基于高通量測序技術挖掘分子遺傳標記及其在蛋雞生產中的精準應用
2022-10-20 08:27:48趙金波潘洪彬王光正豆騰飛李子健葛長榮
中國畜牧獸醫 2022年10期
趙金波,譚 磊,潘洪彬,黃 暢,劉 永,王光正,豆騰飛,王 坤,李子健,葛長榮
(1.云南農業大學動物科學技術學院,昆明 650201;2.黑龍江省農業科學院畜牧獸醫分院,齊齊哈爾 161005)
中國地方畜禽品種資源豐富,畜禽遺傳多樣性的可持續利用是打好種業翻身仗,解決產業“卡脖子”核心技術的最主要育種素材,也是保障食物充足供應的先決條件。《全國畜禽遺傳改良計劃(2021—2035年)》遠景規劃著重指出“力爭用10~15年的時間,建成比較完善的商業化育種體系,顯著提升畜禽生產水平和品質水平,自主培育一批具有國際競爭力的突破性品種,確保國內畜禽品種核心種源自主可控”[1],特別是在蛋雞遺傳改良計劃中重點提出了基因組選擇進入實質的應用階段[2]。中國自2004年完成第一個紅原雞全基因組測序之后[3],相繼完成了牛[4]、山羊[5]、豬[6]、草魚[7]、綿羊[8]等家養動物的參考基因組測序工作,在此基礎上通過高通量測序技術的不斷優化提升,一系列與生產密切相關的畜禽基因組變異被充分挖掘出來,進而在DNA分子水平上通過分子生物學手段進行分子標記,為加速現有畜禽品種改良和早期選種提供了強大的技術支撐。理想的基因組變異檢測技術是在全基因組測序基礎上進行的,基于高通量測序技術充分挖掘分子遺傳標記,能夠高效、精準的捕獲生物體大量遺傳變異,挖掘基因資源,促進物種的基礎研究及其上下游產業的發展。將測序技術與常規育種方法相結合,能夠挖掘大量可利用遺傳變異,提高育種的遺傳進展,從而為畜禽種業創新發展提供良好平臺。
1 測序技術的發展歷程及應用領域
1.1 測序技術的發展歷程及應用平臺比較分析
自2003年完成人類基因組計劃后,基因組測序技術取得了長足的進步,這也使測序的成本大幅度下降[9]。在第一代Sanger測序技術的基礎上,相繼又發展起來了二代和三代測序,因二代測序技術具有通量高、成本低、讀長短等特點[10],又被稱為高通量測序技術或下一代測序技術。測序技術經歷3個階段的發展過程:第一階段以1977年Sanger和Walter Gilbert發明傳統測序技術為開端[11],第一臺測序儀誕生標志著大規模的基因序列測定成為了可能,首個人類基因圖譜的繪制就是以改進的Sanger測序技術為基礎產生的,至今,Sanger技術仍然被廣泛應用,但一次測序只能完成700~1 000 bp,無法滿足現代科學技術對挖掘生物基因序列的迫切需求,因此高通量測序技術應運而生;第二階段即第二代測序技術在第一代測序技術的基礎上進行了革新性改造,一次運行測序就能完成幾十萬條至幾百萬條的核酸分子序列,其讀長短、準確性低的缺點帶來測序深度不深和拼接技術的準確性不夠等問題;第三階段針對第二代測序技術存在的上述問題,開發了第三代測序技術,又稱為單分子實時DNA測序技術,該技術在保證通量的基礎上,從單條長度從頭測序,能夠直接得到數百萬個核酸的序列信息。從應用平臺的比較分析來看,三代基因測序技術的應用平臺在不斷地提升測序能力,目前就市場形勢來看,高通量測序技術仍在市場中占有絕對優勢。第三代測序技術也在近幾年的科學研究中快速發展。