斑馬魚模型在評價多糖類成分增強免疫活性研究中的應用
2022-10-26 10:35:40王海強胡瑞雪文興建
重慶理工大學學報(自然科學) 2022年9期
王海強,饒 唱,曾 易,胡瑞雪,文興建,勵 娜,胡 光
(1.重慶理工大學 藥物化學與分子藥理學重慶市重點實驗室, 重慶 400054;2.重慶市中藥研究院, 重慶 400065)
1 概述
斑馬魚的免疫系統發育的比較完備,同時具備固有免疫與獲得性免疫;而其固有免疫系統可以區分自我和非自我rRNA[5],利用斑馬魚可以建立很多人類病原的感染模型及水生動物病原感染模型。斑馬魚免疫系統發育較早,在胚胎形成之后的1 d內,就可以檢測到固有免疫反應。ELAVL1a是一種儲存在早期胚胎中的膜磷壁酸(lipoteichoic acid,LTA)結合蛋白,它也是一種母體免疫因子,可以保護斑馬魚胚胎免受細菌感染[6]。腎臟作為斑馬魚的主要造血器官,相當于哺乳動物中的骨髓,其中含有T細胞、B細胞及樹突狀細胞(dendritic cells,DCs)等[7]。巨噬細胞專門識別調理病原體和非健康細胞(如凋亡和壞死細胞 )發出的信號。它們在固有免疫系統中發揮重要作用,可以直接來源于卵黃囊和胚胎肝臟,也可以來源于循環單核細胞。在炎癥條件下,單核細胞在組織中轉化為巨噬細胞。此外,巨噬細胞主要可分為:① 經典活化的巨噬細胞(M1),它們是宿主用來抵御病原體的介質,還具有抗腫瘤免疫作用[8]。② 替代活化的巨噬細胞(M2),具有抗炎和促進傷口愈合功能[9]。為了開發疫苗、治療一些慢性炎癥性疾病[10-11]和重新培養巨噬細胞以進行新的癌癥治療[12-13],人們有意追求免疫細胞的特異性和靶向性。細胞因子和趨化因子是免疫系統重要的組成部分。斑馬魚的相關因子已陸續得到分子鑒定,并顯示出與其他魚類的因子有很高的相似性。白細胞介素-6(interleukin-6,IL-6)是一種高度通用的細胞因子,不僅在免疫細胞中具有多效性作用,而且在其他細胞類型中也具有多效性作用;此外,已經證明了IL-6還參與多種生理中樞神經系統過程,如神經元穩態,星形膠質細胞生成和神經元分化[14]。白細胞介素-8(interleukin-8,IL-8)是在炎癥中最有效的嗜中性粒細胞化學誘導物之一,在其他幾種功能中,負責引導中性粒細胞穿過組織基質到達損傷部位;而斑馬魚可能擁有一個中性粒細胞亞群,其在炎癥區域的募集獨立于IL-8趨化因子[15]。干擾素在哺乳動物對病原體的抵抗中起重要作用,對于病原體的抵御與清除、控制腫瘤的滋生等免疫反應至關重要[16]。干擾素-γ(interferon-γ,IFN-γ)是一種T輔助細胞1型(Th1)細胞因子,在免疫和炎癥反應的幾乎所有階段都發揮重要作用,IFN-γ可能參與炎癥反應,并與哺乳動物的干擾素一樣促進Th1免疫反應[17]。白細胞介素-10(interleukin-10,IL-10)由多種細胞產生,是一種高度多效性的細胞因子;盡管它有時也顯示出一些似乎相互矛盾的差異效應[18],但是它仍被證明可能與自身免疫性疾病和癌癥的發病機制和發展相關。
