木林子國家級自然保護區珍稀植物伯樂樹天然種群結構與動態研究
2022-11-08 05:45:38薛衛星劉松柏艾訓儒王志鳴
湖北林業科技 2022年5期
關鍵詞:結構
胡 澳 艾 鑫 薛衛星 朱 旻 劉松柏 艾訓儒 王志鳴
(1.湖北民族大學林學園藝學院 恩施 445000;2.湖北木林子國家級自然保護區管理局 恩施 445000;3.五峰土家族自治縣林業局 宜昌 443499)
種群結構與種群動態的研究是探究植物種群的重要方向,兩者都是基礎生態學的主要內容[1,2]。在對種群結構和種群數量及動態分析時,通常會用到靜態生命表、存活曲線、動態量化、生存分析等方法[3]。通過靜態生命表、種群存活曲線的分析,進而引入生存函數后進行探究,既可分析種群的生存現狀,也能展示種群對外界因子的適應性[4-6]。種群動態量化分析可以較好地預測種群的動態變化[1,7]。種群結構及其動態研究不但能夠反映整個種群的生存現狀,還能夠反映種群和環境之間的相互作用關系及種群在整個群落中的地位與功能,對種群資源保護和開發與利用具有重要意義[8-10]。
伯樂樹Bretschneiderasinensis又名鐘萼木,為疊珠樹科伯樂樹屬落葉喬木。樹高通常可達20多米,樹干體呈通直挺拔的姿態,被平滑的褐色樹皮包裹,塔形的樹冠高懸林中。伯樂樹是中性偏陽樹木,常散生在較濕潤的溝谷坡地以及溪邊的常綠落葉闊葉混合林中,其主根直、側根比較發達,深根性樹木的特點使伯樂樹抗風能力較強,也稍能抗寒,但卻不耐熱[11]。作為笫三紀古熱帶植物區系的孑遺植物,伯樂樹是研究被子植株的種系發生學和古地理學、古氣候學等的首選材料。但長期以來,由于原生境的自然破環與人為擾動導致伯樂樹野外種群數量銳減,再加上自然結實率較低、自然更新困難等原因,使伯樂樹陷入了瀕危狀況[12],在2021年7月8日公布的《國家重點保護野生植物名錄》中被列為國家II級保護植物。近年來,人們對伯樂樹的研究主要集中在苗木栽培[13]、遺傳特征[14]以及生態習性[15]等方面,對種群結構與動態、資源量估算等方面的研究相對較少。本研究以木林子國家級自然保護區內野生的伯樂樹種群為研究對象,探究野生伯樂樹種群的種群結構與動態特征、資源分布特征,探討野生伯樂樹種群生存現狀及其發展趨勢,旨在為野生伯樂樹種群的保護和利用提供理論依據。