二代測序技術以羅氏454測序儀(Roche GS FLX Sequencer)、Illumina公司的Solexa基因組分析儀和ABI的SOLiD測序儀為主要的測序平臺。三代測序技術以Pacific Bioscience的SMRT技術、The Oxford Nanopore Technologies公司的納米孔單分子測序技術為主要測序平臺,解決了二代測序讀長短、準確性低的問題。測序技術的應用平臺的優劣勢、測序原理以及應用領域總結如表1。
1.2 測序技術的應用領域
在高通量測序技術的基礎上,通過對物種的個體或群體進行基因組測序或差異分析,從而獲得大量的與生物表型密切相關的遺傳變異,如單核苷酸多態性位點(SNP)[27]、小片段的插入和缺失(Indel)、結構變異(SV)、拷貝數變異(CNV)[28]以及轉座子變異等,用于開發分子遺傳標記,建立遺傳多樣性數據庫,為物種的進化關系、挖掘功能基因、高效精準的早期選擇以及輔助分子育種提供重要的數據支撐。以中心法則為核心的高通量測序技術也演變出很多分支測序技術,如不需要任何參考序列就能對某一物種進行基因組測序的Denovo技術,也稱為從頭測序[29];對已知基因組序列的物種進行不同個體的基因組測序的全基因組重測序技術(WGS)[30];在生物體整個基因組捕獲轉錄因子或組蛋白修飾DNA靶點的全基因組甲基化測序技術(WGBS)等[31]。由于連鎖不平衡的作用,利用全基因組關聯分析(GWAS)分析的目標性狀關聯的序列區域具有較大的寬度,不能精準地逐漸聚焦而定位到目標性狀的核心區域。研究表明,GWAS發現的SNP會影響到附近區域基因的表達量,最終造成鎖定的目標區域不是與生物表型性狀高度相關的區域[32],因此,聯合轉錄組學,從復雜的區域逐步鎖定控制表型性狀的關鍵基因,GWAS和轉錄組聯合分析可以實現結果準確性的相互驗證。一直以來,研究認為轉錄調控受RNA的直接影響,但隨著雞、鴨全基因組測序數據的公布,發現參與基因轉錄調控的編碼蛋白約占全部序列的2%,其余98%是受非編碼蛋白RNA調控[33]。因此,高通量測序技術也產生了轉錄組、全轉錄組、空間轉錄組以及單細胞轉錄組等新技術。隨著轉錄組技術的不斷優化提升,原本被認為是“轉錄噪音”的非編碼序列越來越多被發現,以小RNA(microRNA)、長鏈非編碼RNA(lncRNA)、環狀RNA(circRNA)以及與PiWi蛋白互作的RNA(piRNA)為代表的非編碼RNA在細胞分化、增殖、生長、凋亡等重要的生理過程中發揮著重要作用[34-36]。單一的組學技術在解釋復雜性狀的生物學現象方面往往缺乏系統性,對畜禽生長發育機理、重要經濟性狀功能基因的挖掘都不深入,多組學聯合分析可分別從基因、mRNA、轉錄因子、調控因子、蛋白、代謝路徑等不同層次上進行信息的整合,構建基因互作調控網絡,解析復雜性狀形成的分子機理和遺傳基礎[37]。高通量測序技術在基因組、轉錄組以及多組學聯合分析方面的應用領域如圖1所示。
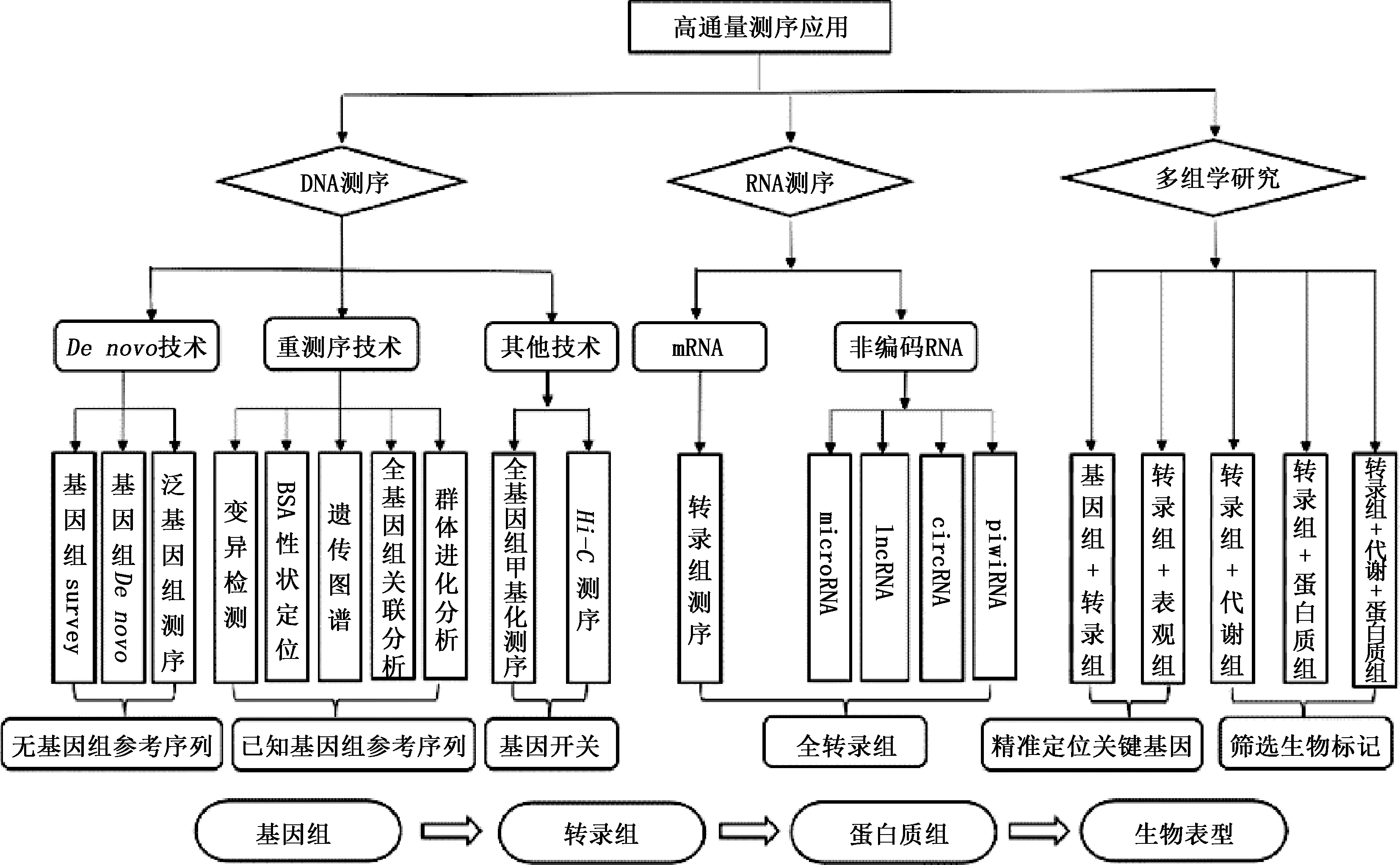
圖1 高通量測序技術應用領域Fig.1 Application fields of high-throughput sequencing technology