近年來,很多學者的研究表明,植物多糖對魚類的生長、抗菌抗病、免疫、抗氧化和降血糖血脂等方面有積極作用[19-21];也在生物體和細胞的生命活動中發揮著重要作用,具有廣泛的生物活性。例如:在飼料中添加膳食殼聚糖硒納米顆粒(CTS-SeNP)可顯著增強斑馬魚的免疫力和抗病性[22];β-葡聚糖不僅能提高斑馬魚免疫能力,還能改善營養不良斑馬魚的運動性能和線粒體功能[23]。更有趣的是,現已證明β-葡聚糖具有跨代提高免疫力的作用,β-葡聚糖可以通過誘導溶菌酶分子從母體轉移到卵子,并刺激其在卵母細胞中的內源性生產,從而提高后代的溶菌酶水平[24]。此外,多糖很有可能不會在腸道通過消化吸收的途徑發揮功能,而是直接激活免疫信號通路,刺激免疫細胞的活化[25],或者通過改善腸道菌群結構,提高魚類的免疫力[26]。
綜上所述,斑馬魚在藥物活性的研究中有著諸多的優點,包括,早期的胚胎為透明狀態,便于連續觀察胚胎發育的全過程等,適用于高通量篩選[27]。同時,作為脊椎動物,斑馬魚免疫系統發育的比較完備,同時具備固有免疫與獲得性免疫;加上野生型斑馬魚維護與實驗成本比轉基因型斑馬魚低了很多,使其更適合應用于較多數量藥物的初篩過程。本文以經濟的野生型斑馬魚為主要對象,總結了該模型在評價多糖的免疫促進作用的研究,并按照多糖的種類、多糖給藥方式及其免疫活性評價方法、免疫活性評價用病原菌、研究涉及的信號通路等幾方面,進行了分類說明。
2 采用斑馬魚模型評價多糖的免疫促進作用
2.1 多糖的種類
目前已有大量的天然多糖在科研和生活中被應用,例如常用于科研的甜菊糖、白芷多糖、榴蓮果皮多糖、川芎多糖、枸杞多糖等。
Snega 等[28]以榴蓮果皮為原料,提取多糖凝膠(PG),并用PG與枯草芽孢桿菌、豐年蝦幼蟲(artemia nauplii)分別組合后飼養斑馬魚,然后研究斑馬魚在弧菌浸泡激發(5 d)后誘導的免疫應答的效果。總紅細胞計數、溶菌酶活性測試、體重和長度分析表明,與對照組相比,喂食PG的斑馬魚具有更高的免疫誘導活性、存活率和生長率。當檢測免疫相關基因的表達時,研究顯示與其他飼料相比,用含有PG的豐年蝦喂養的魚類的IL-1β、溶菌酶(lysozyme,Lyz)、腫瘤壞死因子-α(tumor necrosis factor,TNF-α)、超氧化物歧化酶(superoxide dismutase,SOD)的基因表達顯著上調(P<0.05)。
甜菊糖是一種具有藥用功能的甜味劑,因其獨特的生理功能而在食品和醫藥領域受到廣泛關注[29]。SRP70-1是甜菊糖的主要有效成分,由甘露糖、葡萄糖、半乳糖和阿拉伯糖組成,摩爾比為1.15∶1.00∶3.77∶4.45,絕對分子量為7 698 Da。SRP70-1能顯著促進細胞凋亡、吞噬并增加一氧化氮和細胞因子(包括IL-1β、IL-6和TNF-α)的釋放,進一步的斑馬魚實驗證實了SRP70-1的免疫增強作用[29]。
