β-catenin基因對合作豬絨毛生長發育的影響
2022-10-19 00:28:56孔夢潔耿永琪秦永斌黃曉宇滾雙寶
甘肅畜牧獸醫 2022年10期
孔夢潔,耿永琪,楊 磊,秦永斌,黃曉宇,2*,滾雙寶,2
(1.甘肅農業大學 動物科學技術學院,甘肅 蘭州 730070;2.甘肅省現代養豬工程技術研究中心,甘肅 蘭州 730070)
合作豬又被稱為蕨麻豬、山豬,具有適應高海拔惡劣環境、抗寒、抗病、耐粗飼等特點,個體較小,體軀較短,四肢結實緊湊,周身被毛黑色,少數出現棕色。寒冷季節,合作豬身上會長出棕色絨毛,溫暖炎熱季節身上絨毛退化,直至消失,這種現象可能與合作豬周期性生長絨毛以抵御寒冷環境,從而適應高海拔高寒地區的環境有關。因此,探究合作豬絨毛生長相關基因對于研究合作豬的抗寒性能具有重要意義。
β-catenin基因是機體內一種重要的功能蛋白,可與很多蛋白相互結合形成復合體[1-2]。其遍布動物體各個結構中,功能十分豐富,且在動物進化過程中非常保守。截至目前,主要在動物間質組織和上皮組織中發現了β-catenin基因的存在。研究證實,β-catenin基因在毛發生長中具有調節作用。當β-catenin基因發生突變或被敲除后,存在于動物體皮膚中的毛囊則不會繼續發育,而補償缺失的β-catenin基因后,毛囊又會長出新的毛發,這充分說明β-catenin基因在毛囊生長發育過程中是必須存在的[3-4]。還有研究表明,在毛囊的發生發育過程中β-catenin基因會被激活,以此促進細胞的增殖分化,從而促進毛囊結構的發生發育[5]。此外,Plikus等[6]以小鼠皮膚組織作為模型探究毛囊膨出處的毛干細胞調控機制時,發現β-catenin基因在炎熱與寒冷環境下分別表現為高活性和低活性,說明其在毛干細胞調控過程中與溫度變化呈正相關。目前關于β-catenin基因的研究主要是圍繞人體β-catenin基因開展,而關于豬β-catenin基因及其在絨毛生長發育過程中的研究鮮有報道。
本試驗以夏季和冬季的合作豬皮膚組織為研究材料,通過qRT-PCR探究β-catenin基因在合作豬絨毛生長過程中的表達特征及作用。
1 材料與方法
1.1 試驗材料
1.1.1 實驗動物 本研究以甘南州迭部縣8月齡合作豬為研究對象,在夏季極熱和冬季極冷的時間點采集5 cm×1 cm的豬皮膚。具體采集頸部、肩部、側部、臀部等組織樣品作為本次試驗樣品。
1.1.2 主要試劑、耗材 Trizol新型RNA抽提劑、2*Taq PCR MasterMix、反轉錄試劑盒、dNTP Mixture、Oligo dT Primer、RNase Inhibitor、DNA凝膠回收試劑盒等。
1.2 試驗方法
1.2.1 樣品采集 在夏季極熱和冬季極冷時間點分別選擇健康的8 月齡合作豬各20 頭。將兩組試驗對象宰殺后,在無菌條件下采集其頸部、肩部、側部、臀部等部位大小為5 cm×1 cm的皮膚組織樣品備用,將樣品在液氮中快速冷凍,放到-80 ℃冰箱存放。