1 研究區概況
木林子國家級自然保護區(以下簡稱“木林子保護區”)地處武陵山余脈,位于湖北省恩施土家族苗族自治州鶴峰縣以北,東經109°59′30″~110°17′58″,北緯29°55′59″~30°10′47″,平均海拔為1 100.0~2 095.6 m,主峰牛池峰屬于湖北長江以南的第一高峰。研究區氣候環境屬于中亞熱帶濕潤季風氣候,年平均氣溫15.5 ℃,絕對高溫達39 ℃,絕對低溫至-17.1℃,年平均降水量1 700~1 900 mm,年平均相對濕度為82%,無霜期270~279 d。自然土壤從低至高依次為山地黃壤、山地黃棕壤、山地棕壤[16]。保護區內植物物種資源豐富,據初步調查統計有維管束植物203科918屬2 689種,域內有珙桐Davidiainvolucrata、水青樹Tetracentronsinense、伯樂樹、香果樹Emmenopteryshenryi、連香樹Cercidiphyllumjaponicum等多種珍稀瀕危植物,其中伯樂樹為保護區典型的珍稀瀕危物種之一,分布于保護區海拔1 000~1 500 m的溝谷中。
2 研究方法
2.1 樣地設置與調查方法
在伯樂樹可能分布的區域(鶴峰縣中營鎮三家臺蒙古族村、下坪鄉小園村麻油榨、木林子大東州黑灣及場灣核心區),按照可通行性、群落代表性原則設計調查線路。沿調查線路用望遠鏡尋找兩側伯樂樹種群個體,在發現有伯樂樹種群個體分布的地點,調查并記錄伯樂樹胸徑、樹高、東西冠幅、南北冠幅、枝下高、生長狀態、萌蘗情況等立木因子,同時以目的物種為圓心調查直徑10 m范圍內的植被情況。
2.2 種群年齡結構劃分
了解伯樂樹種群的年齡結構是研究伯樂樹種群動態發展的基本方法。根據從木林子保護區調查到的伯樂樹種群單株數據,以5 cm為徑級差值,將伯樂樹天然種群劃分為8個徑級,根據伯樂樹天然種群的樹高特征,以5 m為高度差,將種群劃分為7個高度級(表1)。