Sacks等[30]從白芷中獲得水溶性多糖ADP80-2。ADP80-2由阿拉伯糖和微量葡萄糖組成,分子量為9950 g/mol。ADP80-2可以顯著提高巨噬細胞的吞噬功能,促進一氧化氮(NO)的產生和細胞因子(IL-6、IL-1β和TNF-α )的分泌。在用ADP80-2處理后的斑馬魚模型中,蛋白質含量顯著增加。這些生物學結果表明:ADP80-2具有免疫調節作用,有望被用于開發成為新的免疫調節劑。
張紹杰等[31]首次從川芎中分離到一種具有免疫作用的阿拉伯糖LCP70-2A。體外細胞實驗表明,LCP70-2A可以通過增強吞噬功能和增強免疫調節因子(包括NO、TNF-α、IL-6和IL-1β)的分泌來激活巨噬細胞。此外,使用斑馬魚模型發現LCP70-2A可促進ROS和NO的釋放,這表明LCP70-2A可進一步開發為免疫增強的候選補充劑。在該團隊尋找生物活性多糖作為免疫調節劑的過程中[32],從藤黃皮中純化并表征了一種阿拉伯呋喃多糖(GMP90-1),該多糖(絕對分子量:5 300 g/mol )由阿拉伯糖、半乳糖和鼠李糖組成。構象分析顯示GMP90-1在氯化鈉溶液中以剛性棒狀結構存在。為了探索其作為免疫調節劑的潛力,進行了體外細胞篩選,發現GMP90-1可顯著增強巨噬細胞的吞噬能力,并提高巨噬細胞的NO、IL-6、IL-1β和TNF-α的分泌水平。此外,斑馬魚體內實驗證實了GMP90-1的細胞免疫調節活性,這表明具有免疫調節作用的GMP90-1可被認為是一種潛在的免疫調節劑,用于治療免疫性疾病。
(1)投資估算。本工程總投資估算:2504.78萬元,其中建筑工程投資1684.06萬元,機電設備及安裝工程投資2.90萬元,金屬結構設備及安裝工程投資81.41萬元,施工臨時工程投資171.63萬元,獨立費用投資257.62萬元,基本預備費219.76萬元,建設征地移民補償投資39.13萬元,水土保持工程投資38.06萬元,環境保護工程投資10.21萬元。
β-葡聚糖(β-glucans)是從酵母細胞壁中發現的葡萄糖聚合物。在哺乳動物中,β-葡聚糖通過Toll樣受體(toll-like receptors,TLR)TLR2/TLR6異二聚體[33]和小膜受體Dectin-1的識別激活固有免疫。體內注射β-葡聚糖可增強對寄生蟲、細菌和病毒病原體的抵抗作用[34-35]。在飼料中添加β-葡聚糖喂養是向魚類補充β-葡聚糖的最常見方式,該方式增加了固有免疫和獲得性免疫應答的一些關鍵基因的轉錄水平[36]。然而,很少有文獻研究β-葡聚糖喂養后,斑馬魚對病毒挑戰的應答情況;但有文獻報道顯示,用β-葡聚糖的飼養的太平洋鯡魚對病毒性出血熱病毒(VHSV)挑戰表現出更強的抵抗力[37]。另一種葡聚糖應用方式,即單劑量注射,也被用于增強對疫苗的免疫反應,作為提高對病毒的抗病性的有效手段[38]。