1.2.2 引物設計與合成 以GenBank中的豬β-catenin基因核苷酸序列為參考,運用NCBI中的Primer Premier 5.0和Primer-BLAST工具,按照定量引物設計的一般原則,設計本次實時熒光定量PCR分析試驗所需的特異性引物,并由陜西中科羽瞳生物科技有限公司合成,引物信息如表1所示。
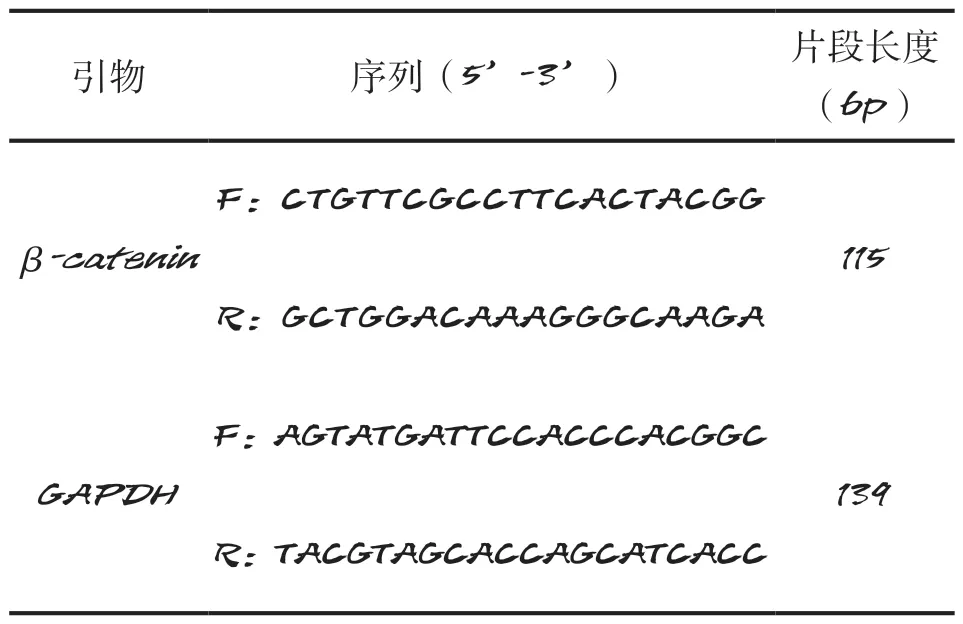
表1 引物信息
1.2.3 RNA提取與cDNA模板合成 采用Trizol試劑抽提合作豬仔豬的臀部、側部、肩部和頸部等部位組織的RNA,分別運用凝膠電泳法、分光光度計法檢測所抽提的RNA(驗證完整性和純度),將得到的cDNA放到-20 ℃冰箱保存。
1.2.4 實時熒光定量PCR 分別以不同部位組織的cDNA為模板,以GAPDH基因為內參基因,采用qRT-PCR檢測β-catenin基因在合作豬臀部、側部、頸部和肩部等組織的表達量。qRTPCR反應體系、反應條件分別見表2、表3。為減少試驗誤差,每組設3個重復,用2-ΔΔCt法計算相對表達量。

表2 qRT-PCR反應體系
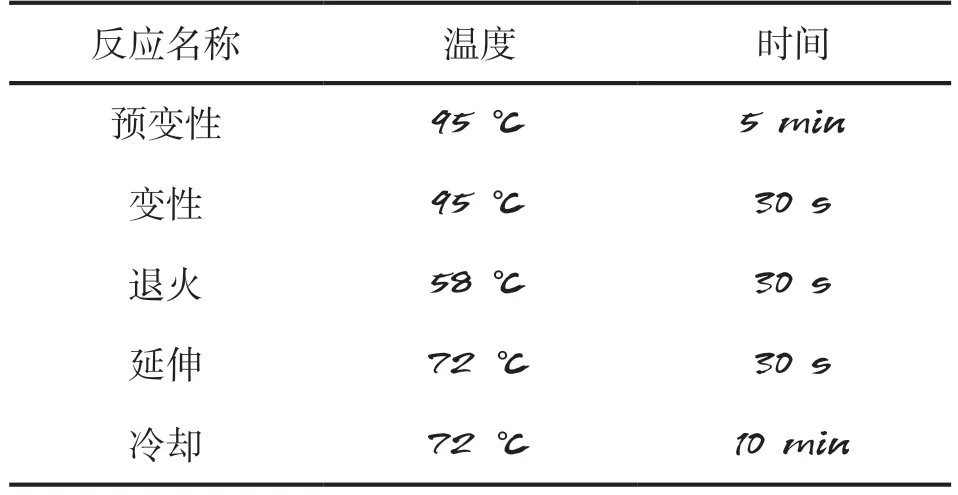
表3 qRT-PCR反應條件
2 結果與分析
2.1 β-catenin基因擴增曲線
從β-catenin基因的擴增曲線可以看出(圖1),雖然曲線在平臺期有所變化,但在指數增長區域的重復性非常好,說明適宜進行聚合酶鏈式反應(PCR)的定量分析。
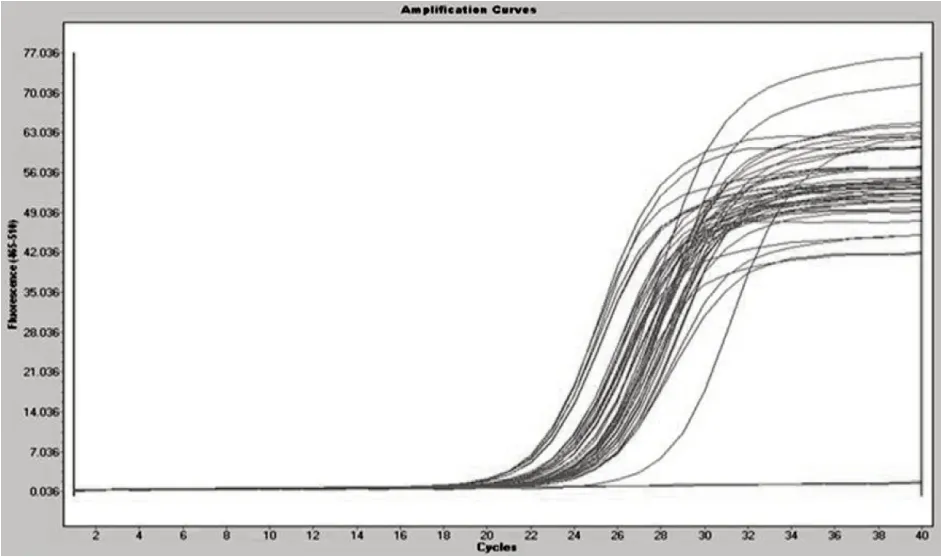
圖1 β-catenin基因的擴增曲線
2.2 β-catenin基因溶解曲線
如圖2所示,β-catenin基因的溶解曲線峰形為單一窄峰,無雜峰出現,說明該試驗所設計的引物特異性良好,可以進行后續試驗分析。
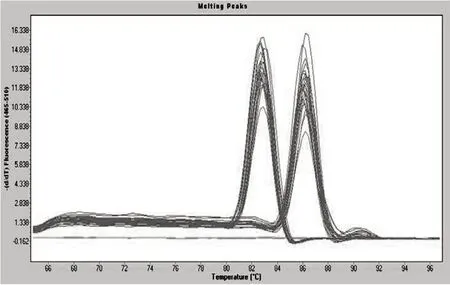
圖2 β-catenin基因的溶解曲線
2.3 β-catenin基因在不同組織中的表達
通過qRT-PCR檢測合作豬各組織中β-catenin基因的表達水平(表4、圖3),發現合作豬4個部位中β-catenin基因的表達量夏季均高于冬季,說明β-catenin基因的表達量與溫度變化呈正比。此外,冬季合作豬β-catenin基因在肩部(0.85±0.13)和臀部(0.79±0.14)的表達量相比側部(0.62±0.11)和頸部(0.50±0.077)要高,而夏季肩部(0.85±0.14)和臀部(0.89±0.0071)較側部(1.00±0.08)和頸部(0.92±0.091)的表達量低,說明合作豬肩部和臀部的β-catenin基因隨著溫度變化表達量較為恒定,而側部和頸部的β-catenin基因表達量隨溫度變化起伏較大。以上結果表明,合作豬β-catenin基因與溫度變化呈正相關,但隨著溫度的變化,肩部和臀部的β-catenin基因表達較為恒定,這或許是冬季合作豬肩部和臀部絨毛生長的原因之一,但具體分子調控機制還有待研究。