高通量測序技術是以中心法則為核心建立起來的技術體系,在中心法則中,DNA水平的基因組學處于最上游,決定了基因的基本屬性,RNA水平的轉錄組學處于中央樞紐地位,但基因不能最終決定生物表型,它還受到表觀遺傳修飾、轉錄、翻譯、轉錄后加工以及環境等因素的影響,并且這些影響和改變最終都體現在代謝物的差異上,代謝組學常被稱作是基因組學和表型之間聯系的紐帶,也是能夠用生物標記物解釋復雜生物性狀最直接的體現[38]。因此,多組學聯合分析也是高通量測序技術的升級版,產生了“基因組+轉錄組”、“轉錄組+代謝組”、“轉錄組+表觀組”以及“轉錄組+代謝組+蛋白質組”,這些組學之間既各自獨立又相互關聯,從不同的角度解析了生物性狀形成的分子機理和遺傳基礎。
2 分子遺傳標記檢測技術在現代蛋雞種業創新中的應用
隨著高通量測序技術的不斷更迭,海量的分子標記被挖掘,分子遺傳標記挖掘的準確性取決于測序深度和組裝拼接的準確性。通過高通量測序技術進行基因挖掘、定位、候選基因的篩選,并對候選目標基因區域SNP、Indel、CNV和SV等基因組變異進行結構注釋和功能注釋,同時,將分子遺傳標記與常規育種手段結合,這樣才能更好地為現代蛋雞種業創新服務。目前,按照遺傳標記檢測技術發展的先后順序,常見的分子遺傳標記檢測技術主要劃分為3個階段,以下針對分子遺傳標記的3個階段進行闡述。
2.1 第一代分子遺傳標記
第一代分子遺傳標記主要包括限制性片段長度多態性(RFLP)、隨機引物擴增多態性(RAPD)和擴增片段長度多態性(AFLP)。RFLP是最早應用于群體遺傳多樣性分析的分子遺傳標記[39],利用限制性內切酶酶切DNA分子后形成特定大小的DNA片段,因其具有共顯性的特點,可以通過電泳條帶直接分析基因型,適合構建遺傳圖譜,同時其具有不受性別、年齡和生長階段限制的優點,因而被廣泛應用于群體多樣性分析、功能基因數量性狀基因座(QTL)多態位點挖掘、傳染性疾病診斷等領域。曹頂國等[40]利用RFLP-PCR技術檢測汶上蘆花雞、濟寧百日雞和萊蕪黑雞極低密度脂蛋白受體(VLDLR)基因的SNP,分析發現VLDLR基因的V8467和V13876的SNP可以作為蛋雞蛋黃性狀的分子標記;邵勇剛等[41]利用RFLP-PCR技術檢測拜城油雞公雞的脂肪型脂肪酸結合蛋白(A-FABP)基因多態性與生產性能相關性的研究發現,在拜城油雞公雞中共檢測到3個A-FABP基因型,分別為AA、AB、BB型,其中BB型為優勢基因型,BB型個體與脂肪性狀相關的生產性能高于AA型個體,且不同年齡不同基因型之間存在差異,這也就進一步解釋了A-FABP基因可以作為拜城油雞公雞的分子遺傳標記用于育種;Parviz等[42]利用RFLP-PCR技術對影響蛋殼質量的蛋殼基質蛋白卵黃蛋白原-32(OCX-32)基因進行基因分型,SNP (c.193A>C)是一種錯義突變(天冬酰胺取代組氨酸),同時也解析了OCX-32基因能夠中止鈣化,與鈣結合蛋白D28K作用相反,影響蛋殼中鈣的沉積量,從而調控蛋殼顏色和蛋殼質量。