BYDV GAV編碼蛋白質的定位、互作及致病相關功能研究
2022-11-03 11:54:34韓曉玉李慶倫姜興林楊靈玲史亞娟李洪連陳琳琳
華北農學報 2022年5期
韓曉玉,李慶倫,姜興林,王 賀,楊靈玲,史亞娟,李洪連,陳琳琳,楊 雪,施 艷
(河南農業大學 植物保護學院,河南 鄭州 450002)
大麥黃矮病毒(Barleyyellowdwarfviruses,BYDVs)可以感染各種禾本科作物,例如小麥(Triticumaestivum)、大麥(Hordeumvulgare)和燕麥(Avenasativa)等[1-2],對禾本科作物生產造成嚴重的產量損失[3],是世界范圍內禾本科作物上的重要病毒[4-5]。BYDVs引起的小麥黃矮病是我國小麥生產上重要的病毒病[6],目前該病毒主要分布在西北、華北、東北、華中、西南及華東等冬麥區、春麥區及冬春麥混種區。小麥感病后植株矮化,葉尖出現倒“V”字形黃化,一般減產10%~20%,嚴重的可達到50%以上,個別地塊甚至可造成絕產。病毒感染會導致宿主植物顯示葉片變黃的癥狀,這種現象通常與葉綠素生物合成功能損害有關,導致光合作用減弱[7-9]。病毒編碼蛋白質在病毒侵染過程中發揮重要作用,因此,研究BYDVs編碼蛋白質的功能將為后續解析該病毒的致病機制奠定基礎。
BYDVs是一組由黃癥病毒屬、馬鈴薯卷葉病毒屬和未歸屬的種組成的正單鏈RNA病毒[3],通過蚜蟲進行持久非增殖型傳播,基于蚜蟲介體的特異性和病毒的序列,BYDVs可分為10個典型株系,分別為:黃癥病毒屬的BYDV PAV、PAS、MAV、kerⅡ和kerⅢ株系,馬鈴薯卷葉病毒屬的谷類黃矮病毒(Cerealyellowdwarfvirus,CYDV)RPV和RPS株系,以及玉米黃矮病毒(Maizeyellowdwarfvirus,MYDV)RMV株系和黃癥病毒科未歸屬的BYDV GPV和SGV株系[10-12]。在我國,目前報道的BYDVs株系有4個,分別為:BYDV GPV、GAV、PAV和RMV。其中,BYDV GAV與美國MAV有著相同的血清學反應,全基因組序列一致性最高,為90.4%[10],但是二者傳播介體不同,GAV可以由麥長管蚜和麥二叉蚜傳播,MAV僅能通過麥長管蚜傳播,因此,GAV是我國特有的一種BYDVs,已成為我國小麥黃矮病的主要病原[13-15]。
BYDVs基因組全長約5.7 kb,5′端無帽子結構,3′端無poly(A)尾,編碼7個閱讀框,分別為:ORF1和ORF2編碼與復制相關的蛋白質P1和P2;ORF3編碼外殼蛋白(CP);ORF4編碼移動蛋白(MP),有研究表明,MP還可以作為沉默抑制子發揮功能[16];ORF5編碼CP通讀蛋白,該蛋白質與病毒粒子的穩定性和蚜蟲傳播有關[17];ORF6編碼P6蛋白,其在黃癥病毒中變異較大,目前的研究認為,該蛋白質是BYDV GAV的沉默抑制子蛋白[18];ORF3a編碼P3a蛋白,其是由非AUG起始的一個40~50 個氨基酸的小蛋白質,研究表明,該蛋白質能夠影響馬鈴薯卷葉病毒屬蕪菁花葉病毒(Turnipmosaicvirus,TuMV)的系統移動[19]。
目前,BYDV GAV編碼蛋白質MP和P6已有相關研究[18,20],而有關P1、P2和CP的功能研究較為缺乏,3個病毒蛋白是否通過與其他病毒蛋白協作參與病毒侵染,是否參與病毒的致病過程尚不清楚。鑒于此,本研究對3個蛋白質的亞細胞定位、互作情況和致病相關功能進行研究,分析P1、P2和CP與BYDV GAV編碼蛋白質的互作情況和對病毒致病性的影響,旨在為研究該病毒的致病機制提供理論依據。
1 材料和方法
1.1 試驗材料