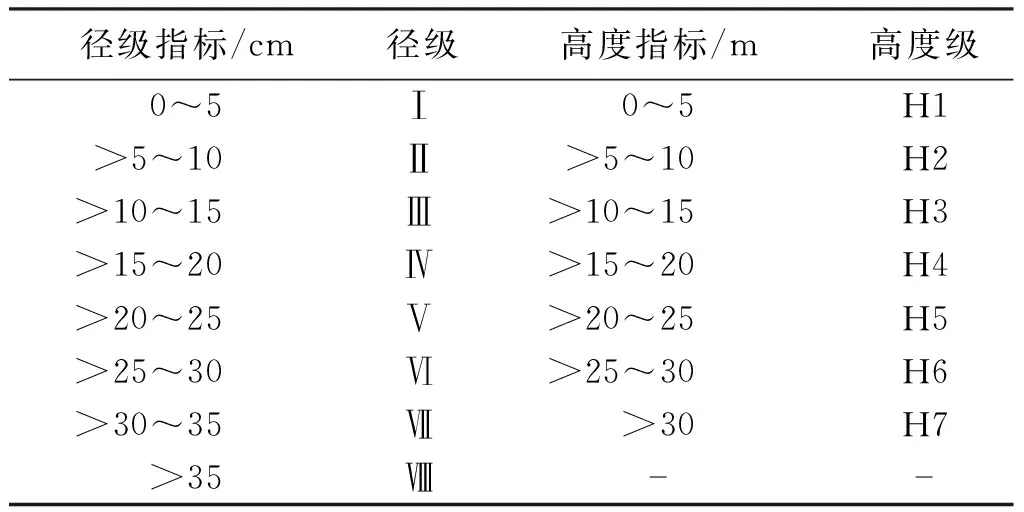
表1 伯樂樹種群年齡結構劃分
2.3 動態量化方法
采用生態學中以徑級代替齡級方法分析種群結構,根據徑級結構將伯樂樹種群年齡劃分為8個齡級。對種群結構采用動態量化分析方法,參考陳曉德[17]種群與群落結構動態量化分析法對伯樂樹種群年齡結構動態進行定量描述。
2.4 靜態生命表與存活率曲線、死亡率曲線
靜態生命表又叫作特定時間生命表[18],是描述種群死亡過程的具有固定格式的表,通常記述一個世代全部死亡的整個過程的生存和生殖情況,從而分析影響種群數量的各個因素。編制靜態生命表并對其進行分析,能夠較為清楚地展示出種群的結構現狀,對種群生命史、未來發展以及保護的研究起到重要意義。但由于靜態生命表所反映的年齡動態是種群多個時代重疊的結合,并不是對這個種群全部生活史跟蹤[19],因此在實際研究中經常會出現負數死亡率的情況,對此,本研究采取勻滑技術[20]的方法對各個齡級的個體數進行勻滑調整,對野生伯樂樹種群進行靜態生命表的編制。
2.5 伯樂樹種群生存分析
本研究對伯樂樹種群結構的生存分析參考楊鳳翔[21]的生存分析理論分為4個生存函數,分別為:生存率函數(Si)、累積死亡率函數(Fi)、死亡密度函數(i)和危險率函數(λi)。
3 結果與分析
3.1 種群結構
3.1.1 齡級結構
種群齡級結構能清晰地表現種群的生活狀況[22]。本研究于木林子保護區共調查野生伯樂樹個體108株,其中最大胸徑為45.0 cm;根據徑級結構分析(表2),可知在研究區域內的伯樂樹種群中0~5 cm胸徑株數占比最多;采用徑級代替齡級的方法對伯樂樹種群的齡級結構進行分析,由圖1可知,其中低齡級(Ⅰ~Ⅲ齡級)個體數的總和占總株數的76.85%,分別為:Ⅰ齡級個體43株,占比39.81%;Ⅱ齡級個體25株,占比23.15%;Ⅲ齡級個體15株,占比13.89%。隨著伯樂樹種群年齡的增長,Ⅰ-Ⅳ齡級的個體數逐漸減少,Ⅴ齡級之后個體數量穩定在每個齡級4株,表明隨著野生伯樂樹年齡的增長,大徑級個體表現出對環境的逐漸適應性。但在總體上可以看出研究區域內野生伯樂樹種群徑級結構呈反“J”型分布,表現為增長型種族。