為了克服RFLP技術上存在的多態位點低、技術復雜、成本高等缺點,RAPD在RFLP技術的基礎上進行優化提升,滲透于基因研究的各個領域,主要原理是隨機設計引物(8~10 bp),通過PCR反應非定點擴增DNA片段,經過凝膠電泳、溴化乙錠染色,結果顯示出擴增片段DNA多態性,該技術具有操作簡單,設計的隨機引物可以用于不同基因組分析,用1對引物就可以擴增出很多片段,具有不需要同位素、安全性好等優點,但也存在準確性和重復性差的缺點。AFLP是在RFLP和RAPD檢測技術基礎上發展而來的,具有RFLP高重復性和RAPD技術操作簡單等雙重優點,它將基因組DNA成對的經限制性內切酶酶切產生的片段用接頭(與酶切位點互補)連接起來,并通過5′-端與接頭互補的半特異性引物擴增得到大量DNA片段,從而形成指紋圖譜的分子標記技術。目前,AFLP技術已經在畜禽遺傳育種中發揮了它的優勢。
2.2 第二代分子遺傳標記
第二代分子遺傳標記主要包括簡單重復序列(SSR)和簡單重復序列間區(ISSR)。在真核生物基因組中普遍存在許多非編碼的重復序列,其大小只有1~6 bp,串聯成簇的長度為50~100 bp,稱為短串聯重復序列(STR),又稱為簡單重復序列(SSR)[43],因其具有多態性(PIC)高、數量多且廣泛分布在基因組中的特點,被廣泛應用在個體及親緣關系鑒定、構建遺傳連鎖圖譜、雜交優勢預測、群體遺傳距離分析以及功能基因定位及QTL定位[44]。每個微衛星兩側一般都是相對保守的單拷貝序列,據此可以涉及專一性引物,通過PCR技術擴增微衛星片段,擴增產物經凝膠電泳分離后,不同個體間因核心序列的重復次數不同而產生DNA多態性。研究表明,隨著SSR堿基的增加,條帶會逐漸減少,多態性的區域就會變得更加明顯,基因分型就越準確[45]。曲魯江等[46]利用28對個微衛星檢測中國地方品種大骨雞和北京油雞2個保種場保種效果比較分析,發現各個群體間均有較高的多態性,雜合度為0.55,各位點等位基因數目為2~22個,除LEI10194和MCW0032外,其余26個微衛星位點都處于基因平衡狀態。伍革民等[47]利用7個微衛星標記對貴州省的4個烏骨雞群體進行群體遺傳結構和遺傳距離的分析發現,4個烏骨雞群體多態信息含量在0.6431~0.6913,雜合度在0.7069~0.7385,當PIC>0.5時,表現為高度多態,4個烏骨雞群體平均雜合度越大,表明群體的遺傳變異程度越高,群體遺傳多樣性越豐富。Hillel等[48]充分利用SSR技術對來自多個國家的52種雞群的群體內和群體間遺傳多樣性進行了評估,發現商業品系的各個位點平均雜合度為0.29,平均群體內雜合度為0.47,而地方品種平均雜合度為0.50,說明在雞群進化過程中地方品種內部由于遺傳漂變等因素的影響,導致了群體遺傳多樣性要比商業品系高,為利用好地方品種遺傳資源提供了新的路徑和新的見解。ISSR技術是在SSR技術基礎上建立起來的加錨核苷酸技術,ISSR技術應用于群體遺傳多樣性研究、親屬關系以及系統發育方面的研究。武玉珍等[49]利用ISSR技術對褐馬雞兩大種群各35只雞進行親緣關系分析發現,個體遺傳相似性的平均值為0.5061和0.7591,遺傳距離平均為0.4939和0.2409,表明在相同生態地理條件下種群的親緣關系較近。ISSR解決了SSR技術在物種間引物設計費時費力的缺點,無需預先克隆和測序,節約減少了試驗前期的準備工作的強度[50]。
2.3 第三代分子遺傳標記