胞外多糖(exopolysaccharides,EPS)是由紅色微藻合成的一種有價值的代謝產物。它是一種無毒的天然藥物,可作為免疫刺激劑使用。Risjani等[39]利用斑馬魚胚胎模型在體內進行了卟啉胞外多糖的毒性試驗,隨著EPS濃度的增加,總血細胞(THC)值、吞噬活性(PA)和呼吸爆發(RB)都有所增加。結果表明,卟啉菌胞外多糖是安全的,能夠快速提高斑馬魚的免疫參數,并具有作為免疫刺激劑或免疫調節劑的能力,可作為弧菌病的預防劑。
枸杞(L.barbarum)果實或提取物是一種常見中藥,用于調節身體免疫力和抗衰老,枸杞多糖(LBPs)一直以來被認為是其有效成分。Xia等[40]將斑馬魚胚胎連續暴露于不同濃度的LBPs中3 d,熒光吖啶橙和SA-β-gal染色結果表明,受精后24 h和72 h,細胞凋亡和衰老主要發生在頭部。此外,在低劑量LBPs下,尤其是在3.0 mg/mL濃度下,觀察到對復制性衰老的抵抗。與衰老相關的基因,如p53、p21和Bax的表達在LBPs治療后降低,而Mdm2和TERT基因的表達在LBPs治療后增加。結果表明,LBPs對細胞凋亡和衰老的影響可能是通過p53介導的途徑實現的。LBPs通過抑制發育早期的細胞死亡和凋亡,降低p53、p21和Bax基因的表達水平,增加Mdm2和TERT基因的表達,緩解了斑馬魚的衰老[40]。
巖藻依聚糖來源于褐藻的富含巖藻糖的硫酸化多糖。來自不同藻類來源的巖藻依聚糖表現出獨特的性質,例如抗炎、抗過敏、抗腫瘤或抗病毒作用[41],并因此被認為是功能性食品和治療劑的預期成分[42]。Wakako等[43]分別通過浸泡(100和500 μg/mL,3 d)和喂食(3周)處理斑馬魚幼魚和成年斑馬魚。褐藻糖膠處理僅輕微影響幼魚體內微生物群的組成以及中性粒細胞和巨噬細胞的數量,用500 μg/mL褐藻糖膠處理的幼魚中,促炎和抗炎細胞因子基因表達水平上調,喂食褐藻糖膠則明顯改變了成年斑馬魚的腸道微生物組成[43]。揭示了在活生物體內巖藻依聚糖對未受攻擊的斑馬魚的微生物群和免疫反應的調節作用,這強調了巖藻依聚糖在飲食-微生物群-宿主相互作用中發揮調節作用的潛力。采用不同種類的多糖提高斑馬魚免疫活性的研究如表1。
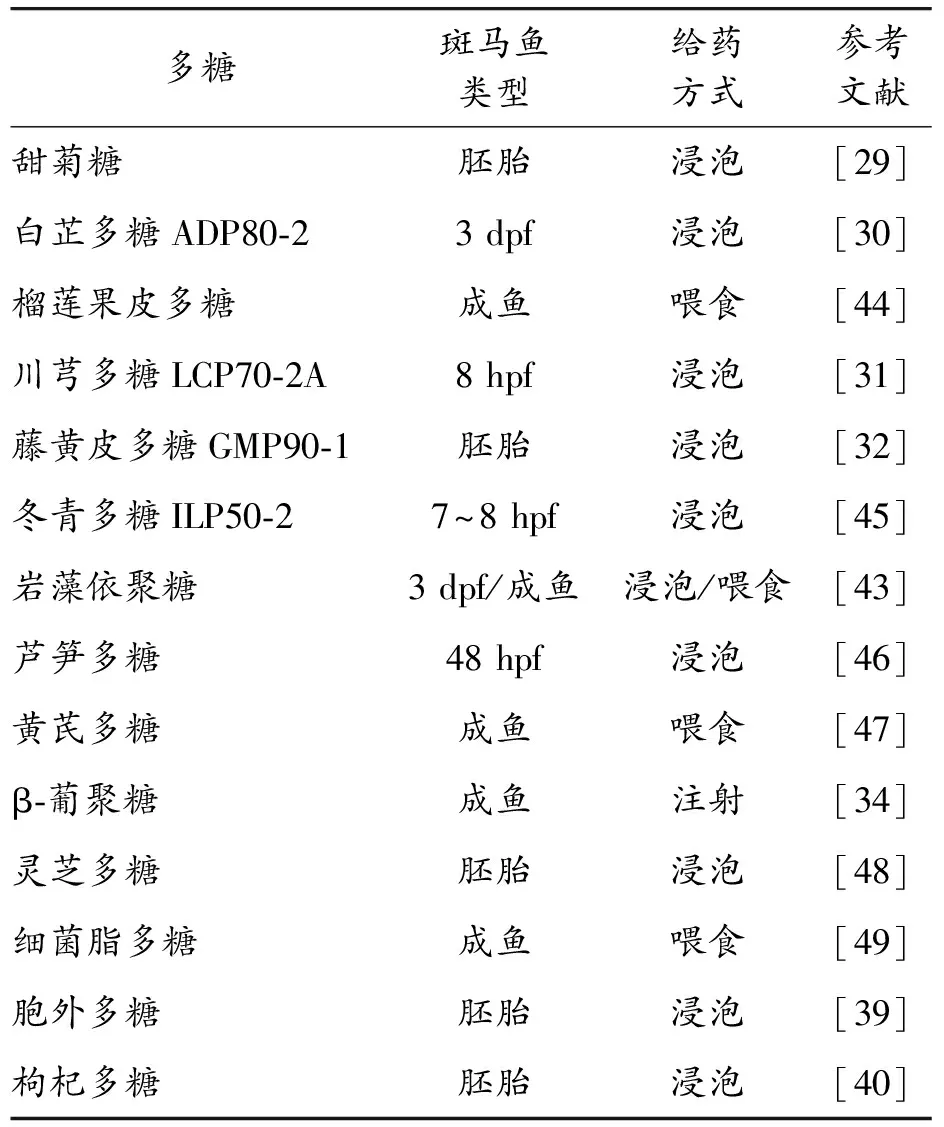
表1 采用不同種類的多糖提高斑馬魚免疫活性的研究列表
2.2 多糖給藥方式及其免疫活性評價方法