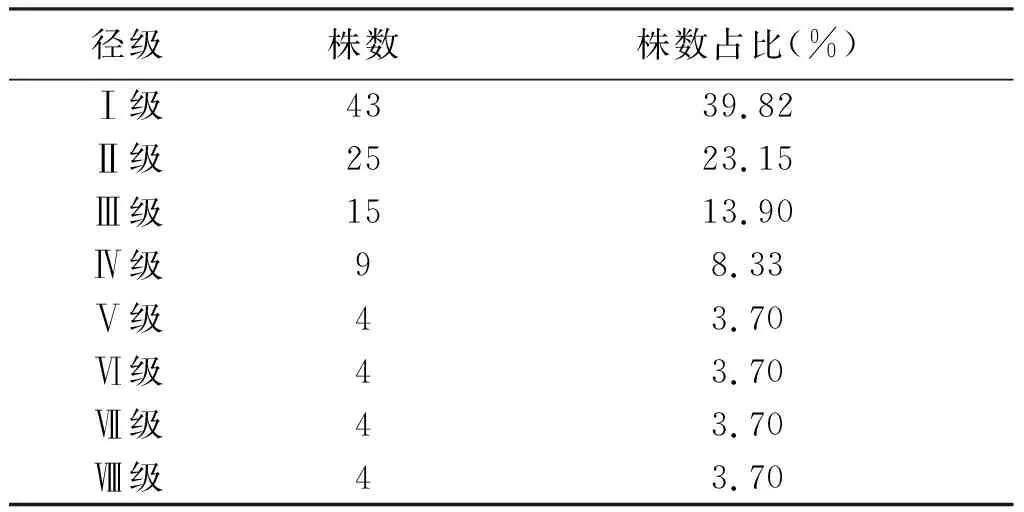
表2 伯樂樹天然種群徑級結構
3.1.2 樹高結構
種群樹高結構能夠較為清晰地顯示不同高度的個體在群落空間中所處的地位[23],是體現種群的生存與發展狀態的重要指標[24]。通過調查到伯樂樹種群的數據對其樹高進行劃分,共分為7個高度級(表3)。由表3可知,伯樂樹種群高度級在H1~H2個體數上升,H2之后開始遞減到H6降低到最小值1株,整體呈先上升后下降最后穩定的趨勢。H1~H3的個體較多,占了總株樹的84.26%,H2級株樹最多,占總株樹的37.03%,在H4級之后個體株樹只有17株,只占了總株樹的15.75%。綜合以上分析結果,可知伯樂樹種群的高度多分布于15 m以下(H1~H3),野生伯樂樹種群在自然群落中并不處于優勢地位,這也是該種群天然分布范圍逐漸縮小的原因之一。如果將H1~H2級看作一個整體,可認為伯樂樹種群的樹高結構呈現反“J”型分布。
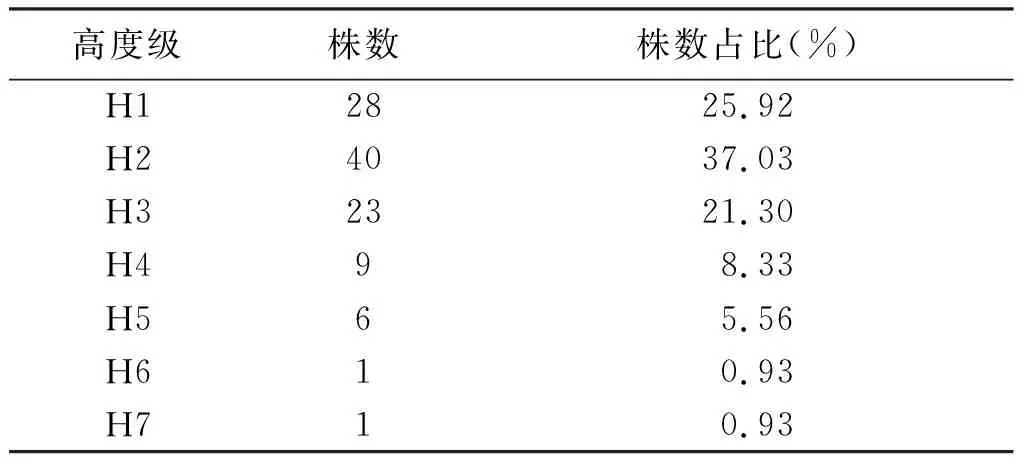
表3 伯樂樹天然種群樹高結構
3.2 種群動態量化分析
根據種群動態量化分析方法對伯樂樹種群進行動態量化分析,分析結果如表4所示。由表4可知伯樂樹種群動態指數V1、V2、V3、V4均大于0,說明Ⅰ~Ⅱ、Ⅱ~Ⅲ、Ⅲ~Ⅳ齡級之間呈現增長的結構動態,V5、V6、V7、V8均等于0,說明Ⅴ~Ⅵ、Ⅵ~Ⅶ、Ⅶ~Ⅷ齡級之間呈現穩定的結構動態。Vpi=37.50%>0說明種群整體為增長的趨勢,V'pi=1.17%接近于0,兩者的關系為Vpi>V'pi>0,隨機干預風險最大值Pmax=3.13%。綜上所述,木林子保護區伯樂樹種群呈現增長的趨勢,且增長勢較大,對外界環境干擾的敏感性較高。
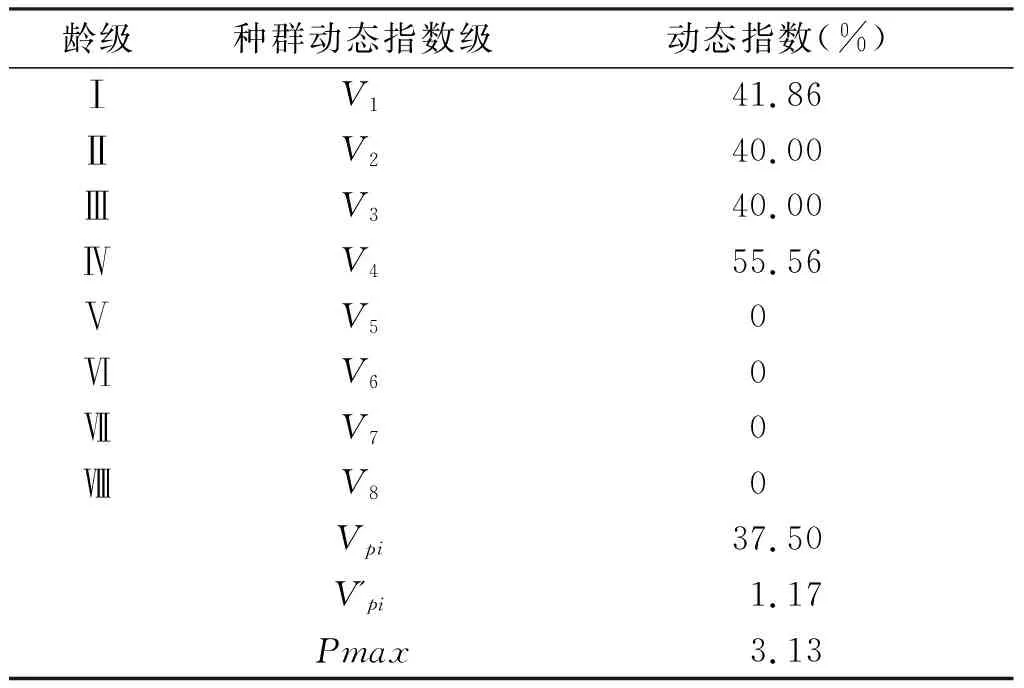
表4 伯樂樹天然種群動態變化指數
3.3 靜態生命表與存活曲線分析
3.3.1 靜態生命表
