快骨骼肌亞型肌鈣蛋白Ⅰ對雞前脂肪細胞增殖的影響
2022-11-08 07:10:16孔令喆李金煒張新宇旦增晉美孫澤孫嬰寧
高師理科學刊 2022年10期
孔令喆,李金煒,張新宇,旦增晉美,孫澤,孫嬰寧
快骨骼肌亞型肌鈣蛋白Ⅰ對雞前脂肪細胞增殖的影響
孔令喆,李金煒,張新宇,旦增晉美,孫澤,孫嬰寧
(齊齊哈爾大學 生命科學與農林學院,黑龍江 齊齊哈爾 161006)
實驗室前期研究發現,KLF7促進雞前脂肪細胞增殖,且調控基因的表達,但是TNNI2在雞前脂肪細胞增殖過程中的作用還不清楚.為此,探討基因對雞前脂肪細胞增殖的影響.qRT-PCR檢測永生化雞前脂肪細胞系(immortalized chicken preadipocyte cell line,ICP1)增殖過程中基因的表達情況;將TNNI2過表達載體pCMV-HA-轉染至ICP1細胞,利用Western blot實驗檢測過表達效果.采用CCK-8,EdU法檢測過表達TNNI2對細胞增殖的影響,qRT-PCR檢測增殖標志基因的表達.基因在ICP1細胞增殖過程中呈先下降后逐漸升高的趨勢;Western blot實驗結果表明,基因過表達成功;CCK-8結果表明,過表達TNNI2后ICP1細胞在450 nm處的吸光值顯著上升(<0.05);EdU結果顯示,過表達TNNI2后實驗組EdU摻入的細胞數占細胞總數的百分比顯著高于對照組(<0.01);qRT-PCR結果顯示,增殖標志基因表達水平有上調趨勢,但是差異并不顯著(>0.05).基因對雞前脂肪細胞的增殖具有促進作用.
雞;前脂肪細胞;過表達;TNNI2;增殖
體脂率過高是肉雞生長發育過程中的一個關鍵問題.過度的脂肪堆積會降低種雞的產蛋率,雞蛋的孵化率和受精率[1],也會影響飼料的轉化率,因此減少雞脂肪的過度沉積是肉雞行業急需解決的問題[2].脂肪沉積是一個受多因素影響的過程,從細胞水平上看,脂肪細胞數目變多和體積增大都會引起脂肪沉積[3].
快骨骼肌亞型肌鈣蛋白I(fast skeletal muscle troponin I,TNNI2)的編碼基因是脊椎動物肌鈣蛋白I(Troponin I,TnI)基因的一種亞型[4].作為決定動物肉質特征的潛在候選基因,的表達會影響肌肉纖維的組成,從而影響肉質的特征[5].最初的研究認為只在肌肉中表達并調節肌肉收縮[6],但隨著研究的深入,TNNI2更多的功能被發現.當在軟骨組織中表達時,TNNI2蛋白會抑制軟骨組織的發育[7],在乳腺癌細胞中,該基因作為雌激素受體相關受體的活化物來發揮功能,TNNI2在胰腺癌細胞中可以改變SIRT1的表達從而促進胰腺癌的發展[8].另外,研究發現遺傳性骨病的病因包括基因的突變[9],說明TNNI2在哺乳動物的骨骼發育過程中也發揮重要作用.在小鼠體內基因突變會阻礙小鼠的骨骼發育[10],基因的缺失或錯譯是導致遠端關節病的重要原因[11]等.近期研究發現,在豬、牛和雞的脂肪組織中表達,表明該基因在脂肪發育過程中也發揮作用[12].實驗室前期研究發現,TNNI2抑制雞前脂肪細胞分化,但是TNNI2在雞前脂肪細胞增殖中的作用還不清楚.因為天生胰島素抵抗,雞是研究脂肪組織發育的理想模型[13-15].本文以永生化雞前脂肪細胞系(immortalized chicken preadipocyte cell line,ICP1)為實驗材料,探究TNNI2對ICP1細胞增殖的影響.研究結果為深入研究肉雞脂肪過度沉積提供了依據,同時也為揭示肥胖發生的分子機制提供參考.
1 材料與方法
1.1 材料
ICP1細胞系(東北農業大學農業農村部雞遺傳育種重點實驗室);pCMV-HA質粒(美國Clontech公司);pCMV-HA-[16](齊齊哈爾大學基因功能研究實驗室).
1.2 方法
ICP1細胞系使用添加10%胎牛血清和1%青霉素-鏈霉素溶液的DMEM/12培養基在37 ℃ 5% CO2培養箱中培養.