第三代分子遺傳標記主要包括SNP和表達序列標簽(EST)。SNP是DNA上單個核苷酸變異導致DNA序列多態性,大約1 000 bp存在1個SNP,SNP檢測技術是第三代遺傳分子標記中最具應用價值的分子標記技術[51],常見的有堿基的轉換、顛換、缺失或替換導致的DNA序列多態性[27]。SNP分子標記因其密度高、遍布基因組、易于實現自動化等優點被廣泛應用[52],但僅因1個位點的SNP變異很難影響到基因的表達和編碼蛋白序列,進而影響到表型的差異。2014年Carneiro等[53]在研究家兔馴化遺傳機制方面發現,通過全基因組測序技術共發現5 000萬個SNPs,但真正能影響到氨基酸編碼的錯義突變只有7萬個SNPs,在馴化過程中保留下來的錯義突變只有14個SNPs,說明SNP分子標記技術也有其局限性。對于挖掘SNP主要取決于采用的SNP研究策略,目前主要采用低通量的實時熒光定量PCR和高通量測序技術制備SNP芯片來挖掘SNP。針對SNP具有數量大且只有一小部分屬于關鍵功能變異的特點,SNP的研究和應用進入了瓶頸階段,目前,CNV成為了繼SNP之后的關注熱點,主要是因為CNV屬于大片段結構變異,覆蓋較大區域的基因組序列,甚至能夠影響基因序列的表達[54],一些重要的CNV已經在人[55]、牛[56]、羊[57]、豬[58]、雞[59]等上被挖掘,并表現出與一些數量性狀和質量性狀形成的遺傳機制高度相關,可以作為一種繼SNP分子標記之后的另一個理想的分子遺傳標記,用于畜禽育種。EST是指來源不同的組織的cDNA文庫,隨機挑選克隆、測序得到的cDNA序列,EST可以代表生物體某種組織某一時間的1個表達基因,目前來看,EST標記技術廣泛應用于植物基因組研究,關于應用EST標記技術在動物基因組研究中鮮見報道。
3 高通量測序技術在蛋雞生產中的精準應用
3.1 群體遺傳多樣性及進化分類
利用分子遺傳標記在評價群體遺傳多樣性方面,很多研究都集中在運用雜合度(He)、純合度(Ho)、有效等位基因數(Ne)以及PIC等指標來評價。在評價群體進化方面,尤桂爽等[60]利用簡化基因組GBS測序方法對四川盆地6個雞群遺傳關系進行了研究,利用He、PIC以及核酸多態性(Pi)等指標評價了6個雞群的遺傳多樣性,巖水雞黑羽群體的遺傳多樣性較低(He=0.154,PIC=0.125),大寧河雞遺傳多樣性最為豐富(He=0.197,PIC=0.162)為低多態性,雜合度和多態信息含量是衡量群體標記多態性的重要指標[61],四川盆地的6個地方雞的He范圍在0.154~0.197,這也和Cendron等[62]研究意大利6種地方雞的研究結果一致。韓威等[63]利用簡化基因組測序技術(RAD-Seq)對19個地方雞種進行基因組SNP標記,在19個雞種中共鑒定出400 562個SNPs,進一步證實了中國地方品種遺傳多樣性豐富,利用He、Pi、近交系數(Fis)、遺傳分化系數(Fst)基因流(Nm)評價了19種地方雞的遺傳多樣性和遺傳分化情況,瓢雞和文昌雞的遺傳多樣性最為豐富,Ho分別為0.2468、0.2430,Pi分別為0.2781、0.2655;河南斗雞的遺傳多樣性相對匱乏,Ho為0.1560,Pi為0.1752;東鄉綠殼蛋雞和邊雞的近交系數最高(Fis>0.1600)。瓢雞與文昌雞、惠陽胡須雞、藏雞、大圍山微型雞,惠陽胡須雞與文昌雞,藏雞與茶花雞間的遺傳分化最低(Fst<0.1000),對應的基因流最高(Nm>0.2400),為中國地方雞品種資源保護、開發及利用提供了技術支撐。
