定。
試驗儀器:多樣品組織研磨機(型號:Tissuelyser-48;產地:上海);紫外可見分光廣度計(型號:T6新世紀;產地:北京)。
1.2 方法
1.2.1 試驗設計 將苦瓜種子溫湯浸種消毒后,置于覆有雙層濕潤濾紙的培養皿中,并放置在28℃的恒溫培養箱中催芽,露白后單粒播種于10 cm×10 cm的營養缽中。栽培基質按泥炭土∶珍珠巖∶有機肥=6∶3∶1的比例配制,在第3片真葉展開時將幼苗從育苗室移至田間。分別于2019年11月在海口市海南大學園藝學院試驗基地定植苦瓜F2170個單株,2021年1月在瓊海市中原基地定植苦瓜140個F2:3株系,2個群體實驗分別在海口和瓊海2個環境中完成。采用隨機區組設計的方法F2:3每株系定植5株,重復2次,株行距分別為0.5 m和0.8 m。
取感白粉病嚴重的苦瓜葉片,用毛筆刷取葉片上的孢子于配制好的濃度為0.05%的Tween-20溶液中,攪拌均勻獲得孢子懸浮液。再用血球計數板在顯微鏡下觀察并計數分生孢子數,使接種液濃度為每毫升1.0×106個孢子。當植株長至6葉1心時,用小型手持噴霧器將上述孢子懸浮液均勻噴灑在6片真葉表面且呈水珠而不滴落的狀態。
1.2.2 白粉病抗性鑒定 在接種白粉病菌26 d后調查發病情況,進行白粉病抗性鑒定,病情級別的劃定參考SAKATA等[29]和粟建文等[30]的方法,分級標準如表1。
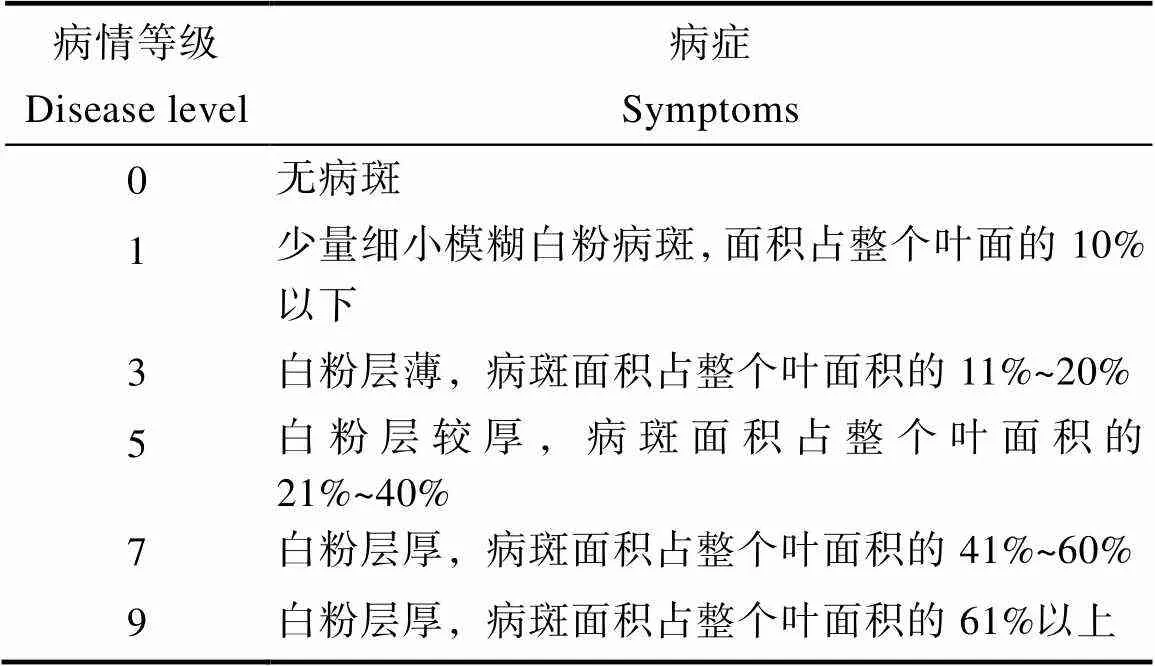
表1 病情等級
病情指數(disease index, DI)計算如下:DI=∑(各病級葉數×相對級數值)/(調查葉總數×9)× 100。植株的抗性鑒定結果依據病情指數分為5級(表2)。
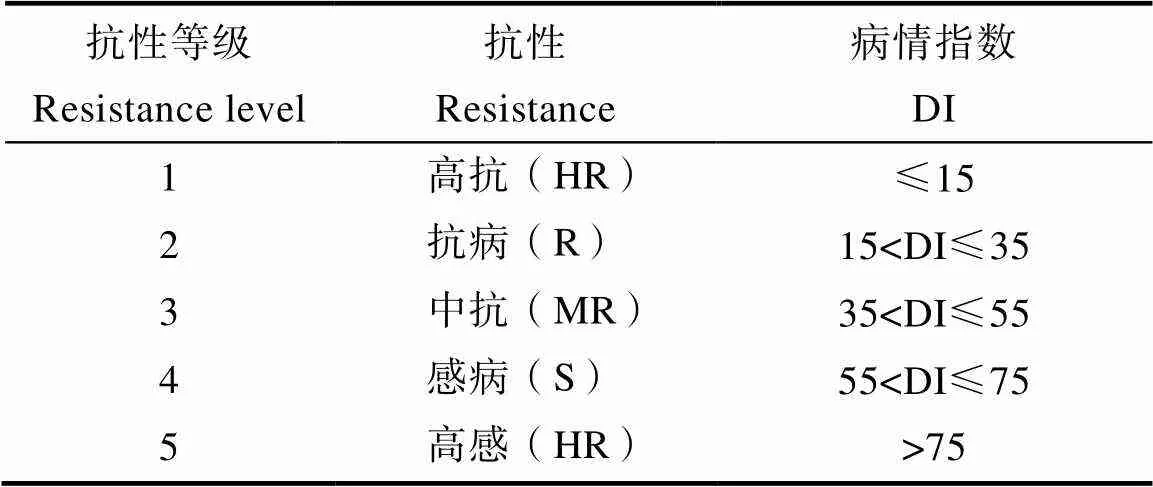
表2 抗性等級
1.2.3 葉綠素含量的測定 葉綠素含量的測定參照李合生[31]的方法,略作改動。接種白粉病菌26 d后達到發病高峰期,在同一植株上分別取發病的第6片真葉和1片未發病的功能葉,將新鮮的葉片剪碎混勻,取0.08 g置于2 mL離心管中,加入0.5 mL 95%乙醇,再加入2顆直徑4 mm和1顆直徑為3 mm的鋼珠,放入研磨機中(50 HZ 3 min)充分研磨。研磨后離心10 min,5000 r/min,將上清液轉入10 mL的棕色容量瓶中,加95%乙醇定容至10 mL。將葉綠素提取液倒入光徑1 cm的比色杯內,分別在665 nm(葉綠素a)、649 nm(葉綠素b)波長下測定吸光值(),根據以下公式分別計算出葉綠素a、葉綠素b、總葉綠素的含量,再計算正常功能葉和發病葉的含量差值。
a=13.95665–6.88649
b=24.96649–7.32665
總葉綠素含量(mg/g)=(××)/(×1000)
式中:為色素含量(mg/L);為提取液體積(mL);為稀釋倍數;為樣品鮮質量(g)。
1.3 數據處理