病毒序列來源:BYDV GAV病毒編碼蛋白序列來源于河南農業大學植物保護學院土傳病害實驗室保存的BYDVGAV病毒性克隆質粒。
供試植物:本氏煙(Nicotianabenthamiana)種植于本實驗室溫室。
菌株與載體:大腸桿菌DH5α感受態細胞及農桿菌GV3101感受態細胞購于莊盟生物公司,保存于-80 ℃冰箱。pEG104載體用于亞細胞定位的觀察,pGR106載體即馬鈴薯X病毒(PotatovirusX,PVX)載體,用于致病因子的測定,cYFP和nYFP載體用于雙分子熒光互補(Bimolecular fluorescent complimentary,BiFC)試驗。以上載體均保存在本實驗室。
1.2 BYDV GAV編碼P1、P2和CP蛋白的同源進化分析
在NCBI網站查找BYDVs不同株系BYDV GAV、GPV、PAV和RMV的基因組,將編碼蛋白質P1、P2和CP的核苷酸序列進行Blast比對,對同源性高的核苷酸序列采用鄰接法(Neighbor joining,NJ)進行系統進化分析。利用Mega 7.0軟件建立系統發育樹。
1.3 載體構建所用引物的設計
通過DNAMAN軟件設計引物,送由上海生工生物工程技術服務有限公司合成,引物序列見表1。

表1 引物序列Tab.1 Primer sequences
1.4 BYDV GAV編碼P1、P2和CP蛋白的亞細胞定位觀察
用Gateway方法利用BPBYDVP1F/BPBYDVP1R、BPBYDVP2F/BPBYDVP2R和BPBYDVCPF/BPBYDVCPR分別構建BP-P1、BP-P2和BP-CP中間載體。用中間載體分別和pEG104載體進行LR反應構建P1-YFP,P2-YFP和CP-YFP熒光表達載體,轉化農桿菌感受態GV3101,挑取單菌落驗證正確后搖菌。將含有P1-YFP、P2-YFP和CP-YFP質粒的農桿菌用MMA緩沖液(10 mmol/L MgCl2,10 mmol/L MES,200 μmol/L 乙酰丁香酮,pH值5.6)懸浮后,OD600值定為1,靜置2 h后浸潤健康本氏煙葉片(第7,8片葉)下表皮,接種2 d后在激光共聚焦顯微鏡(Zeiss 810)下觀察本氏煙葉片表皮細胞中YFP熒光,其激發光波長為514 nm。
1.5 BYDV GAV蛋白質互作的BiFC驗證
通過BiFC技術可以將具有相互作用親和力的2個蛋白質組裝成完整的熒光蛋白。利用PNCBYDVP1F/PNCBYDVP1R、PNCBYDVP2F/PNCBYDVP2R、PNCBYDVCPF/PNCBYDVCPR、PNCBYDVMPF/PNCBY
DVMPR和PNCBYDVP6F/PNCBYDVP6R等引物分別構建BYDV P1、P2、CP、MP和P6的nYFP和cYFP載體。將構建成功的BYDV P1、P2、CP、MP和P6的nYFP和cYFP載體轉化農桿菌感受態GV3101,得到的農桿菌用MMA緩沖液懸浮至OD600值為0.1。將不同編碼蛋白質的nYFP和cYFP菌液分別組合,浸潤3—4葉期本氏煙,接種2 d后用激光共聚焦顯微鏡觀察熒光并拍攝。
目前,廣西各項社會保險費的征繳主要采取社會保險經辦機構征收的模式,即由人力資源和社會保障部門社會保險經辦機構負責征收基本養老保險費(含城鎮職工基本養老保險、城鄉居民基本養老保險、機關事業單位養老保險)、基本醫療保險費(含城鎮職工基本醫療保險和城鄉居民基本醫療保險。自2017年7月1日起,新型農村合作醫療保險并入城鄉居民基本醫療保險)、失業保險、工傷保險、生育保險等社會保險費。
1.6 PVX介導的BYDV GAV蛋白質系統表達