生命期望值(ex)能夠表現出從x齡級開始,每個齡級的種群平均期望生存能力[25]。如表5所示,伯樂樹種群的個體存活數量在每個齡級都比較大,隨著年齡的增加表現出下降的趨勢。在Ⅰ~Ⅴ齡級時,標準化存活數減少的較快,說明伯樂樹種群的幼苗、幼樹抗外界干擾能力較弱,生存率下降到了0.093%。從第Ⅴ齡級之后,標準化存活數達到穩定,此時不再有個體死亡,說明第Ⅴ齡級之后的伯樂樹個體生存能力已經可以適應外界環境因子干擾。生命期望值在第Ⅴ齡級時達到峰值,說明此時的伯樂樹種群個體的林木質量最佳,但從第Ⅴ齡級生命期望開始降低,說明伯樂樹種群在此之后會開始逐漸衰老。
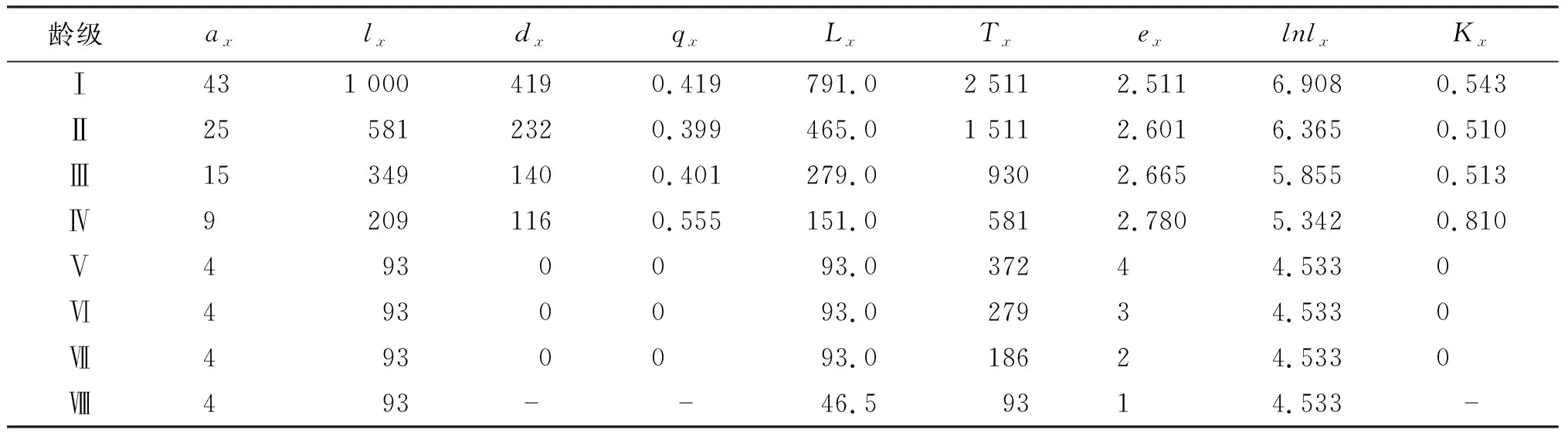
表5 伯樂樹天然種群靜態生命表
3.3.2 存活曲線分析
存活曲線是調查植物種群數量動態變化,以及進行種群統計研究的主要工具[26]。本研究以伯樂樹群體的標準化存活數對數為縱坐標,以齡級為橫坐標來描繪伯樂樹種群的標準化存活曲線,結果如圖1所示。對存活曲線采用Deevey-II和Deevey-Ⅲ數學模型建立模型,結果分別為:
Inlx=7.083 9e-0.066x(R2=0.878 4);
Inlx=7.194 5x-0.237(R2=0.923 5)。
對比建立的兩個數學模型R2值,發現冪函數模型R2值明顯高于指數模型,說明Deevey-Ⅲ型的模型結構與伯樂樹種群的存活曲線模型結構更加接近。由此可見,伯樂樹種群的幼苗、幼樹死亡率更高,這與種群結構和靜態生命表的分析結果一致。
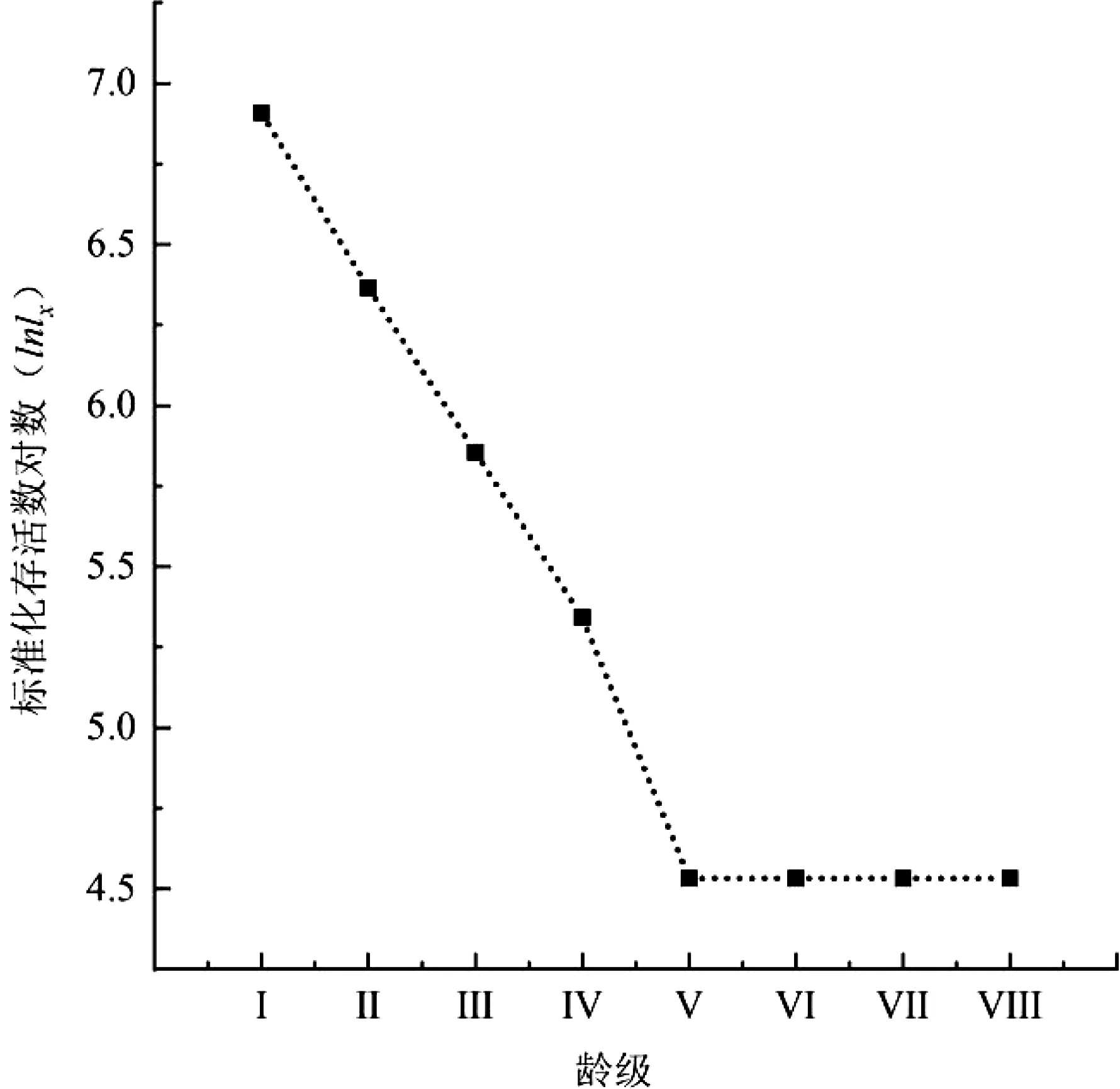
圖1 木林子伯樂樹種群存活曲線
3.3.3 死亡率與消失率曲線分析
種群數量動態變化隨著齡級增長需要通過死亡率和消失率曲線來表現[1]。木林子保護區伯樂樹種群死亡率與消失率曲線如圖2所示,伯樂樹天然種群死亡率與消失率曲線變化一致,整體表現為隨著年齡的增大,曲線趨勢先上升后遞減為0,結合靜態生命表可以看出伯樂樹種群的幼苗儲備量較大,在Ⅰ~Ⅳ齡級時苗木的死亡個體數較大,且在第Ⅳ齡級時達到死亡個體數的峰值。在Ⅰ齡級階段死亡率達到了41.90%,Ⅰ~Ⅳ齡級均擁有高于30%的死亡率,因此能夠達到下一個生長階段的苗木存活數只有93株,說明在苗木生長中期只有少數個體在面對外界環境因子干擾時能進入到下一個生長階段。