斑馬魚的免疫給藥方式大多數仍以注射為主,也有用喂食配制的多糖飼料,和用多糖溶液浸泡的方式進行的。但通過人工注射法進行免疫,不僅人工和時間成本較大,且容易引起魚體強烈的應激反應,影響魚體生長。故浸泡免疫是能普遍被接受的免疫接種方式[50]。Regla M等[38]研究了免疫刺激劑β-葡聚糖的抗病毒作用,在腹腔注射β-葡聚糖的斑馬魚中檢測到與固有免疫反應有關的IL-1β、IL-6、IL-8、IL-10和TNF-α轉錄物的表達增加。Wang等[24]通過向斑馬魚投喂Miero魚類食品或添加含1.25 g/kg β-1,3葡聚糖的魚類食品,證明了β-葡聚糖可以提高幾種魚類的固有免疫和抗感染或抗致病菌的能力。Oyarbide等[51]為了評估β-葡聚糖對基因表達的影響,斑馬魚胚胎和幼魚被浸泡在3種不同濃度的β-葡聚糖(50、100和150 μg/mL )中,且浸泡不同的暴露時間。當暴露時間較長且使用年齡較大的幼魚時,觀察到更強的基因誘導。β-葡聚糖最明顯的作用是誘導TNF-α、MPO、TRF和LYZ基因的過度表達。此外,研究還發現在β-葡聚糖浸泡后,斑馬魚對鰻弧菌的抗性暫時增加。這表明不同給藥方式都能提高斑馬魚的免疫水平,但浸泡接種方式被認為與病原感染類似,接近自然環境中魚類免疫反應激活的方式,并能有效刺激魚類的黏膜免疫應答,產生良好的免疫保護力[50]。在浸泡系統中使用黏液黏附納米載體是給藥和免疫調節劑的一種替代方法,因為納米系統可以黏附在魚的黏膜表面,對水的殘留影響很小[52]。此外,浸泡在不同的納米藥物中也有很大區別:Crecente-Campo等[53]將斑馬魚暴露于4種納米系統的5種不同濃度溶液中,證明了納米范圍內的微小變化能導致免疫細胞和生物分布特征的顯著變化。是否能成功應用多糖治療魚類疾病,取決于能否確定每個物種的最有效劑量和給藥時間[54]。
總之,與沒有給藥的斑馬魚相比,多糖給藥后能顯著提高斑馬魚的抗菌、抗病毒能力。此外,從文獻的報道數量來看,通過細菌感染來評價多糖對斑馬魚免疫系統增強的文章數量少于未采用細菌未感染來評價的文章數量,大部分研究直接通過檢測多糖給藥后斑馬魚體內的免疫水平來給出結論。
目前,評價多糖促進免疫活性的指標的相應檢測手段最主要為實時熒光定量PCR(quantitative real-time-PCR,qRT-PCR)、蛋白質印跡 (western blot,WB)、酶聯免疫吸附測定(enzyme linked immunosorbent assay,ELISA)等。同時,檢測部位是以斑馬魚腸道組織為主,其次是肝臟、腎臟或斑馬魚全身。Snega等[44]使用RNA iso plus溶液均質組織(5條/缸),從中腸提取總RNA,并通過紫外可見分光光度計定量。使用PrimeScript TM II逆轉錄酶試劑盒將總RNA(1 μg)轉化為cDNA。在Applied Biosystems Step One儀器中,使用SYBR GreenTM qPCR master mix測量基因表達。 Oyarbide等[51]通過RT-qPCR檢測與免疫系統相關的某些選定基因的差異表達,表明受精后72 h的幼魚處于評估所選基因表達的最佳發育階段。Blanca 等[55]使用WB分析檢測治療后斑馬魚血漿中的重鏈蛋白A-瓊脂糖珠和抗Ig M斑馬魚抗體。