3.2 群體遺傳圖譜的構建和功能基因定位
利用高通量測序技術挖掘分子標記,很難定位到目標性狀的候選區域,因此對分子遺傳標記的選擇顯得十分重要。遺傳圖譜能夠縮小分子標記的數量,可以根據專一分子標記或基因在染色體上的相對位置排列而成,遺傳圖譜是全基因組關聯分析成功的一個重要的前提。在雞的NCBI SNP數據庫中有超過900萬個SNPs標記[64],雞的遺傳圖譜的相對長度為3 800 cM,在雞的基因組中由10億個堿基組成[3],大約200 bp存在1個SNP突變位點[65]。吳丹等[66]對北京油雞的生長性狀進行關聯分析,利用雞的60K SNP芯片對其28、56、80和100日齡的體重進行GWAS,發現影響北京油雞體質量性狀的全基因組關聯位點為9個,潛在關聯位點為96個,找到了LIM結構域結合基因(LDB2)、LYR基序含子基因(LYRM1)、配體依賴性核受體輔阻遏物樣基因(LCORL)等候選基因,這些候選基因和分子標記為北京油雞種質資源的評價、利用及保護提供了育種素材。孫艷發等[67]利用60K SNP芯片對雞的體重、脛長和脛圍進行全基因組關聯分析,達到5%顯著性全基因組關聯位點分別有1和4個,潛在關聯位點分別為4和25個,分布在4號染色體14.93 Mb區域的SNP位點與脛長和脛圍有著顯著或潛在的聯系,這個區域內含有LIM結構域結合因子2(LDB2)、細胞分裂染色體雙軸取向1-樣1(BOD1L1)以及醌式二氫蝶啶還原酶(QDPR)等候選基因,8號染色體上有1個SNP位點位于ELA樣蛋白4(ELAVL4)基因下游的52 248 bp處,13號染色體上有1個SNP位點位于同源異型盒蛋白Nkx-2.5(NKX2.5)基因下游的138 834 bp處,這些候選位點都為解析脛長和脛圍的遺傳機制提供了新思路。Julien等[68]利用全基因組測序技術解析了日本鵪鶉的社會行為、侵略性等指標,利用2 145個分子標記構建了其遺傳圖譜,其中1 479個標記可定位在不同的28個連鎖群上,平均遺傳連鎖圖譜長度為3 057 cM,平均標記距離為2.1 cM,在QTL定位結果中發現45個QTLs,其中22個與行為有關,23個生產性能相關,這些QTL定位區域的發現也為優化動物福利模式提供了遺傳機制上的新見解。因此在蛋雞生產中應用分子標記進行分子育種、構建遺傳圖譜、定位功能基因,有助于加快新品種育成速度和解決蛋雞產業“卡脖子”關鍵技術。
3.3 數量性狀形成的遺傳機制解析
吳常信等[69]在基于國外育種公司提出的“100周產500枚蛋”計劃是否能實現,進行了可行性的分析,雖然這種設想在2020年實現“100~500”計劃是不可能的,但也對中國蛋雞產業具有很強的借鑒意義,確定了“提高80~100周齡的產蛋率”和“增強產蛋后期蛋品質選擇”是中國蛋雞育種的目標。在這2個育種目標的指引下,很多研究人員都進行了有益的探索。