使用引物PVXBYDVP1-F/PVXBYDVP1-R、PVXBYDVP2-F/PVXBYDVP2-R和PVXBYDVCP-F/PVXBYDVCP-R,利用同源重組的方法將BYDV P1、P2和CP構建到pGR106載體上。將構建成功的PVXP1、PVXP2、PVXCP表達載體進行農桿菌轉化,對利用菌液PCR驗證正確的單菌落進行搖菌。將PVXP1、PVXP2、PVXCP農桿菌分別單獨浸潤本氏煙,以PVX作為對照,OD600值定為0.5,浸潤后黑暗處理12 h。觀察植株是否出現危害癥狀,記錄天數并進行葉片拍照,取樣放入-80 ℃備用。
1.7 Western Blot檢測
接種PVXP1、PVXP2、PVXCP和PVX 5 d后,取植株系統葉。將取樣葉片進行蛋白質提取,以PVX載體作為對照,將蛋白質樣品進行SDS-PAGE凝膠電泳;利用20 V、20 min條件進行轉膜;用5%的脫脂奶粉對PVDF膜進行封閉;一抗是PVX-CP抗體,稀釋8 000倍,二抗是過氧化物酶標記的羊抗小鼠抗體,稀釋10 000倍;用底物化學發光劑eECL(諾唯贊生物試劑公司)進行顯色。
2 結果與分析
2.1 BYDV GAV編碼蛋白質P1、P2和CP的遺傳進化分析
通過Mega 7.0構建系統進化樹,對BYDVGAV株系P1、P2和CP蛋白的核苷酸序列進行同源進化分析,如圖1所示,根據P1的核苷酸序列,BYDV GAV的分離物聚到一小支,與BYDV PAV株系分離物在一個大的分支上,與BYDV GPV和RMV株系分離物親緣關系較遠;BYDV PAV中國分離物聚到一個小支,美國和澳大利亞分離物聚在一個小支。根據P2的核苷酸序列構建的系統進化樹與P1的較為相似,BYDV GAV與PAV株系分離物親緣關系近,與BYDV GPV和RMV株系分離物親緣關系較遠(圖2)。根據CP的核苷酸序列構建的系統進化樹表明,BYDV GPV運城分離物(KP096592.1)與其他BYDVs分離物親緣關系都較遠,單獨形成一支,GAV與PAV親緣關系最近,二者能夠與RMV分離物形成一個分支,而與GPV親緣關系遠(圖3)。綜上表明,BYDVs的4種分離物中,PAV與GAV在核苷酸水平上的親緣關系最近。

圖1 P1編碼序列的同源進化分析Fig.1 Homologous evolution analysis of the coding sequence of P1
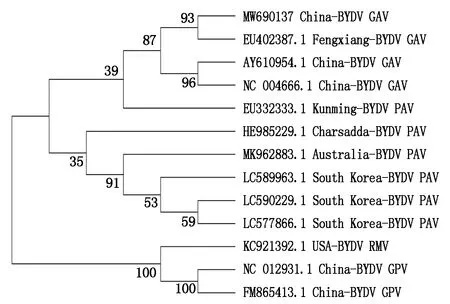
圖2 P2編碼序列的同源進化分析Fig.2 Homologous evolution analysis of the coding sequence of P2

圖3 CP編碼序列的同源進化分析Fig.3 Homologous evolution analysis of the coding sequence of CP
2.2 BYDV GAV編碼蛋白質 P1、P2和CP的亞細胞定位
分別構建P1-YFP、P2-YFP和CP-YFP重組表達載體,將含有P1-YFP、P2-YFP和CP-YFP質粒的農桿菌菌液分別浸潤本氏煙葉片,接種后2 d通過激光共聚焦顯微鏡觀察YFP熒光,結果表明,P1、P2和CP均定位在核質中(圖4)。

圖4 P1、P2和CP的亞細胞定位Fig.4 Subcellular localization of P1,P2 and CP
2.3 BYDV GAV編碼P1、P2和CP蛋白與其他蛋白質的體內互作驗證
分別擴增P1、P2、CP、MP和P6的全長閱讀框,構建到雙分子熒光互補載體nYFP和cYFP上,通過兩兩組合共15個組合浸潤本氏煙,接種后2 d通過激光共聚焦顯微鏡觀察本氏煙細胞,結果表明,P1-nYFP和P1-cYFP共浸潤后可以在細胞四周觀察到點狀YFP熒光,表明P1能夠在體內互作(圖5),其他組合均未觀察到熒光(表2)。