采用SPSS.19以及Excel 2019軟件對數據進行統計分析和親本間差異顯著性分析,分別根據蓋鈞鎰[21]提出的單個分離世代群體的遺傳模型方法即主基因+多基因混合遺傳模型分離分析法,進行F2和F2:3世代遺傳分析。通過極大似然法對混合分布中的有關參數作出估計,選擇AIC值最小或較小的幾個模型作為最佳模型和備選模型,再根據均勻性12、22、32檢驗、Smironv檢驗和Kolmogorov檢驗等適合性檢驗選出最佳模型。由此估計出混合分布中各成分分布參數、一階和二階遺傳參數。
2 結果與分析
2.1 白粉病菌脅迫下葉綠素相關性狀特征及次數分布
如圖1和圖2所示,高感親本09-b(DI=86.20)在感病26 d后從下到上大部分葉片上面有厚重白粉層且失綠,感病嚴重的葉片白粉層覆蓋整個葉面,而高抗親本09-a(DI=14.07)僅少部分葉片有零星的斑點及薄薄的白粉層。白粉菌脅迫下P1的感病葉片與健康功能葉相比葉綠素a、葉綠素b、總葉綠素降解量分別為0.352、0.129、0.481 mg/g,P2為0.919、0.285、1.204 mg/g,接種白粉病菌后感病自交系葉綠素a、葉綠素b、總葉綠素降解量極顯著高于抗病自交系,且葉綠素a的含量變化大于葉綠素b。病情指數與葉綠素降解量相關性分析表明(表3,表4),F2與F2:3群體接種白粉菌后,其葉綠素a、葉綠素b及總葉綠素的降解量均與病情指數呈極顯著正相關,F2群體病情指數與葉綠素a、葉綠素b及總葉綠素的降解量相關系數分別為0.572、0.515、0.565,F2:3群體相關系數分別為0.540、0.489、0.535。綜上表明苦瓜對白粉病的抗性可能與葉綠素降解量幅度的大小有關。

圖1 09-a(P1)和09-b(P2)感病后植株表型
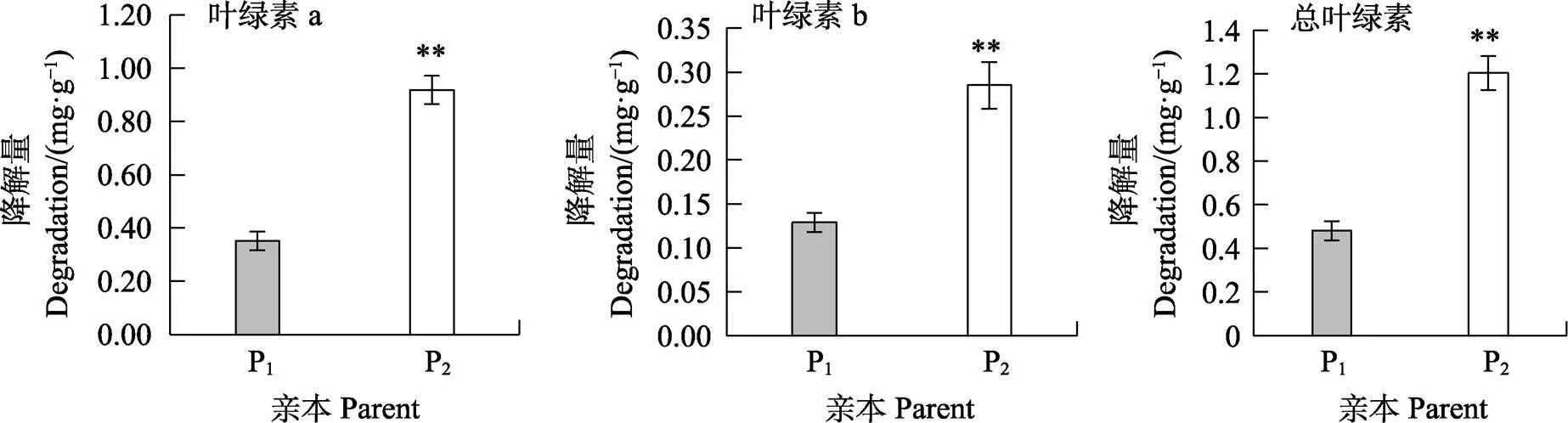
*表示差異顯著(P<0.05),**表示差異極顯著(P<0.01)。
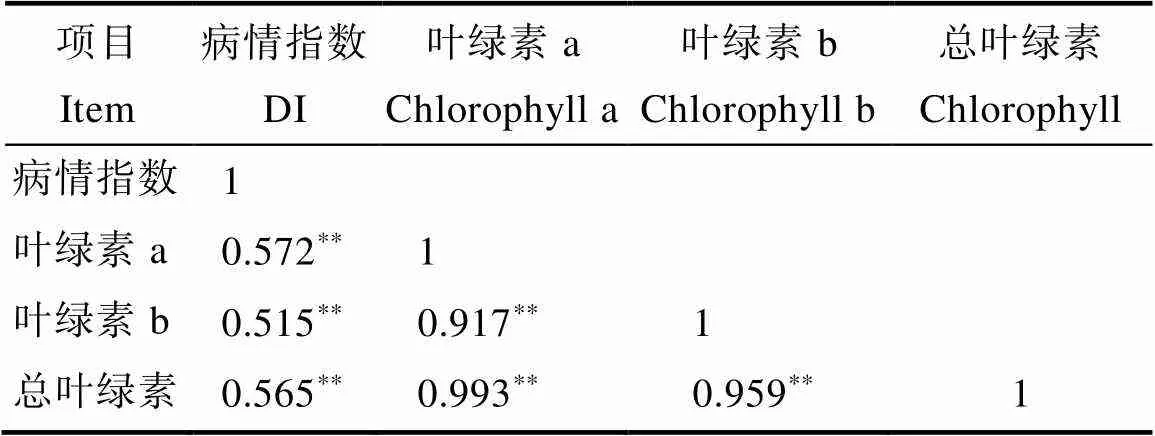
表3 F2病情指數與葉綠素降解量相關性分析
注:*表示顯著相關(<0.05),**表示極顯著相關(<0.01)。
Note:*indicates significant correlation (<0.05),**indicates extremely significant correlation (<0.01).
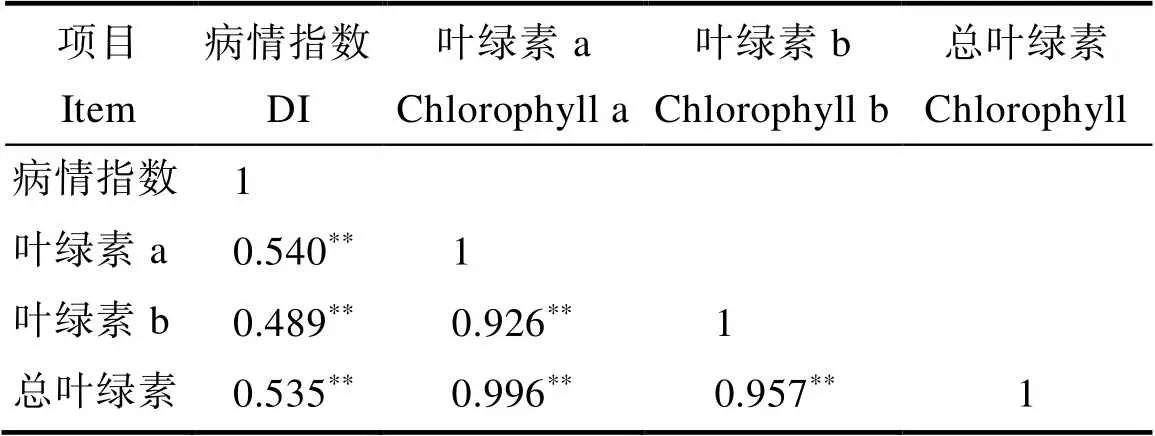
表4 F2:3病情指數與葉綠素解量相關性分析
注:*表示顯著相關(<0.05),**表示極顯著相關(<0.01)。
Note:*indicates significant correlation (<0.05),**indicates extremely significant correlation (<0.01).