在增強蛋品質方面,現在大多采用全基因組關聯性分析和分子標記輔助選擇技術進行了大量的研究,對雞的初生重以及48和60周齡的產蛋量和蛋黃顏色,使用全基因組關聯性分析,獲得了26個與蛋品質顯著相關的SNP位點[70];用高密度陣列的全基因組關聯研究對東鄉藍殼雞與白來航雞雜交產生的F2代資源群中獲得的禽蛋,通過雙性狀動物模型和基因注釋獲得了蛋殼的候選基因,分別是調節突觸膜胞吐2(RIMS2)、溶質載體家族(SLC25A32)、液泡分選蛋白13B(VPS13B)和G蛋白信號調節器3(RGS3)[71];結合分子標記輔助選擇技術,為同時表現藍蛋殼和矮化性狀的純種雞提供了一種快速育種方法[72]。目前在蛋品質研究中所產生的候選基因、大量的QTL和致病基因,但由于分子標記的遺傳效應過小,所能解釋表型差異的生物學現象有限,分子輔助標記選擇至今沒有一個完整的、成功的育種方案,導致這些成果在商業品系中并不能得以應用[73]。因此,在后續的研究中著重于對候選基因重要功能SNP位點的挖掘,以及調控這些功能基因的上下游信號通路的生物學機制的研究應用到蛋雞品系培育中,更好地為家禽的早期選育以及重要經濟性狀應用奠定基礎。在提高產蛋率方面,家禽產蛋受環境、內分泌、遺傳因素的共同調節。隨著分子遺傳學技術和DNA分子標記的出現,家禽產蛋性能相關性狀的QTL用于分子標記輔助選擇的研究迅速發展[74],現多使用高通量測序,利用分子育種策略,可以使家禽養殖行業最大限度地降低生產成本。采用RNA-Seq測序技術,對蘆花雞下丘腦、垂體和卵巢組織進行了轉錄組分析,從15個文庫中產生了76.09 Gb的RNA-Seq序列,每個文庫的平均大小為5.07 Gb[75];Liu等[76]使用線性混合模型對1 078只洛島紅雞進行了全基因組關聯研究篩選出大量的候選基因,但并沒有發現任何影響總蛋數的候選基因。Wu等[77]對抗繆勒管激素(AMH)外顯子設計引物并擴增,發現京海黃雞AMH基因第二外顯子的多態性及其與繁殖性狀之間的關系,并檢測出4個SNP位點為家禽育種提供了分子標記區段;Wolc等[78]通過使用品系特異性50K Affymetrix Axiom SNP芯片進行全基因組關聯分析,發現了參與卵母細胞成熟的Wiskott-Aldrich綜合征蛋白和富含脯氨酸同源物(WASH1)、調節卵泡刺激素分泌的神經肽VF前體(NPVF)及參與卵泡的卵母細胞成熟和排卵的叉形頭轉錄因子3(FOXO3);Rowland等[79]使用SNP芯片進行GWAS關聯分析發現,3個候選基因膜泡蛋白分揀36(VPS36)、雄激素受體(AR)和無翅型小鼠乳房腫瘤病毒整合位點家族(WNT11B)在卵泡發育、性腺生長和真皮發育方面起重要作用;Azmal等[80]發現,京紅雞3號和13號染色體上的12個SNPs與產蛋后期受精率顯著相關。因此,利用高通量測序技術挖掘與產蛋性狀相關基因的SNP變異位點,并對其進行分子標記,能夠在蛋雞生產中進行高效選種,提高選種強度,進而實現較快的遺傳進展。
3.4 質量性狀形成的遺傳機制解析