2.3 免疫活性評價用病原菌
微生物的致病機理與宿主、病原和環境這3個要素息息相關。斑馬魚模型作為研究宿主與病原之間關系的模式動物在免疫學中得到了廣泛應用,許多重要病原菌的致病機理通過斑馬魚模型也得到了深入的研究[56]。通過斑馬魚篩選有抗菌作用的飼料添加劑,可以預防這些細菌所引發的魚類疾病[57]。所以使用斑馬魚建立細菌侵染模型,能很好地用于研究硬骨魚類的抗病毒免疫應答[56-58]。嗜水氣單胞菌是一種人畜共患病原體,對經典類抗生素具有高水平的耐藥性。噬菌體或原噬菌體編碼的裂解酶已顯示出對抗病原菌的潛力。 Wang 等[59]從嗜水氣單胞菌D4中鑒定出一種完整的原噬菌體(命名為phAhD4)。phAhD4在所有10個已發表的嗜水氣單胞菌序列型(ST)251菌株中高度保守,并且是ST251菌株所特有的。通過原核表達獲得由phAhD4編碼的獨特內溶素PlyD4。在此基礎上,運用斑馬魚模型證明了PlyD4在體外對多種細菌具有殺菌活性,包括嗜水氣單胞菌、維氏氣單胞菌、副溶血性弧菌、銅綠假單胞菌等。斑馬魚在分枝桿菌引起的感染動物模型中贏得了一席之地。這種模型宿主特別有助于研究肉芽腫的作用,肉芽腫是分枝桿菌疾病的炎癥病變特征。在固有免疫的背景下,這種光學透明的斑馬魚幼魚為肉芽腫發展的初始階段提供了一個窗口。 Varela 等[60]使用熒光染料和轉基因標記物使固有免疫機制(如自噬和炎癥體)如何在感染的巨噬細胞中被激活實現了實時可視化。影像學、遺傳學和化學方法的結合表明,巨噬細胞和分枝桿菌之間的相互作用是組織播散和肉芽腫發展的主要驅動因素,而中性粒細胞在早期肉芽腫中具有保護作用。人類和斑馬魚之間保存的不同趨化因子信號軸已被證明可招募允許分枝桿菌生長的巨噬細胞,控制其殺微生物能力,推動其擴散和聚集,并介導肉芽腫血管生成。最后,用斑馬魚幼魚來探索細胞的死亡過程,這也是肉芽腫擴張的關鍵因素。
除上述的幾種病原菌外,斑馬魚也被應用于其他的病原菌研究中(表2)。采用綠色熒光蛋白GFP標記的鰻弧菌感染斑馬魚的模型闡明了腸道及皮膚在鰻弧菌感染魚類中的作用[61],而該菌感染斑馬魚后體內免疫相關基因的轉錄水平變化也已經被研究[50,62]。銅綠假單胞菌(pseudomonas aeruginosa)的三型分泌系統在致病性中的作用以及斑馬魚體內的囊性纖維化跨膜傳導調節蛋白(cystic fibrosis transmembrane conductance regulator,CFTR)對該菌的抵抗作用也得到了很好的闡述[63-64]。
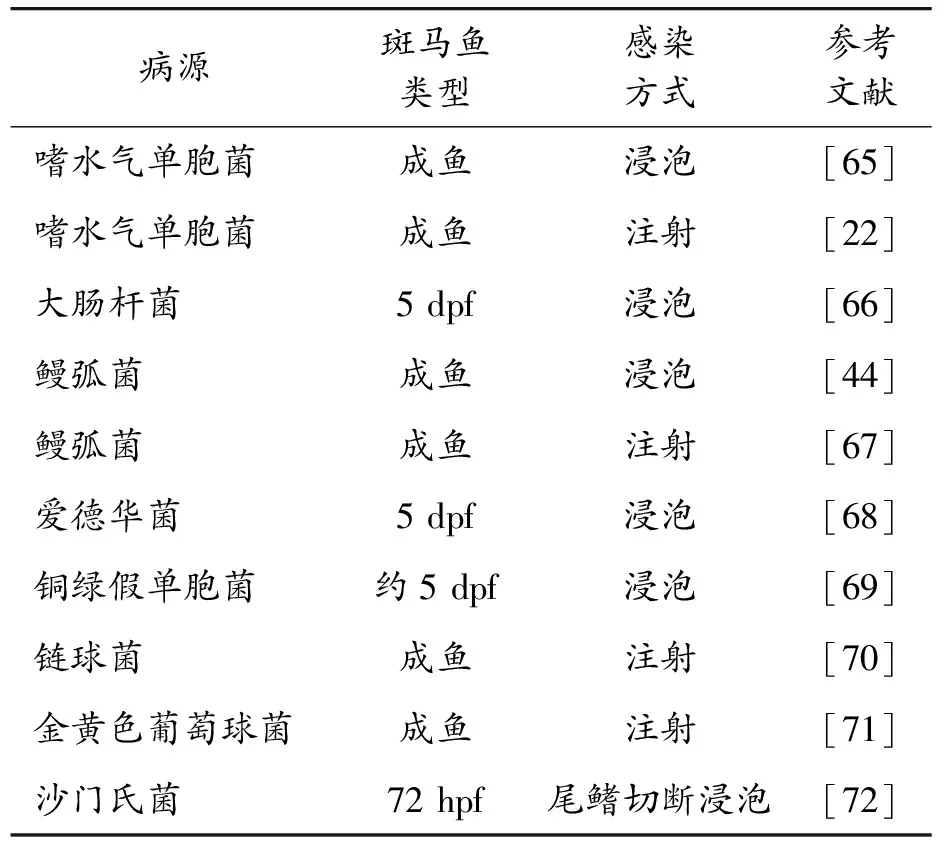
表2 斑馬魚免疫活性評價用病原菌及感染方式列表
2.4 研究涉及的信號通路