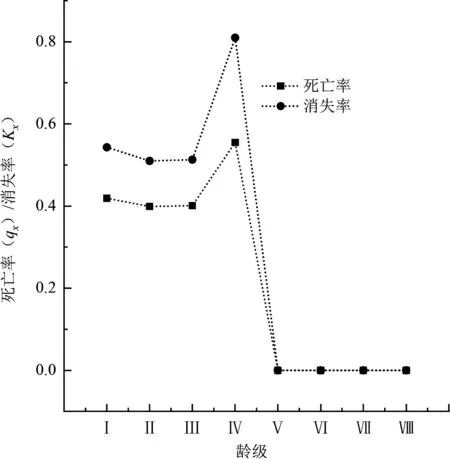
圖2 木林子伯樂樹天然種群死亡率與消失率曲線
3.4 生存分析
伯樂樹種群生存函數估計值見表6,分析可知伯樂樹種群在第V齡級之后穩定。木林子保護區伯樂樹種群以齡級為橫坐標,四種生存函數為縱坐標,分別對伯樂樹種群的生存率函數(Si)、累計死亡率函數(Fi)、死亡密度函數(fi)、危險率函數(λi)進行繪制曲線圖(圖3、圖4)。隨著種群年齡的增大,種群生存率(Si)呈現先下降后穩定的趨勢,累計死亡率(Fi)呈現先上升后穩定趨勢。在Ⅰ~Ⅳ齡級時生存率為下降的趨勢,且在Ⅰ~Ⅱ齡級下降最快,下降到了34.90%。累計死亡率與生存率呈互補,在Ⅰ~Ⅱ齡級之間相交,后累計死亡率開始大于生存率峰值達到了90.70%,生存率只有9.30%,之后變化穩定。死亡密度函數在Ⅰ~Ⅴ齡級緩慢下降,在Ⅰ齡級達到峰值8.30%,在Ⅴ齡級穩定下來到0,危險率函數在Ⅰ~Ⅲ齡級緩慢下降在到達Ⅲ齡級開始快速上升到第Ⅳ齡級之后急速下降至0,說明伯樂樹天然種群在第Ⅳ齡級時種群的發展遇到了較大的危機。根據以上4個函數的分析,表明了伯樂樹種群具有前期減少,中期遇到發展危機,后期穩定的特征。
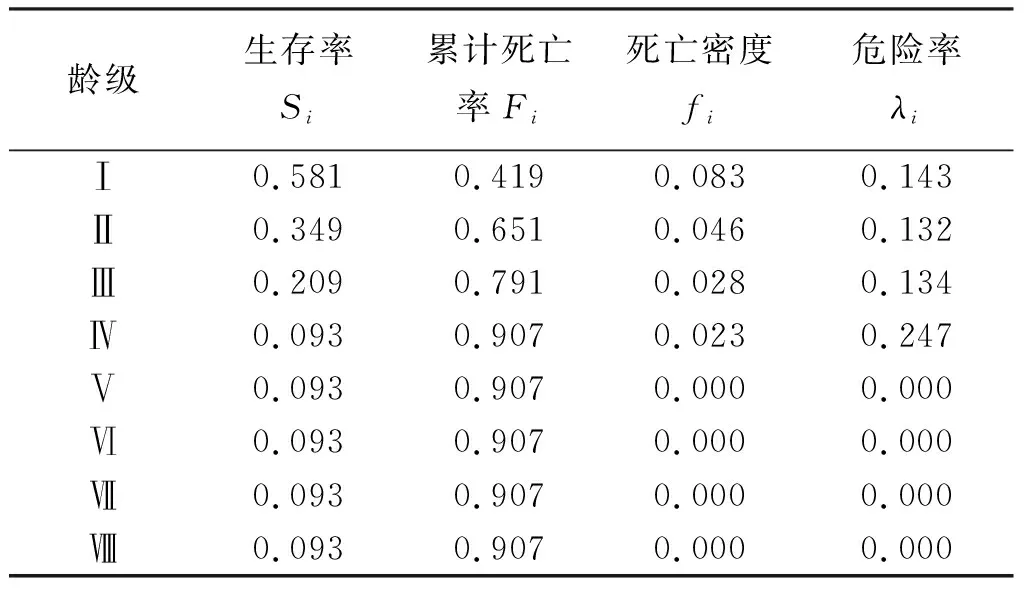
表6 伯樂樹天然種群生存函數估計值
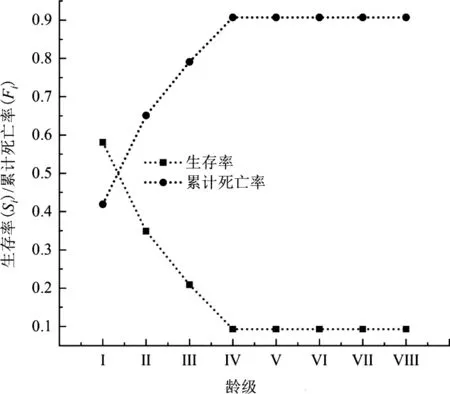
圖3 伯樂樹種群生存率和累計死亡率曲線
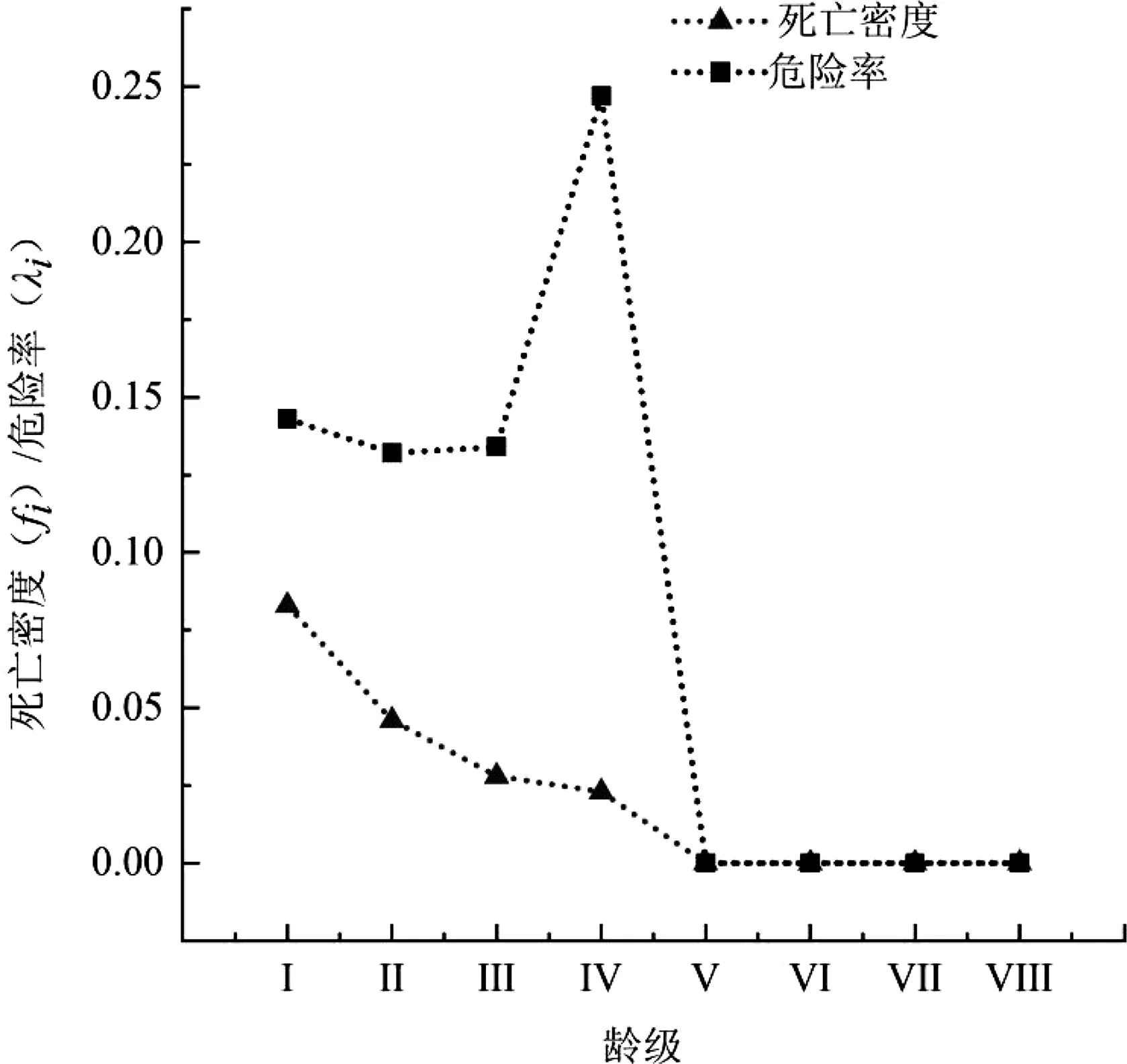
圖4 伯樂樹種群死亡密度和危險率曲線
4 結論與討論
4.1 伯樂樹種群結構