1.2.1細胞轉染使用Lipo8000轉染試劑(Beyotime)進行轉染,質粒(μg)∶Lipo8000轉染試劑(μL)=1∶1.6.以6孔板為例,將細胞接種至6孔板,待細胞匯合度至70%~75%時進行轉染.125 μL無血清DMEM細胞培養基稀釋2.5 μg質粒,用移液器輕輕混勻,再加入4 μL Lipo8000轉染試劑,用移液器輕輕混勻后,立即均勻滴加至6孔板中,搖勻后置于37 ℃ 5% CO2細胞培養箱中繼續培養.轉染4~6 h后更換1次細胞培養基.于轉染后24,48,72 h分別收集細胞,用于提取細胞總RNA.
1.2.2CCK-8檢測細胞活力將細胞接種至96孔板并培養至完全貼壁,轉染pCMV-HA或pCMV-HA-載體,培養6 h后更換新鮮的培養基,放到37℃ 5%CO2培養箱中繼續培養24,48,72h后,向孔中加入10%培養基體積的CCK-8溶液,放入培養箱繼續孵育,2h后酶標儀測定450nm處吸光值.
1.2.3EdU檢測細胞增殖將細胞接種至6孔板中,在細胞生長到匯合度約為70%~90%時轉染pCMV-HA或pCMV-HA-載體,6 h后更換新鮮的培養基,于37 ℃ 5% CO2培養箱中繼續培養24 h后進行EdU孵育Hoschst染色,并于熒光顯微鏡下觀察[17].
1.2.4Western blot驗證過表達效果將雞前脂肪細胞接種至6孔板,分別轉染pCMV-HA-TNNI2以及pCMV-HA質粒,轉染48 h后,使用RIPA裂解液(含1% PMSF)(Beyotime)提取細胞中的總蛋白,4 ℃ 12 000 r/min離心5 min,上清即為總蛋白,加入5×蛋白上樣緩沖液(Beyotime)煮沸5 min后進行SDS-PAGE電泳,電泳結束后將蛋白樣品轉移至PVDF膜(Millipore),室溫封閉1 h,anti-HA標簽抗體孵育1 h,二抗孵育1 h.采用Odyssey雙紅外激光成像系統檢測TNNI2蛋白表達水平.拍照完成后,采用一抗二抗洗脫液(Beyotime)洗脫1 h,采用內參抗體anti--actin孵育1 h,二抗孵育1 h,檢測內參蛋白-actin的表達水平.
1.2.5RNA提取及qRT-PCR待ICP1細胞生長匯合度80%~90%提取RNA,向培養基中加入適量的RNAiso Plus后混勻,冰上靜置5 min后,12 000 r/min離心5 min,轉移至新1.5 mL離心管中,加入適量體積的三氯甲烷,渦旋振蕩至溶液呈乳白色,將上清液轉移至新的離心管中,再加入等體積的異丙醇并混合均勻,冰上靜置10 min后,12 000 r/min離心10 min,加入適量75%乙醇,晃動離心管并7 500 r/min離心5 min,去上清液保留沉淀,待離心管干燥后,加入30 μL DEPC水溶解RNA,即可作為模板進行反轉錄.反轉錄按照HiScript?II Q Select RT SuperMix for qPCR(+gDNA wiper)(Vazyme)說明書進行操作.首先去除基因組的DNA,反應體系為:總RNA 0.5 μg,4×DNA wipe Mix 2 μL,加入RNase free ddH2O,總體系為8 μL.體系用移液器輕輕吹打混勻,42 ℃ 2 min.反轉錄時向體系中加入5×HiScript?II qRT SuperMix II 2 μL,用移液器吹打均勻,25 ℃ 10 min,50 ℃ 30 min,85 ℃ 5 min,置于-20 ℃冰箱中保存.qRT-PCR反應按照ChamQTMSYBR?qPCR Master Mix(Without ROX)(Vazyme)說明書進行操作.反應體系:2×ChamQ SYBR qPCR Master Mix 5 μL,cDNA模板 1 μL,上、下游引物(10 μmol/L)各0.2 μL,ddH2O 3.6 μL.反應程序:95℃預變性30s,95 ℃變性10s,55 ℃復性延伸30s,共40個循環.每個樣品設置3個重復.以無POU域八聚體結合蛋白(non-POU doman containing octamer-binding protein,NONO)為內參照基因.利用2-ΔΔCt算法將原始Ct值轉換為基因相對表達量.所用引物序列見表1.