宿主固有免疫反應是抵御入侵微生物的第一道防線。它的功能是在感染的第一階段識別入侵的病原體并啟動適當的免疫反應。病原體相關分子模式(pathogen-associated molecular patterns,PAMPs)和損傷相關分子模式(damage associated molecular patterns,DAMPs)被模式識別受體(pattern recognition receptors,PRRs)識別,其中Toll樣受體(toll-like receptors,TLR)家族的研究最為廣泛。髓樣分化因子(myeloid differentiation factor 88,MyD88)是TLRs信號通路中的重要適配器蛋白,因為它被除TLR3以外的所有TLRs[73]所使用。其C端TIR結構域能夠與TLRs相互作用,N端死亡結構域能夠與白細胞介素-1受體相關激酶4(interleukin-1 receptorassociated kinase 4,IRAK4)相互作用,后者又招募白細胞介素-1受體相關激酶(interleukin-1 receptorassociated kinase 1,IRAK1)或白細胞介素-1 受體關聯激酶2 (interleukin-1 receptorassociated kinase 2,IRAK2)形成“Myddosome”信號傳導復合物,激活核因子κB(nuclear factor kappa-B,NF-κB)和絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號傳導[74]。Myddosome還在白細胞介素-1(interleukin-1,IL-1),IL-18和IL-33受體的下游起作用,并且它與IFN-γ受體信號傳導有關,增加了其在炎癥和宿主防御中的核心作用。TLRs是一類重要的PRRs,可識別微生物和危險信號。它們在配體結合時的下游信號對于固有免疫反應的啟動至關重要。在人類和哺乳動物模型中,MYD 88以其在IL-1受體(IL-1R)和TLRs信號傳導中作為適配器分子的核心作用而聞名。Van der Vaart M[75]描述了一個帶有截短的MyD 88的斑馬魚系,作為TLRs信號組件的第一個斑馬魚突變體,證明免疫受損的突變體在標準飼養條件下的存活率較低,并且更容易受到急性細菌病原體遲緩愛德華氏菌和鼠傷寒沙門氏菌的挑戰,基因微陣列分析和定量PCR分析顯示,固有免疫(包括NF-κB和AP-1)和促炎細胞因子IL-1β的轉錄因子的基因表達在這些細菌感染期間依賴于MyD 88信號傳導。
NOD樣受體(NOD-like receptor,NLR)是一大類細胞內PRRs,具有核苷酸結合域(nucleotide binding domain,NBD)和富含亮氨酸的重復(leucine rich repeat,LRR)區域的特征性排列。NLR最突出的功能是識別細胞內病原體相關分子模式(PAMPs)[76]。