白粉病菌脅迫下F2和F2:3世代的葉綠素a、葉綠素b和總葉綠素降解量均表現出連續的正態分布,偏度和峰度均小于1,且雙向超親分離,屬于數量性狀(圖3,表5),適合進一步遺傳分析。3個性狀在不同群體中呈單峰或多峰分布,表明苦瓜響應白粉病葉綠素a、葉綠素b和總葉綠素的遺傳符合主基因-多基因遺傳特征。F2群體3個性狀的變幅均較F2:3大,且3個性狀分離群體中的變異系數介于33.1%~56.1%之間,可見兩親本組合產生的F2和F2:3群體有較好的遺傳多樣性。
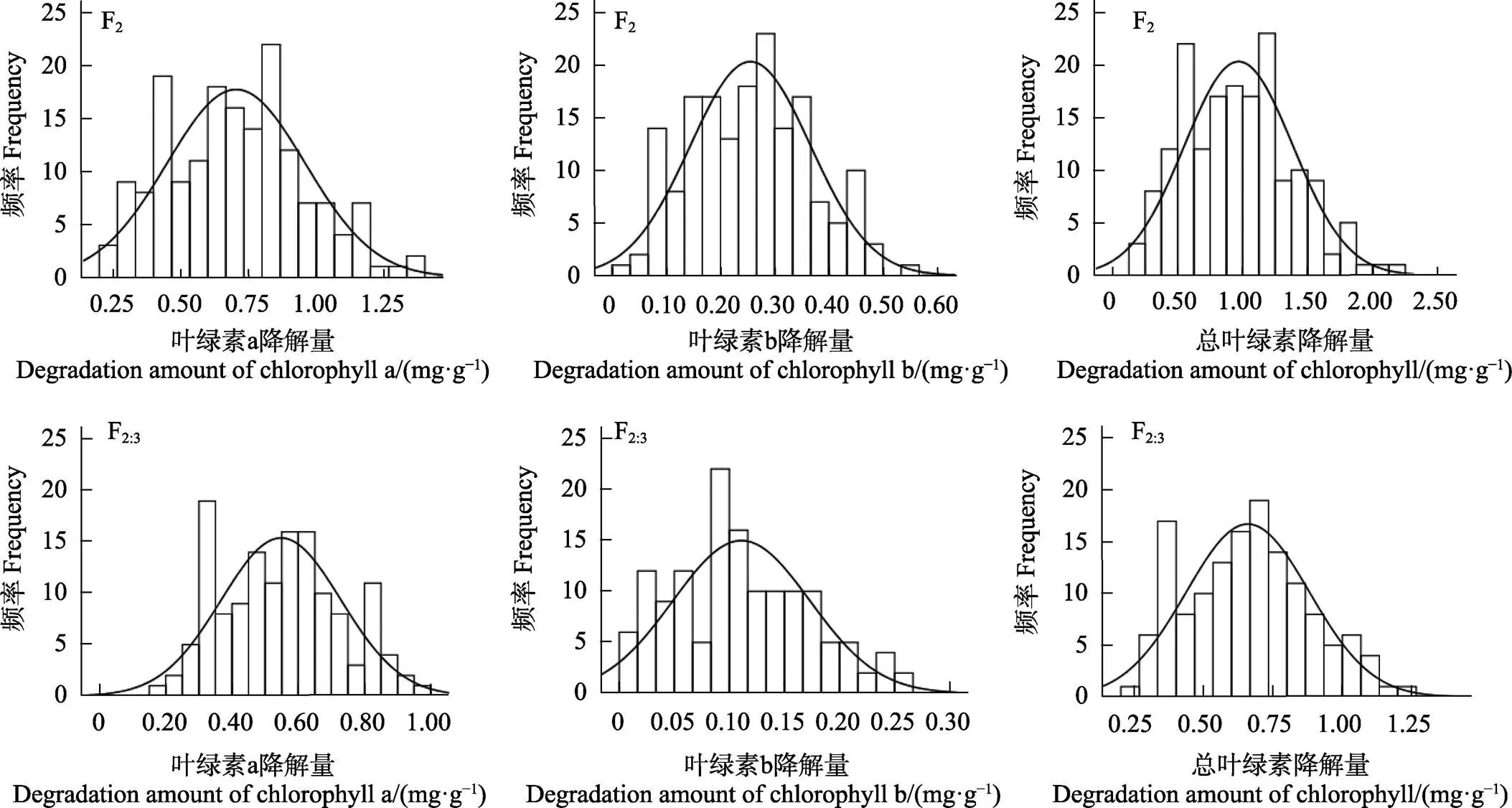
圖3 F2和F2:3群體葉綠素降解量頻率分布圖
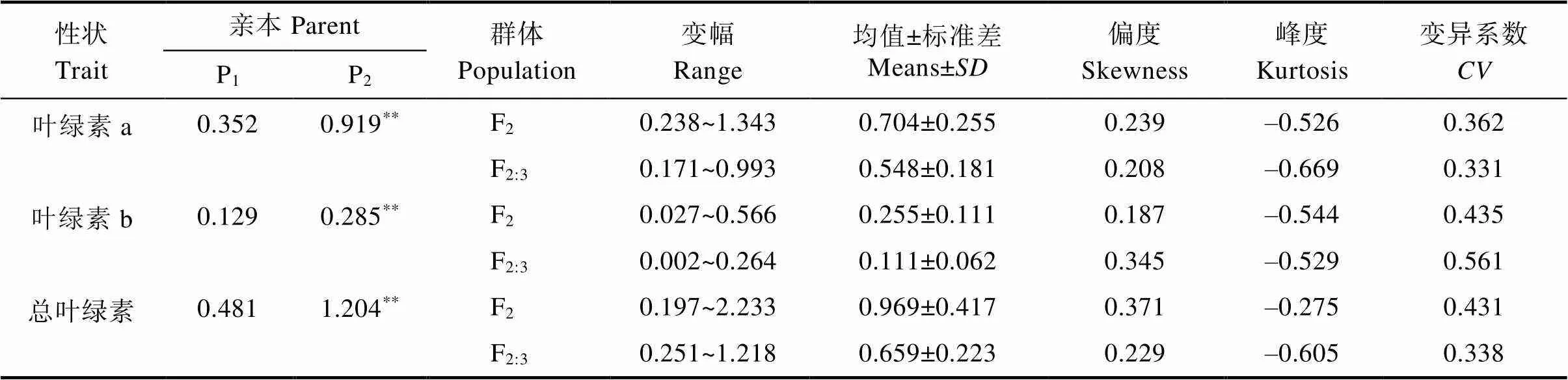
表5 白粉病菌脅迫下F2和F2:3群體相關性狀描述
注:*表示顯著相關(<0.05),**表示極顯著相關(<0.01)。
Note:*indicates significant correlation (<0.05),**indicates extremely significant correlation (<0.01).