表4 熒光定量分析結果

圖3 β-catenin基因在合作豬夏、冬季節不同組織中的表達水平
3 討論
毛囊的生長發育依靠多種信號分子的共同作用,其中大部分的信號分子屬于Wnt信號通路以及一些其他傳導通路和因子等。上皮細胞和真皮細胞受到這些信號分子的共同作用,促進細胞的增殖與分化,并最終形成毛囊結構。Wnt/β-catenin信號通路與分子激活在毛囊的生長和發育中起著重要的調節作用[7]。β-catenin是經典Wnt/β-catenin信號通路中的主要調控蛋白中的一種,具有促進細胞增殖的作用[8]。β-catenin基因是毛發生成過程中的重要存在,其突變會導致毛囊基板的產生被阻斷,敲除毛囊中β-catenin基因會導致毛發掉落[9]。目前關于β-catenin基因的研究主要是圍繞人體β-catenin基因開展,而關于豬β-catenin基因及其在絨毛生長發育過程中的相關研究較少。
動物絨毛的生長是季節性變化的,大部分動物皮膚表層會在秋冬季節長出絨毛,依靠絨毛的防寒性能抵御寒冷[10]。季節性換毛是動物為了應對環境溫度的變化所作出的適應性反應[11]。合作豬絨毛的生長同樣具有季節性,秋季新生絨毛生長出體表,秋冬季快速生長,冬春寒冷季節絨毛密度達到最大,以此抵御寒冷環境氣候。本試驗利用qRT-PCR方法檢測了夏冬兩季合作豬頸部、肩部、體側和臀部皮膚組織中β-catenin基因的表達情況,發現β-catenin基因的表達量夏季高于冬季,且頸部和側部中β-catenin基因的表達量夏季顯著高于冬季(P<0.05),但肩部和臀部中β-catenin基因的表達量隨溫度變化較為恒定。以上結果與Plikus等[6]對β-catenin基因在不同季節小鼠皮膚中的變化規律結果相一致,即合作豬皮膚中β-catenin基因的mRNA表達量與溫度變化呈正相關。在整個絨毛生長過程中,β-catenin基因的mRNA表達量在夏冬兩季存在差異,尤其在冬季絨毛生長發育最迅速的時期,其表達量在頸部和側部顯著降低,但肩部和臀部的表達量與夏季相比較為恒定,表明β-catenin基因的表達可能調控絨毛的發生[12-13],同時β-catenin基因對毛囊干細胞分化起作用[6],從而影響絨毛生長,但具體調控機制還有待研究。
4 結論
通過檢測夏季和冬季β-catenin基因在合作豬豬體中的表達變化,發現合作豬β-catenin基因在頸部與側部中的表達量夏季顯著高于冬季(P<0.05)。因此推測β-catenin基因與合作豬周期性絨毛生長有關。本研究結果可為探索合作豬絨毛生長調控分子機制和毛發周期性生長調控機制提供理論依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