針對冠型、羽色、蛋殼顏色、矮腳等具有代表性的性狀,運用高通量測序技術解析蛋雞的質量性狀形成的遺傳機制,能夠在蛋雞早期根據羽速或羽毛特征來鑒別雌雄、進行高效選種,減少飼養成本給育種帶來的經濟負擔。控制質量性狀的等位基因有不完全顯性和完全顯性的現象,對于不完全顯性的性狀,能從表型中直接分辨基因型,根據表型選擇也就是基因型選擇從而達到高效選種的目的。有研究利用全基因組測序技術對東鄉雞、盧氏蛋雞和綠殼蛋雞Araucana進行測序,發現了有機陰離子轉運家族成員(SLCO1B3)基因上游啟動子區域中部分內源逆轉錄病毒(EAV-HP)的插入導致雞綠殼蛋的形成有關,是由于逆轉錄病毒片段會導致相鄰基因的異位表達或異常表達,從而影響SLCO1B3基因的表達[81-83]。Feng等[84]利用全基因組測序技術揭示了雞絲羽性狀是由癸二烯二磷酸酶亞基2(PDSS2)順式作用調控引起,并對根據該位點建立了絲羽性狀的分子標記用于指導生產。Wright等[85]利用高通量測序技術揭開了雞豆冠形成是由于SRY轉錄因子5(SOX5)基因的一段非編碼序列拷貝數增加而引起,雞冠性狀對于高效選種也十分重要。Cui等[72]利用分子輔助標記選擇策略,對綠殼蛋和矮腳2個性狀進行選擇,培育出綠蛋殼矮腳性狀的純種雞品系,就是應用SLCO1B3和生長激素受體(GHR)基因的分子標記技術培育而來。Luo等[86]利用組織切片、全基因組關聯研究和RNA-Seq技術,進一步研究了羌源麻雞紅/白耳垂形成的組織學和分子遺傳學機制,這些利用質量性狀快速培育雞的新品系的新方法和新視野,為利用蛋雞基因組高效選種進入實質階段提供了技術支撐。
4 小 結
隨著測序技術的不斷更新換代,以新一代測序技術為基礎發展起來的高通量測序技術和單分子實時測序技術都為挖掘基因變異做出了重要貢獻。其中,高通量測序技術是現如今研究物種遺傳變異形成機制的應用最為廣泛的測序技術,高通量測序技術及分子遺傳標記檢測技術在蛋雞生產中的精準應用主要有4個方面:①利用高通量測序技術能夠挖掘與蛋雞產蛋性能表型性狀高度相關的基因組變異,并對這些變異進行檢測,精準定位與產蛋性狀相關的候選區域,形成分子遺傳標記,利用基因組選擇的遺傳變異信息對蛋雞群體或個體的遺傳基礎進行評定,從而達到縮短世代間隔,提高選種的準確性,實現較大的遺傳進展;②在進行分子遺傳標記篩選和功能驗證過程中,利用多組學技術縮小定位產蛋性狀候選性狀區域范圍,更加精準的挖掘產蛋性狀的功能基因;③利用多種技術反復驗證目標性狀的候選基因,從而多維度的解析生物性狀形成的機制,為蛋雞基因組選擇進入實質階段提供技術參考具有重要指導意義;④利用分子遺傳標記技術進行基因分型,選取樣本進行PCR擴增產物測序,而后分析各個基因型對產蛋性狀的遺傳效應,本質上分子遺傳標記檢測技術是高通量測序技術的補充。目前應用高通量測序技術和分子遺傳標記檢測技術已經成功將控制綠殼性狀的SLCO1B3基因應用于綠殼蛋雞的品種培育上,將矮小基因應用于節糧型蛋雞的培育上,因此,該技術助推了蛋雞產業高質量發展。高通量測序挖掘分子遺傳標記及其在蛋雞生產中精準應用如圖2所示。
雖然新一代測序技術在挖掘蛋雞基因組變異上有諸多優勢,但也存在著定位功能主效基因不準確、實際應用存在著諸多問題,因此,未來測序技術在蛋雞生產中的精準應用,一方面要圍繞開發中等密度SNP芯片用于分析雞的重要經濟性狀和利用地方雞資源培育新的蛋雞新品系,解決中國蛋雞品種“卡脖子”關鍵問題,另一方面用圍繞運用多種新興技術諸如染色質可及性分析(ATAC)、高通量染色體構象捕獲技術(Hi-C)、單細胞測序技術等,更深層次解釋表型差異的生物學現象,為解析蛋雞重要經濟性狀形成機理提供技術支撐。