2.2 遺傳模型的選擇及適合性檢驗
利用植物數量性狀的主基因+多基因的混合遺傳模型對F2和F2:3分別進行基于單個分離世代群體的遺傳分析,計算出各個性狀所有模型的極大似然值(MLV)和AIC值(表6,表7)。依據AIC最小原則,選擇AIC值最小為最佳可能模型,及2個與之最為接近的模型為備選模型(表8)。結果表明,F2和F2:3群體葉綠素a和葉綠素b響應白粉病菌脅迫的最小AIC值模型均為A-2,葉綠素a和葉綠素b在F2群體中的備選模型分別為B-3、A-1,在F2:3中的備選模型分別為A-1、A-0。F2和F2:3群體葉綠素降解量的最小AIC模型均為B-2,備選模型分別為F2:A-1、A-0,F2:3:A-2、A-0。經適合性檢驗(表8),發現各個模型在各項檢驗下均未能達差異顯著水平,無顯著區別,所以選擇3個備選模型中AIC最小的為最優遺傳模型。因此2個群體葉綠素a、葉綠素b最佳遺傳模型均為A-2,總葉綠素降解量的最佳模型均為B-2。說明在2個群體中白粉病菌的脅迫下葉綠素a和葉綠素b降解量均受一對加性主基因控制,葉綠素降解量則受2對加性-顯性主基因控制。
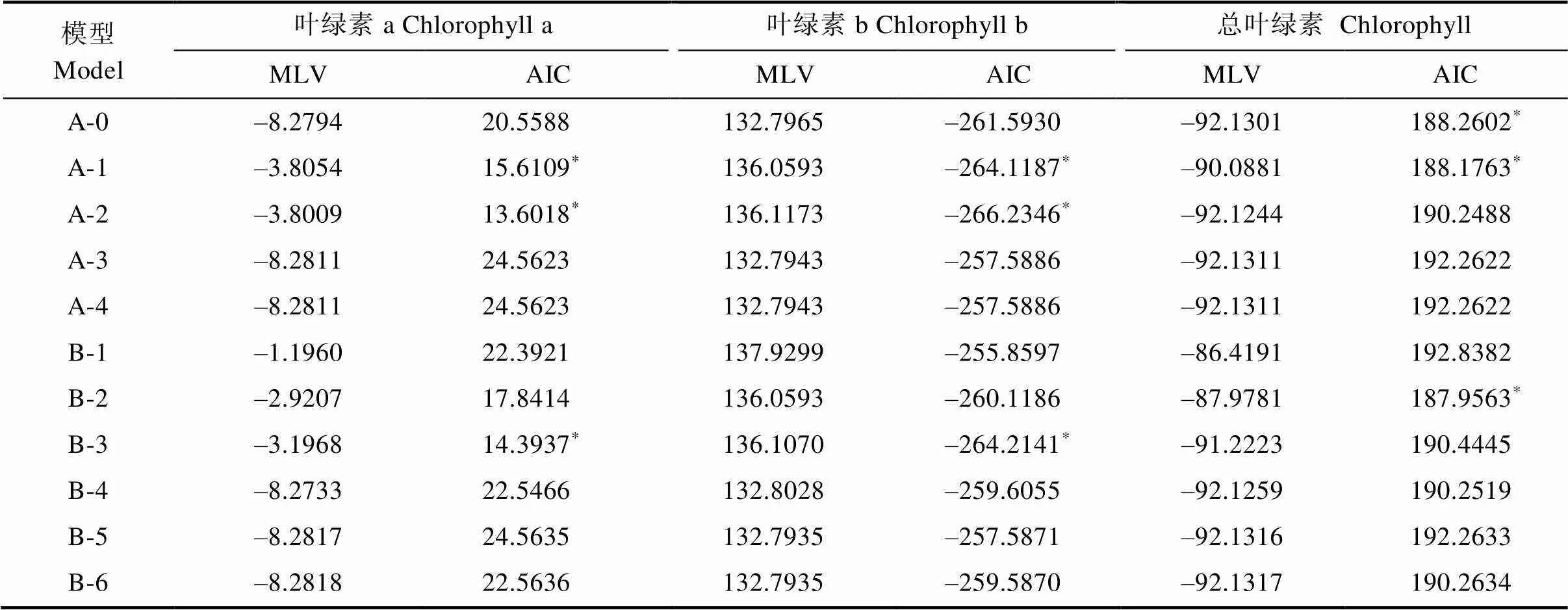
表6 F2各性狀遺傳模型的MLV和AIC值
注:*表示AIC值較小,指示的模型為較優遺傳模型。
Note:*indicates the AIC values were smaller, and indicateing models were better genetic models.
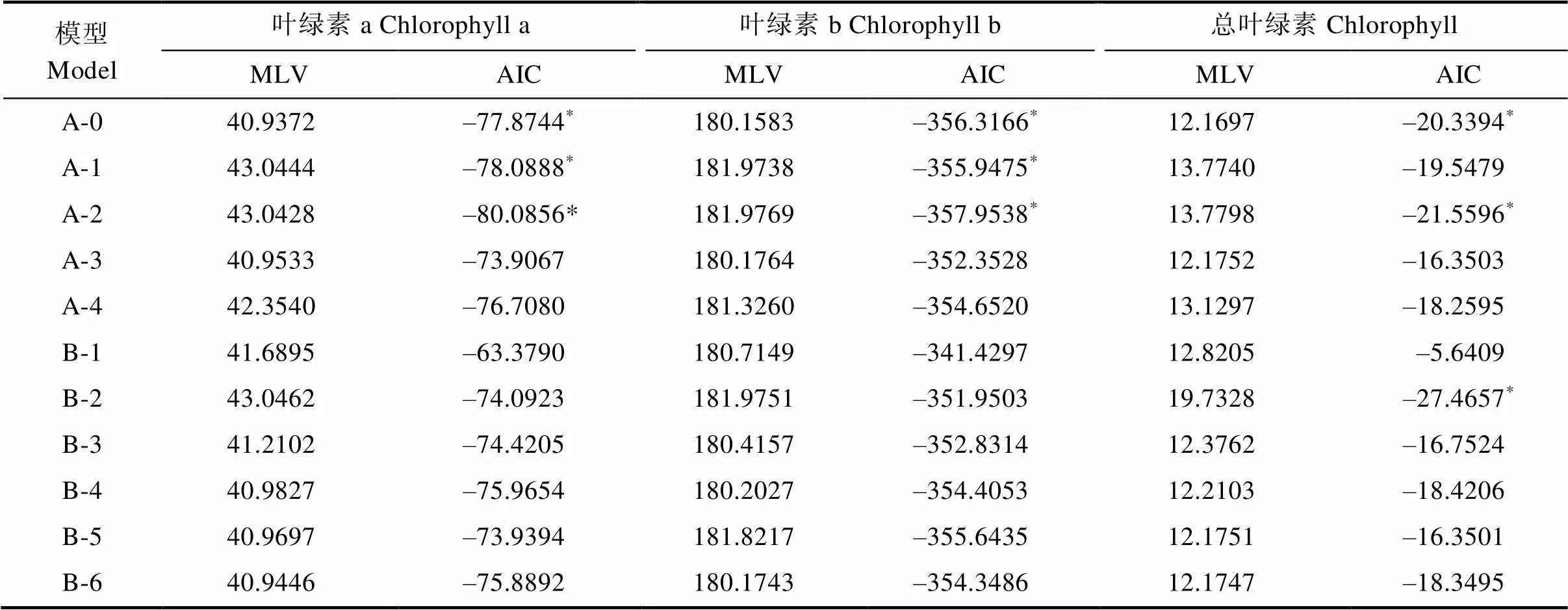
表7 F2:3各性狀遺傳模型的MLV和AIC值
注:*表示AIC值較小,指示的模型為較優遺傳模型。
Note:*indicates the AIC values were smaller, and indicateing models were better genetic models.