圖5 P1在體內相互作用的驗證Fig.5 Verification of the self-interaction of P1 in vivo

表2 BYDV GAV的P1、P2和CP蛋白與其他蛋白質相互作用觀察結果Tab.2 The observation results of the interaction between P1,P2,CP and other proteins encoded by BYDV GAV
2.4 BYDV GAV編碼蛋白質P1、P2和CP異源表達對PVX致病性的影響
分別構建P1、P2和CP的PVX重組表達載體,轉化農桿菌后浸潤本氏煙,接種5 d后異源表達CP的PVX載體處理可以明顯地觀察到系統花葉癥狀,而PVX對照和PVXP1、PVXP2處理都尚未產生系統癥狀,表明CP促進了PVX的癥狀形成(圖6-A)。采用PVX-CP的抗血清通過Western Blot檢測PVX的積累量,結果與癥狀一致,PVXCP處理可以明顯觀察到PVX的積累,而其他重組載體處理未能檢測到病毒的積累(圖6-B)。為了確定PVX、PVXP1和PVXP2的侵染性,采集接種后10 d的系統葉進行Western Blot檢測,結果表明,接種后10 d,PVX、PVXP1、PVXP2和PVXCP處理都可以檢測到CP的積累。
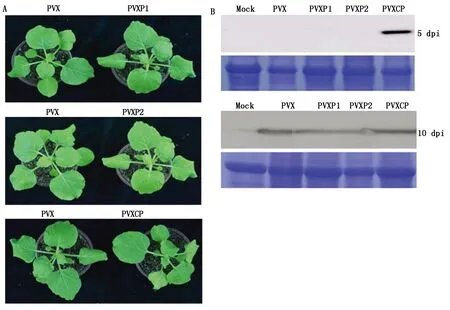
A.接種本氏煙后5 d的植株癥狀;B.分別在接種后5,10 d檢測病毒積累量。A.The symptom of plants at 5 d post infiltration(dpi);B.Detection of viral accumulation at 5,10 dpi,respectively.
3 結論與討論
BYDV GAV是我國小麥黃矮病的主要病原[14],P1、P2和CP是該病毒編碼的3個蛋白質,目前這3個蛋白質在BYDV GAV侵染中的作用鮮有報道。本研究通過瞬時表達觀察三者的亞細胞定位發現,P1和P2蛋白定位于核質,這與PSORT Ⅱ在線預測(https://psort.hgc.jp/form2.html)的結果一致;CP-YFP融合蛋白定位于核質,而PSORTⅡ和LOCALIZER(http://localizer.csiro.au/)在線預測CP定位于線粒體和細胞核。
研究病毒蛋白質間的相互作用發現,P1能夠在體內自身互作,而其他蛋白質之間體內互作強度可能較弱,因而沒有檢測到。本研究中,P1和P2的表達對PVX的侵染沒有明顯影響,推測P1和P2可能參與了病毒侵染的其他過程。已有研究表明,馬鈴薯卷葉病毒(Potatoleaf-rollvirus,PLRV)編碼的P1和P2參與了病毒的復制[21],病毒蛋白質之間的互作能夠有助于病毒的復制和移動。本研究中,通過體內互作的方式確定了P1的自身互作,表明P1可能通過形成點狀聚集體的形式參與復制。
黃癥病毒屬甜菜西黃病毒(Beetwesternyellowsvirus,BWYV)的CP突變體能夠抑制該病毒的系統侵染[22],本研究發現,BYDV GAV CP蛋白的表達可以顯著促進PVX的系統侵染和癥狀形成,與黃癥病毒屬BWYV的結果一致。此外,研究發現,抗小麥黃矮病相關蛋白激酶TiDPK1與BYDV PAV和GAV株系的CP蛋白能夠互作[23],表明CP可能參與了相關病毒的致病過程。
BYDV-GAV編碼的蛋白質P1、P2和CP均通過定位于核質中發揮作用,P1在體內能夠以點狀二聚體的狀態參與病毒的侵染,CP異源表達能夠顯著促進PVX的系統性侵染。