種群結構特征分析是揭示種群現狀和更新能力的重要途徑之一[27]。本研究通過對伯樂樹種群的徑級、樹高結構以及齡級結構進行分析得出,伯樂樹天然種群徑級結構、樹高結構總體呈現反“J”型分布,小徑級的個體數量豐富,屬于典型的增長型種群。幼樹階段的林木個體在天然更新的過程中對外界環境干擾表現出較強的敏感性,而種群的生長、更新往往伴隨著與其他物種在資源與空間上的競爭[23];由此可見,研究區域內的伯樂樹種群演替前期在群落中并不占優勢,而隨著齡級增加,個體數逐漸開始減少,少部分個體進入高齡級階段,且隨著同種個體的減少與種群在群落中逐漸處于林木上層,種內競爭減少、種間競爭逐漸占據相對優勢,野生伯樂樹種群在高齡級處于相對穩定狀態。而幼樹個體具備一定的數量優勢,從而保證了伯樂樹天然種群的自然更新能在特定環境種順利進行。綜上所述,經過環境的篩選和自然競爭存活下來的個體具有了較強的生存力[28-29],高齡級伯樂樹種群逐漸適應環境,林木個體數穩定。
4.2 伯樂樹種群生存現狀與動態特征
靜態生命表與存活曲線最能反映種群的生活狀況,以及抗外來環境影響因子影響的適應機制[29]。通過對木林子保護區伯樂樹天然種群生命表的分析可知,木林子保護區伯樂樹種群的小齡級個體死亡率比較高,生命期望值(ex)呈現先上升后減小的趨勢,消失率曲線(Kx)與死亡率曲線(qx)的趨勢隨著種群年齡增大先上升后降低最后達到穩定,存活曲線與數學模型Deevey-Ⅲ型相接近。在趙陽、齊瑞等[30]對尕海-側盆地區紫果云杉Piceapurpurea的研究中發現紫果云杉存活曲線呈Deevey-Ⅲ型;伯樂樹種群與紫果云杉種群的動態特征相似,且在研究區中發現伯樂樹種群伴生喬木多為杉木,而杉木屬于常綠針葉樹種,林下郁閉度較高,所以,杉木對伯樂樹種群的生長可能起到抑制的作用;由此可見,影響伯樂樹種群存活狀態的主要因素可能為環境因子[31]。
從靜態生命表中可以看出伯樂樹種群的幼苗個體數量較多,在幼樹階段時伯樂樹種群個體數快速下降,幼苗在成長為幼樹階段時,要與群落中的喬木層爭奪水分與陽光,而伯樂樹又為中性偏陽樹種,說明伯樂樹在研究區的自然環境中面對種群間的競爭中不占優,加上人為因子以及環境因子,導致伯樂樹種群由幼樹階段成長為小樹階段的個體數較少。