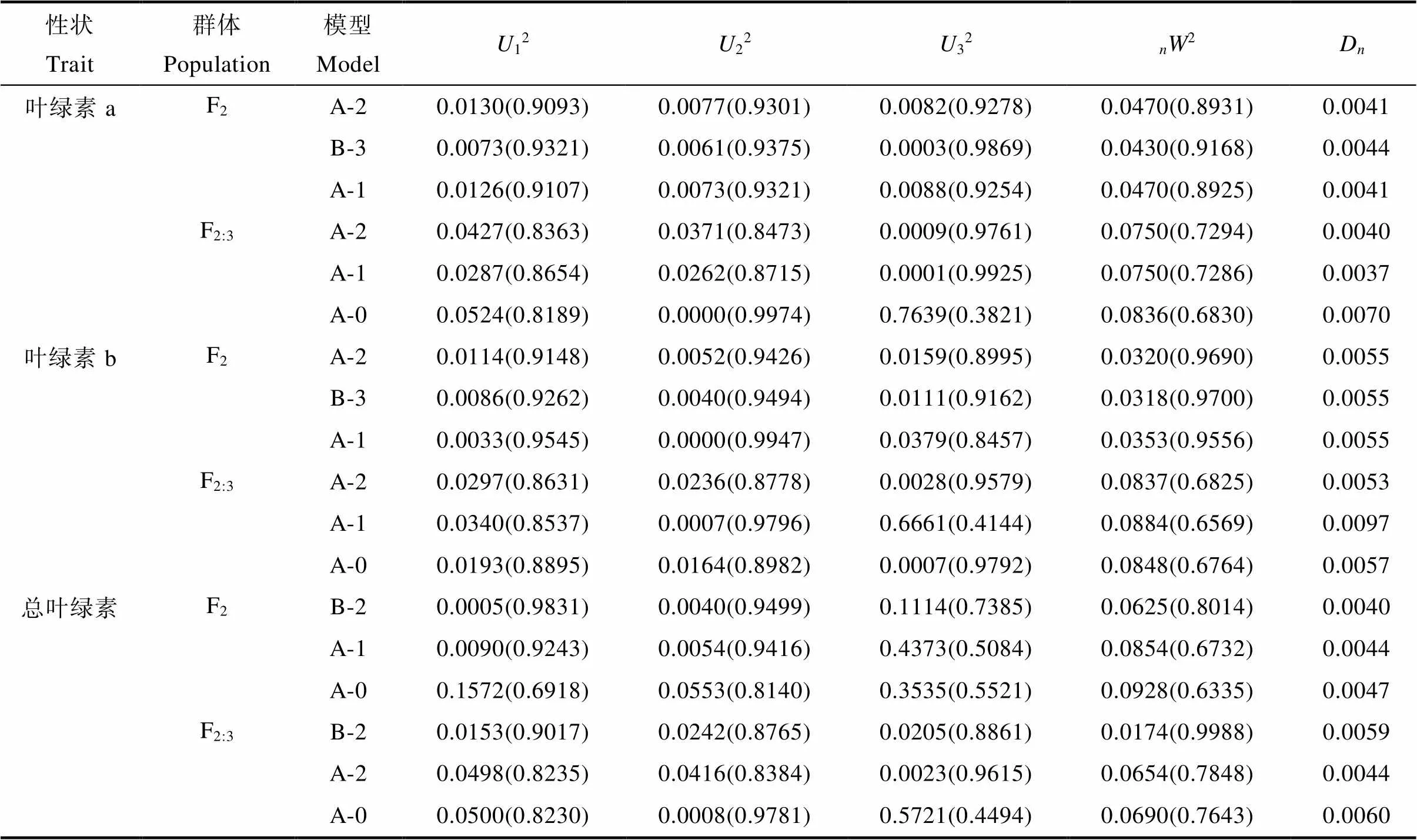
表8 各性狀備選模型的適合性檢驗
注:12、22、32、2、為適合性檢驗的統計量,括號內數據為理論概率。
Note:12,22,32,W2,are the statistics of compatibility test, the data in brackets are theoretical probabilities.
2.3 遺傳參數估計
由表9的一階參數可知,在白粉病菌脅迫下,F2和F2:3群體控制葉綠素a降解量的一對主基因加性效應分別為0.3151、0.1670,控制葉綠素b降解量的一對主基因加性效應分別為0.1333、0.1670,其加性效應均為正,白粉病脅迫下葉綠素降解量受到多個微效基因的累加。
控制感病后總葉綠素降解量的2對主基因在F2中加性效應分別為0.2601和0.3184,顯性效應為?0.3452、?0.2221,F2:3群體中2對主基因的加性效應分別為0.1892和0.1624,顯性效應分別為?0.0465、?0.0897。顯性效應為負值,表明等位基因中其隱性基因決定了葉綠素的降解量。在2個群體中其2對主基因的加性效應|a+b|大于其顯性效應值|a+b|,說明控制總葉綠素降解量以基因累加為主。然而2個群體中第一對主基因的勢能比值a/a為?1.33和?0.25,主基因的效應在2個不同地點的2個群體中表現不一,可能是由于控制數量性狀遺傳的基因其效應大小會隨著種植地點不同而有所差異。2個群體中第二對主基因a/a分別為?0.69和?0.55,主基因的顯性效用小于加性效應,具有反向的部分顯性作用。可見在苦瓜09-a和09-b組合對總葉綠素響應白粉病菌脅迫的遺傳中,2對主基因的加性和顯性效應共同發揮著重要作用。
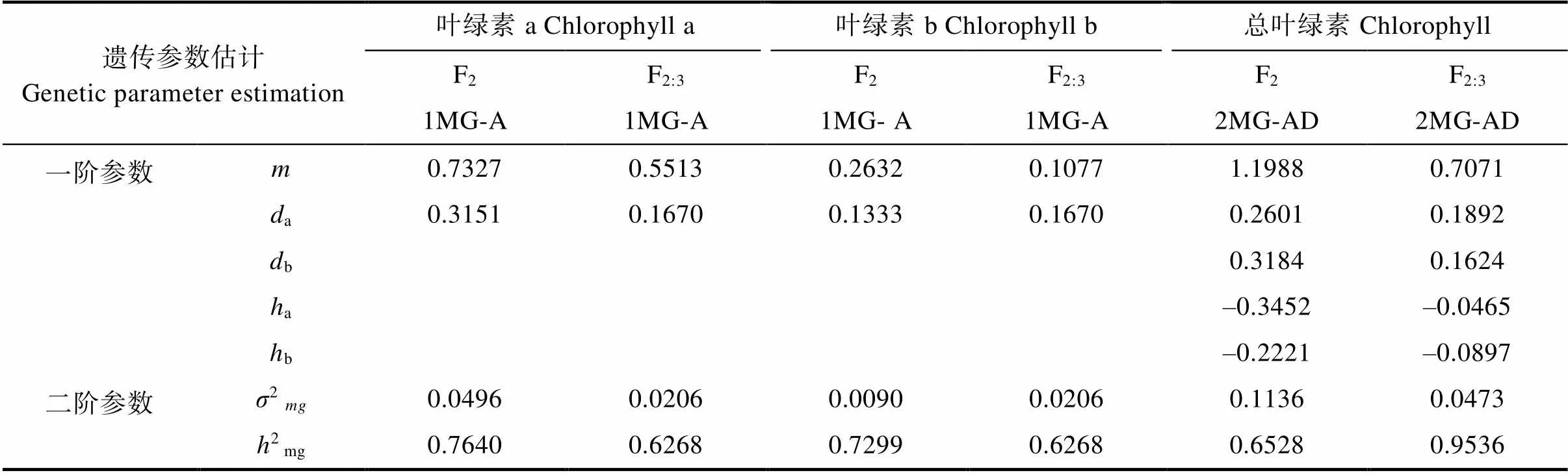
表9 各性狀最佳模型遺傳參數估計值
注:m為均值;a為第一對主基因的加性效應,b為第二對主基因的加性效應;a為第一對主基因的顯性效應,b為第二對主基因的顯性效應;2為主基因遺傳率,2為主基因方差。
Note:is means;ais additive effect of the first pair of major genes,bis additive effect of the second pair of major genes;ais dominant effect of the first pair of major genes,bis dominant effect of the second pair of major genes;his major gene inheritance rate,σ is major gene variance.