系統發育分析揭示了NLR家族中的3個不同的亞家族:NOD(NOD1-2,NOD3/NLRC3,NOD4/NLRC5,NOD5/NLRX1,CIITA),NLRP(NLRP1-14,也稱為NALPs)和由IPAF(NLRC4)和NAIP組成的IPAF亞家族[77]。其中,NOD1是第一個被表征的NLR成員[78]。在哺乳動物中,NOD1可感知D-谷氨酰-內消旋二氨基庚酸(DAP),這是在所有革蘭氏陰性細菌和少數革蘭氏陽性細菌中發現的細菌肽聚糖(peptidoglycan,PGN)片段[79]。配體結合后,NOD1通過CARD-CARD結構域相互作用與受體相互作用蛋白2(RICK/RIP2)相互作用,從而觸發NF-κB和MAPK途徑的激活[80]。這些信號級聯反應有效地上調促炎細胞因子和抗菌分子的產生,構成固有免疫反應。
CD44是一種跨膜黏附分子,是透明質酸的主要受體。除了介導T細胞外滲的關鍵作用[81],調節效應器T細胞反應[82]和T細胞發育,CD44還激活PI3K-Akt途徑,該途徑與包括白血病細胞在內的各種細胞類型的細胞存活有關[83]。有研究已經確定了CD44,TLRs和PI3K-Akt通路在病理條件下的相互作用[84-85]。然而,NOD1、CD44和PI3K-Akt途徑之間的功能相關性是否存在于免疫系統中,特別是在早期本體發生期間,目前尚不清楚。
PRRs對于宿主防御和組織穩態對抗感染病原體至關重要。PRRs在不同物種間具有高度保守性,表明它們在基本生物過程中的關鍵作用。盡管對NOD1受體在固有免疫和獲得性免疫反應的研究已經比較充分,但對NOD1在胚胎和幼魚階段的作用仍然知之甚少。Hu等[86]報道了NOD1對于PI3K-Akt途徑的調節和斑馬魚幼魚存活是必要的。轉錄組分析表明,NOD1-/-斑馬魚中主要在參與新陳代謝和免疫系統過程的通路中發生顯著富集。生化分析表明,CD44a的表達需要NOD1,而CD44a又在幼魚發育過程中激活PI3K-Akt途徑。相反,在NOD1缺陷斑馬魚中CD44a的過度表達恢復了PI3K-Akt途徑的調節并改善了幼魚的存活率。總的來說,Hu等的研究表明,NOD1通過CD44a介導的PI3K-Akt信號傳導的激活,在幼魚存活中起著以前未被發現的保護作用。
3 結論
概述了斑馬魚模型在多糖免疫中的應用和獨特優勢,介紹了采用斑馬魚模型進行多糖免疫相關研究的進展。免疫治療是近年來的研究熱點話題,尤其是在PD-1、PDL-1調節免疫系統抗癌方面更是不斷取得重大突破[87-88]。多糖的種類繁多,研究前景廣闊,在免疫方面具有明顯的促進免疫活性和抗菌作用;然而許多相關的機制尚不清楚,有待于進一步研究與開發。與傳統的經典體內、外模型相比,斑馬魚免疫活性篩選模型兼具體內和體外的優勢:既有體外的操作相對簡單、經濟、快速,適合高通量篩選的特點;又同時屬于整體動物模型,具備相對完整的代謝系統,幼魚整體透明便于觀察,所得到的數據比單純的體外篩選結果更加可信,在今后的藥物篩選研究中依舊有良好的前景。