而在達到小樹階段的伯樂樹種群個體生命期望達到峰值,說明伯樂樹種群在小樹階段能夠適應該研究區的環境且達到最高林木質量,此后的種群生命期望值降低,推斷是因為伯樂樹種群年齡正在逐步走向衰老。伯樂樹種群幼苗階段耐蔭性極強,但結實率不高,因此導致林下更新困難,而幼樹到小樹階段死亡率較高。因此,如何提高林下更新以及較多幼苗進入幼樹階段是當今研究伯樂樹種群資源的首要問題。
生存函數分析比起存活曲線能更直觀、更具體的表現種群結構的動態特征[23]。通過對伯樂樹種群的生存函數分析可以看出伯樂樹種群具有前期減少,中期遇到發展危機,后期穩定的特征,這和伯樂樹種群的靜態生命表、生存率與死亡率函數分析結果一致。由此可見,危險率函數更符合伯樂樹種群的生理和動態特征。植物種群動態是個體生存能力和外界環境的相互作用的結果[32]。本研究中伯樂樹天然種群Vpi=37.50%為正值,說明伯樂樹種群為典型的增長型種群,這與種群結構分析結果一致;而V'pi=1.17% 根據對木林子保護區伯樂樹種群的調查研究發現,伯樂樹天然種群幼苗個體數充足,種群結構表現為增長型,保證了未來一段時間的種群更新。研究區發現伴生喬木多為杉木,并且有人為破壞的痕跡,導致伯樂樹種群在幼樹階段因為人為破壞和群落個體競爭中得不到良好的發育條件。加上人類活動的干擾對生態環境的影響極大,幼苗對外界環境的干擾極為敏感,又因為結實稀少,幼苗無法長出發達的根系,導致種子萌發受限,幼苗更新困難。 綜上所述,可以推斷研究區內伯樂樹種群發展受限是因為人類對環境的破壞以及幼苗、幼樹階段生長環境的限制。因此,對伯樂樹種群的保護重心應該放在種群幼苗培育以及和生長環境保護上,對研究區的伯樂樹種群進行嚴加管護,幼樹個體進行適當的遷地保護,老齡個體采取間伐,伐掉瀕臨死亡的個體以及影響種群發展的個體。開展大眾知識普及,提高全民的保護意識,嚴厲打擊非法采伐等人為破壞環境的行為,保證研究區的伯樂樹種群不因人為破壞環境而減少。采取人工育苗的方式對伯樂樹進行繁殖,建立伯樂樹種子園來保護種群的生存與發展。5 結語
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