由表9的二階參數可知,在白粉病菌脅迫下F2和F2:3群體中葉綠素a降解量主基因遺傳率為76.4%、62.68%。葉綠素b降解量的主基因遺傳率分別為72.99%和62.68%,控制總葉綠素降解量2對主基因遺傳率為65.25%和95.36%。3個性狀無論是F2或者是F2:3群體遺傳率均大于60%,說明主要由主基因控制了苦瓜葉綠素對白粉病菌的響應,此外環境條件對白粉病脅迫下3個性狀的遺傳存在部分影響,可在早代對這3個性狀進行適度定向選擇。
3 討論與結論
葉綠素是地球上含量最為豐富和重要的色素,是光合作用的重要元件,通過捕獲和轉化光能成為有機質,是作物產量的主要來源[32]。葉綠素的降低是綠色植物對外界不利環境最敏感的反應,其含量的降低往往暗示著葉片的衰老、缺乏營養或遭遇脅迫,并與植株的抗性有緊密的聯系。本研究發現高抗和高感苦瓜品系被白粉病菌侵染后,感病品系葉綠素降低量極顯著的高于抗病品系。這與在黃瓜[33]、南瓜[34-35]、苦瓜[36]等瓜類上的發現類似,即白粉病抗性與葉綠素含量成正相關,同時也在葡萄[37]、小麥[38]、豌豆[39]等農作物上有相同發現。然而王惠哲等[40]、贠琰[41]分別發現南瓜、黃瓜在感染白粉病后無論抗病品種還是感病品種的葉綠素含量均有增加。但2種觀點均認為植株抗白粉病的能力與葉綠素含量呈正相關。可見葉綠素含量在抗病中起著舉足輕重的作用。因此掌握葉綠素響應白粉菌侵染的遺傳規律,能夠為抗病品種的選育提供一定的理論依據。
本研究采用植物數量的主基因+多基因的混合遺傳模型對白粉菌脅迫下F2和F2:3群體3個葉綠素相關性狀分別進行基于單個分離世代群體的遺傳分析。結果表明,2個群體的結果均表明白粉菌脅迫下葉綠素a和葉綠素b的降解量符合一對加性主基因遺傳模型,其加性效應均為正。總葉綠素降解量受2對加性顯性主基因控制且在2個群體中其2對主基因的加性效應|d+d|大于其顯性效應值|a+b|,控制總葉綠素降解量的2對主基因均以加性效應為主。而據相關研究報道,在未接種白粉病菌的情況下,以2個苦瓜葉綠素含量有差異的苦瓜品系為親本構建6個世代,利用主基因+多基因混合遺傳模型分析苦瓜葉綠素含量的遺傳規律,發現葉綠素相對含量遺傳受一對主基因控制[27, 42]。而本研究是基于白粉菌脅迫下的葉綠素降解量的遺傳分析,葉綠素含量在脅迫下所呈現的遺傳規律也有所差異。同時也表明,苦瓜在受到白粉菌侵染后,葉綠素會與病原菌相互作用。在瓊海和海口2個地點的2個群體主基因的遺傳效率在62.68%~95.36%之間,數值均較大,說明主要由主基因控制苦瓜葉綠素對白粉病菌的響應,此外環境條件對白粉病脅迫下3個性狀的遺傳存在部分影響。
綜上所述,白粉病抗性與葉綠素的降解量負相關。本研究利用2個群體分別在瓊海和海口2個地點進行遺傳分析,2個群體中,白粉病菌脅迫下葉綠素a和b的降解量均由1對加性主基因決定,總葉綠素降解量均受2對加性-顯性主基因控制,在2個不同地點的群體中得到的結論相互印證。同時環境也對葉綠素響應白粉病菌侵染有部分干擾。因此,在苦瓜白粉病抗性育種實踐中,可將葉綠素相關性狀的降低量結合病情指數在早期世代進行選擇,同時應該關注其加性效應的應用,避免主基因的累加效應所導致植株抗性的降低。而控制葉綠素a和b的2對主基因,是否與控制總葉綠素的2對主基因是否存在某種聯系尚未明確,所以在以后的研究中,將結合苦瓜高密度遺傳圖譜,對葉綠素響應白粉病菌的QTL精細定位,挖掘關鍵基因,為白粉病抗性的鑒定及抗病品種的選育奠定基礎并加速其育種進程。
[1] CUI J J, YANG Y, LUO S B, WANG L, HUANG R K, WEN Q F, HAN X X, MIAO N S, CHENG J W, LIU Z J, ZHANG C Y, FENG C C, ZHU H S, SU J W, WAN X J, HU F, NIU Y, ZHENG X M, YANG Y L, SHAN D, DONG Z S, HE W M, DHILLON N P S, HU K L. Whole-genome sequencing provides insights into the genetic diversity and domestication of bitter gourd (spp.)[J]. Horticulture Research, 2020, 7(1): 85.
[2] DHILLON N P S, LIN C C, SUN Z Y, HANSON P M, LEDESMA D R, HABICHT S D, YANG R Y. Varietal and harvesting stage variation in the content of carotenoids, ascorbic acid and tocopherols in the fruit of bitter gourd (L.)[J]. Plant Genetic Resources, 2016, 15(3): 1-12.
[3] DANDAWATE P R, SUBRAMANIAM D, PADHYE S B, ANANT S. Bitter melon:a panacea for inflammation and cancer[J]. Chinese Journal of Natural Medicines, 2016, 14(2): 81-100.
[4] ZITTER T A, HOPKINS D L, THOMAS C E. Compendium of cucurbit diseases[M]. USA: The American Phytopatholo-gi-cal Society, 1996.
[5] LEBEDA A, K?íSTKOVá E, SEDLáKOVá B, COFFEY M D, MCCREIGHT J D. Gaps and perspectives of pathotype and race determination inand[J]. Mycoscience, 2011, 52(3): 159-64.
[6] SITTERLY W R, SPENCER D M. Powdery mildews of cucurbits[M]//The Powdery Mildews. USA: The American Phytopathological Society, 1987: 359-379.
[7] HE X M, LI Y H, PANDEY S, YANDELL B S, PATHAK M, WENG Y Q. QTL mapping of powdery mildew resistance in WI 2757 cucumber (L.)[J]. Theoretical & Applied Genetics, 2013, 126(8): 2149-2161.
[8] 甘 露, 馬含月, 高京草, 張 顯. 瓜類蔬菜白粉病抗性誘導及抗性遺傳研究進展[J]. 中國瓜菜, 2021, 34(3): 1-6.
GAN L, MA H Y, GAO J C, ZHANG X. Progress on including and inheritance of resistance to powdery mildew in cucurbits[J]. Chinese Cucurbits and Vegetables, 2021, 34(3): 1-6. (in Chinese).
[9] 劉苗苗, 劉宏宇, 顧興芳, 張圣平, 苗 晗. 黃瓜白粉病抗性遺傳規律及分子標記研究進展[J]. 中國蔬菜, 2009(24): 7-12.
LIU M M, LIU H Y, GU X F, ZHANG S P, MIAO H. Research progress on inheritance and molecular markers of resistance to powdery mildew in cucumber[J]. Chinese Vegetables, 2009(24): 7-12. (in Chinese)
[10] 羅晶晶, 齊曉花, 陳學好. 瓜類作物白粉病抗性遺傳機制[J]. 分子植物育種, 2010, 8(3): 556-562.
LUO J J, QI X H, CHEN X H. Advances on genetic mechanisms of powdery mildew resistance in cucurbit crops[J]. Molecular Plant Breeding, 2010, 8(3): 556-562. (in Chinese)
[11] ALEJANDRO P G, DIEGO R, DOLORES F O, FRANCISCO L R, ANTONIO D V, JUAN A T. The powdery mildew fungus(synonym), a constant threat to cucurbits[J]. Molecular Plant Pathology, 2009, 10(2): 153-160.
[12] 劉 麗, 劉長遠, 王 輝, 于舒怡, 關天舒, 王東旭, 李柏宏. 葡萄不同抗霜霉病品種的生理生化指標研究[J]. 沈陽農業大學學報, 2018, 49(5): 594-589.
LIU L, LIU C Y, WANG H, YU S Y, GUAN T S, WANG D X, LI B H. Indices of physiology and biochemistry of various grape cultivars resistant to downy mildew[J]. Journal of Shenyang Agricultural University, 2018, 49(5): 594-599. (in Chinese)
[13] 歐志遠. 葉綠素含量與植物抗病性的關系[J]. 安徽農學通報, 2007(6): 134-135.
OU Z Y. Relationship between chlorophyll content and plant disease resistance[J]. Anhui Agricultural Science Bulletin, 2007(6): 134-135. (in Chinese)
[14] MORADI N, RAHIMIAN H, DEHESTANI A, BABA-EI-ZAD V, YAGHOUBIAN Y. Screening of cucumber cultiv-ars resistant to powdery mildew and comparative assessment of chlorophyll florescence in resistant and sensitive cultivars[J]. Journal of Plant Protection, 2017, 31(3): 83-85.
[15] AZMAT M A, KHAN A A, NIAZ S. Stomatal density and chlorophyll concentration as an indicator of powdery mildew resistance in pea (L.)[J]. Pakistan Journal of Agricultural Sciences, 2016, 53(4): 871-877.
[16] 王好讓, 張 勇, 于春淼, 董全中, 李微微, 胡凱鳳, 張明明, 薛 紅, 楊夢平, 宋繼玲, 王 磊, 楊興勇, 邱麗娟. 大豆突變體黃綠葉基因的精細定位[J]. 作物學報, 2022, 48(4): 791-800.
WANG H R, ZHANG Y, YU C M, DONG Q Z, LI W W, HU K F, ZHANG M M, XUE H, YANG M P, SONG J L, WANG L, YANG X Y, QIU L J. Fine mapping of yellow-green leaf gene () in soybean (L.)[J]. Acta Agronomica Sinica, 2022, 48(4): 791-800. (in Chinese)
[17] 康偉偉, 李哲理, 易自力, 孫志忠, 盛夏冰, 黃安平, 段美娟, 譚炎寧. 水稻黃葉突變體()的鑒定與遺傳分析[J]. 農業生物技術學報, 2020, 28(12): 2108-2117.
KANG W W, LI Z L, YI Z L, SUN Z Z, SHENG X B, HUANG A P, DUAN M J, TAN Y N. Identification and genetic analysis of the yellow leaf mutant()in rice ()[J]. Journal of Agricultural Biotechnology, 2020, 28(12): 2108-2117. (in Chinese)
[18] 袁 鎧. 水稻黃葉突變體()的遺傳分析和基因定位探究[J]. 南方農業, 2020, 14(17): 182-183.
YUAN K. Genetic analysis and gene mapping of the yellow leaf mutant() in rice[J]. South China Agriculture, 2020, 14(17): 182-183. (in Chinese)
[19] 戴 楠, 袁光孝, 劉小敏, 高宏波. 擬南芥黃化突變體的基因作圖定位[J]. 北京林業大學學報, 2017, 39(7): 40-45.
DAI N, YUAN G X, LIU X M, GAO H B. Gene mapping ofyellowing mutant[J]. Journal of Beijing Foresty University, 2017, 39(7): 40-45. (in Chinese)
[20] 葉景秀, 柳海東. 葉綠素含量相關QTL定位研究進展[J]. 青海大學學報, 2019, 37(4): 55-60.
YE J X, LIU H D. Research progress of QTL mapping of chlorophyll content[J]. Journal of Qinghai University, 2019, 37(4): 55-60. (in Chinese)
[21] 蓋鈞鎰. 植物數量性狀遺傳體系的分離分析方法研究[J]. 遺傳, 2005, 27(1): 130-136.
GAI J Y. Segregation analysis of genetic system of quantitative traits in plants[J]. Hereditas, 2005, 27(1): 130-136. (in Chinese)
[22] 田麗波. 苦瓜遺傳圖譜構建及白粉病抗性的QTL定位[D]. 沈陽: 沈陽農業大學, 2015.
TIAN L B. Constructing of genetic maps and QTL mapping of powdery mildew resisitance in bitter melon[D]. Shenyang: Shenyang Agricultural University, 2015. (in Chinese)
[23] 梁慧珍, 余永亮, 楊紅旗, 董 薇, 許蘭杰, 牛永光, 張海洋, 劉學義, 方宣鈞. 大豆葉片性狀和葉綠素含量QTL間的上位性和環境互作效應[J]. 作物學報, 2015, 41(6): 889-899.
LIANG H Z, YU Y L, YANG H Q, DONG W, XU L J, NIU Y G, ZHANG H Y, LIU X Y, FANG X J. Epistatic and QTL × environment interaction effects of QTLs for leaf traits and leaf chlorophyll content in soybean[J]. Acta Agronomica Sinica, 2015, 41(6): 889-899. (in Chinese)
[24] 李洪濤, 許瀚元, 李景芳, 祝 慶, 遲 銘, 王 軍. 玉米葉綠素含量基因效應分析[J]. 作物雜志, 2019, 192(5): 46-51.
LI H T, XU H Y, LI J F, ZHU Q, CHI M, WANG J. Gene effect analysis of chlorophyll content in maize[J]. Corps, 2019, 192(5): 46-51. (in Chinese)
[25] 林婷婷, 王 立, 張 琳, 王建軍, 侯喜林, 李 英. 不結球白菜葉綠素含量的主基因+多基因混合遺傳分析[J]. 南京農業大學學報, 2014, 37(5): 34-40.
LIN T T, WANG L, ZHANG L, WANG J J, HOU X L, LI Y. Genetic analysis of chlorophyll content using mixed major gene plus polygene inheritance model in non-heading Chinese cabbage[J]. Journal of Nanjing Agricultural University, 2014, 37(5): 34-40. (in Chinese)
[26] 張興偉, 王志德, 牟建民, 劉艷華, 任 民. 烤煙葉綠素含量遺傳分析[J]. 中國煙草學報, 2011, 17(3): 48-52.
ZHANG X W, WANG Z D, MOU J M, LIU Y H, REN M. Genetic analysis of chlorophyll content in flue-cured tobacco leaf[J]. Acta Tabacaria Sinica, 2011, 17(3): 48-52. (in Chinese)
[27] 陳虹容, 商 桑, 田麗波, 朱國鵬. 苦瓜葉片葉綠素含量的遺傳分析[J]. 熱帶作物學報, 2021, 42(6): 1572-1578.
CHEN H R, SHANG S, TIAN L B, ZHU G P. Genetic anal-y-sis of chlorophyll content in bitter gourd leaves[J]. Chinese Journal of Tropical Crops, 2021, 42(6): 1572-1578. (in Chinese)
[28] 田麗波, 張 燕, 商 桑, 楊 衍. 基于生理生化性狀和病情指數評價苦瓜種質資源的白粉病抗性[J]. 分子植物育種, 2015, 13(12): 2824-32.
TIAN L B, ZHANG Y, SHANG S, YANG Y. Assessment of powdery mildew resistance of bitter melon germplasm resources based on physiological and biochemical indexes and disease index[J]. Molecular Plant Breeding, 2015, 13(12): 2824-2832. (in Chinese)
[29] SAKATA Y, KUBO N, MORISHITA M, KITADANI E, SUGIYAMA M, HIRAI M. QTL analysis of powdery mildew resistance in cucumber (L.)[J]. Theoretical and Applied Genetics, 2006, 112(2): 243-250.
[30] 粟建文, 胡新軍, 袁祖華, 李勇奇, 劉雪源. 苦瓜白粉病抗性遺傳規律研究[J]. 中國蔬菜, 2007(9): 24-26.
SU J W, HU X J, YUAN Z H, LI Y Q, LIU X Y. The inheritance of resistance to powdery mildew in bitter ground[J]. China Vegetables, 2007(9): 24-26. (in Chinese)
[31] 李合生. 植物生理生化實驗原理和技術[M]. 北京: 高等教育出版社, 2000: 246- 247.
LI H S. Experimental principles and techniques of plant physiology and biochemistry[M]. Beijing: Hight Education Press, 2000. (in Chinese)
[32] FROMME P, MELKOZERNOV A, JORDAN P, KRAUSS N. Structure and function of photosystem I: interaction with its soluble electron carriers and external antenna systems[J]. FEBS Letters, 2003, 555(1): 40-44.
[33] 金海軍, 胡繼軍, 卜立君, 丁小濤, 余紀柱, 張紅梅. 華南型黃瓜自交系白粉病抗性與光合特性的關系[J]. 中國瓜菜, 2020, 177(2): 32-35.
JIN H J, HU J J, BU L J, DING X T, YU J Z, ZHANG H M. Relationship between powdery mildew resistance and photosynthetic characteristics of cucumber inbred lines of South China type[J]. Chinese Cucurbits and Vegetables, 2020, 177(2): 32-35. (in Chinese)
[34] 顏惠霞, 徐秉良, 梁巧蘭, 薛應鈺, 陳榮賢, 梁志福. 南瓜品種對白粉病的抗病性與葉綠素含量和氣孔密度的相關性[J]. 植物保護, 2009, 35(1): 79-81.
YAN H X, XU B L, LIANG Q L, XUE Y Y, CHEN R X, LIANG Z F. Effects of chlorophyll content and stoma density on pumpkin resistance to powdery mildew[J]. Plant Protection, 2009, 35(1): 79-81. (in Chinese)
[35] 郭衛麗, 郭言言, 李新崢. 中國南瓜苗期性狀與白粉病抗性的關系[J]. 北方園藝, 2016(24): 104-108.
GUO W L, GUO Y Y, LI X Z. Relationship between morphological index and resistance to powdery mildew in pum-pkin[J]. Northern Horticulture, 2016(24): 104-108. (in Chinese)
[36] 田麗波, 商 桑, 楊 衍, 司龍亭. 苦瓜對白粉病的抗性與相關生理生化指標的關系[J]. 西北農業學報, 2015, 24(9): 166-173.
TIAN L B, SHANG S, YANG Y, SI L T. Relationship between bitter melon’s resistance to powdery mildew with associated physiology and biochemical indexes[J]. Acta Agriculturae Boreali-occidentalissinica, 2015, 24(9): 166-173. (in Chinese)
[37] 吳 昊, 董華芳, 許延波. 番茄白粉病對番茄葉片葉綠素含量的影響[J]. 北方園藝, 2011(16): 8-10.
WU H, DONG H F, XU Y B. Effect of tomato powdery mildew on chlorophyll content in tomato leaves[J]. Northern Horticulture, 2011(16): 8-10. (in Chinese)
[38] 韓慶典, 楊美娟, 黃擇祥, 閆 麗, 胡曉君. 小麥白粉病菌對小麥幼苗光合生理特性的影響[J]. 基因組學與應用生物學, 2017, 36(10): 4373-4379.
HAN Q D, YANG M J, HUANG Z X, YAN L, HU X J. Effects of wheat powdery mildew on the photosynthetic physi-o-logical characteristics of wheat seedlings[J]. Genomics and Applied Biology, 2017, 36(10): 4373-4379. (in Chinese)
[39] 張麗娟, 楊曉明, 陸建英, 王 昶. 豌豆白粉病抗性相關指標的研究[J]. 甘肅農業科技, 2015(3): 33-36.
ZHANG L J, YANG X M, LU J Y, WANG C. Study on indicators related with pea powdery mildew resistance[J]. Gansu Agricultural Science and Technology, 2015(3): 33-36. (in Chinese)
[40] 王惠哲, 李淑菊, 霍振榮, 龐金安. 黃瓜感染白粉病菌后的生理變化[J]. 2016, 21(1):105-109.
WANG H Z, LI S J, HUO Z R, PANG J A. Physiological changes of cucumber after being infected by[J]. Acta Agriculturae Boreali-Sinica, 2016, 21(1): 105-109. (in Chinese)
[41] 贠 琰. 南瓜抗白粉病機理的初步研究[D]. 南京: 南京農業大學, 2017.
YUN Y. Preliminary study on resistance mechanism of pumpkin to powdery mildew[D]. Nanjing: Nanjing Agricultural University, 2017. (in Chinese)
[42] 張 燕, 王佳楠, 商 桑, 田麗波, 杜卓濤, 周萌萌. 苦瓜苗期幾個主要性狀的遺傳分析[J]. 分子植物育種, 2017, 15(5): 1906-1917.
ZHANG Y, WANG J N, SHANG S, TIAN L B, DU Z T, ZHOU M M. Genetic analysis on several main seedling characters of bitter melon[J]. Molecular Plant Breeding, 2017, 15(5): 1906-1917. (in Chinese)
Genetic Law of Chlorophyll Response to Powdery Mildew Infection in Bitter Melon Leaves
YANG Yi1, TIAN Libo1*, SHANG Sang2*, CHEN Hongrong1, FAN Chenwei1, GUO Xuesong1
1. College of Horticulture, Hainan University / Hainan Key Laboratory of Tropical Horticultural Crop Quality Control, Haikou, Hainan 570228, China; 2. College of Life Sciences, Hainan University, Haikou, Hainan 570228, China
In order to explore the genetic law of chlorophyll a, chlorophyll b and chlorophyll in response to powdery mildew infection in bitter melon seedlings, 170 individual plants of F2and 140 families of F2:3were constructed with bitter melon high-generation inbred lines 09-a and 09-b, The genetic analysis of two populations in two environments based on a single separated generation population was carried out using the major gene + polygene mixed genetic model of plant quantitative inheritance. The results showed that the contents of chlorophyll a, chlorophyll b and chlorophyll decreased under the stress of powdery mildew. The reduction range of susceptible strains was significantly greater than that of resistant strains. The degradation amount was positively correlated with the disease index, and the degradation amount was normally distributed in the two populations. The degradation amounts of chlorophyll a and chlorophyll b in F2and F2:3populations were determined by a pair of additive major genes. The additive effect of a pair of major genes controlling the degradation amount of chlorophyll a in the two populations was 0.3151 and 0.1670 respectively, and the additive effects of a pair of major genes controlling the degradation amount of chlorophyll b was 0.1333 and 0.1670 respectively. The chlorophyll degradation was controlled by two pairs of additive dominant major genes. The additive effect of the two pairs of major genes in F2population was 0.2601 and 0.3184, the dominant effect was ?0.3452 and -0.2221, and the additive effect in F2:3population was 0.1892 and 0.1624, the dominant effect was -0.0465 and ?0.0897, respectively. The additive and dominant effects of the two pairs of major genes played an important role together. The additive effects of the three traits were positive, and the two pairs of major genes controlling the chlorophyll degradation were dominated by additive effects. The major gene heritability of chlorophyll a degradation in F2and F2:3populations was 76.40% and 62.68%. The heritabilities of major genes controlling chlorophyll b degradation was 72.99% and 62.86%, respectively. And the heritability of two pairs of major genes controlling chlorophyll b degradation was 65.25% and 95.36%, both were greater than 60%, indicating that chlorophyll a, chlorophyll b and chlorophyll degradation were mainly controlled by main genes under powdery mildew stress, but it was also affected by some environment. In the breeding practice of bitter melon powdery mildew resistance, the reduction of chlorophyll related traits could be selected in the early generation combined with the disease index. This study would provide a theoretical basis for exploring the disease resistance mechanism and disease resistance breeding of bitter melon powdery mildew.
bitter melon; chlorophyll; genetic analysis; major gene + polygene model
S436.429
A
10.3969/j.issn.1000-2561.2022.09.017
2022-01-26;
2022-04-27
2019年海南省基礎與應用基礎研究計劃(自然科學領域)高層次人才項目(No. 2019RC150);國家自然科學基金項目(No. 32060682);海南省研究生創新科研課題基金項目(No. Hys2020-236)。
楊 怡(1997—),女,碩士研究生,研究方向:園藝植物遺傳育種。*通信作者(Corresponding author):田麗波(TIAN Libo),E-mail:faiy7play@163.com;商 桑(SHANG Sang),E-mail:fair.play@163.com。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