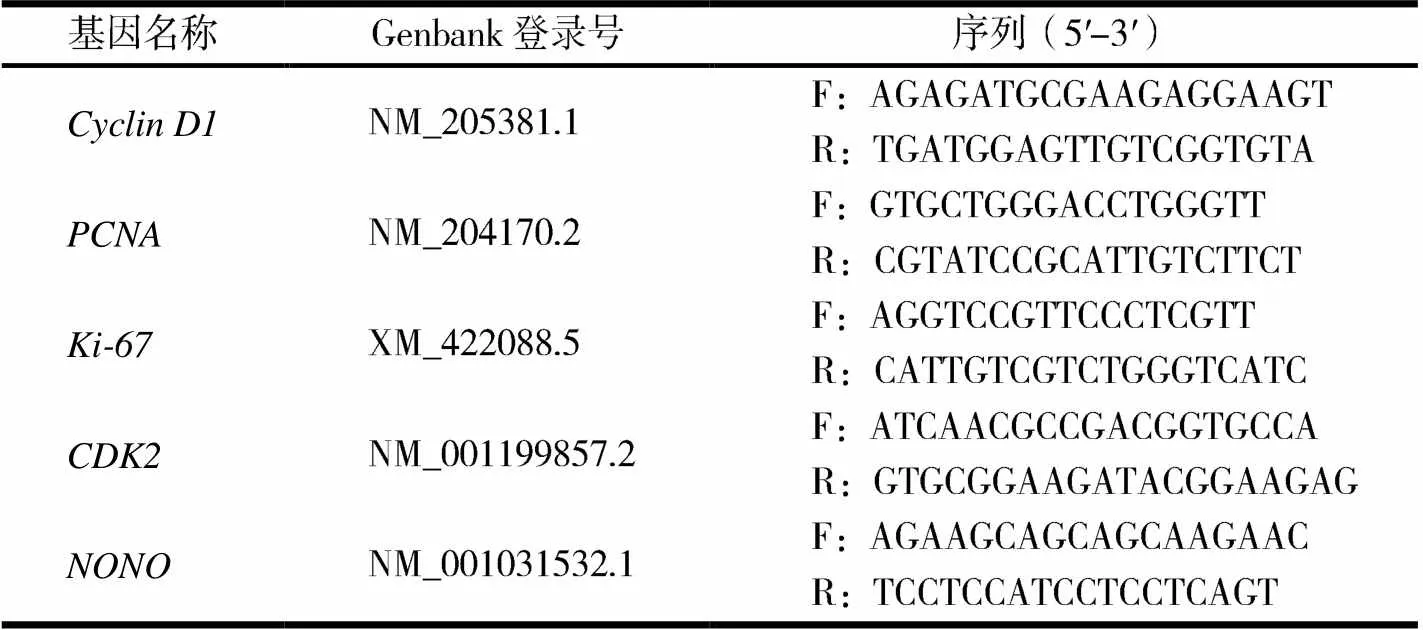
表1 qRT-PCR引物
1.2.6統計分析使用SPSS22.0(IBM Corporation.,Armonk,NY;USA)對實驗數據進行統計分析,使用Graphpad Prism 7.0(Graphpad Software Inc.,San Diego,CA,USA)軟件進行圖形繪制.使用檢驗比較組間差異,<0.05為顯著性差異,<0.01為極顯著性差異,具有統計學意義.
2 實驗結果
2.1 ICP1細胞正常增殖過程中TNNI2基因的表達
復蘇ICP1細胞并分別傳代至6孔板上,待細胞完全貼壁記為0 h,分別于24,48,72 h提取總RNA,采用qRT-PCR法檢測基因在ICP1細胞增殖過程中的表達量變化(見圖1).由圖1可見,在24,48,72 h均有表達且呈現出先下降后上升的趨勢(差異不顯著).表明TNNI2可能對ICP1細胞的增殖起到一定作用.

圖1 ICP1細胞增殖過程中TNNI2 mRNA的表達
2.2 TNNI2過表達效果驗證
利用anti-HA標簽抗體進行Western blot檢測轉染過表達載體pCMV-HA-及空載體pCMV-HA細胞中HA蛋白的表達情況.結果表明,在轉染了pCMV-HA-質粒的ICP1細胞表達出一條約23 kD的特異蛋白條帶,而轉染pCMV-HA質粒的細胞沒有表達出23 kD的特異帶(見圖2),表明真核表達載體pCMV-HA-能在ICP1細胞中成功表達HA-TNNI2蛋白.
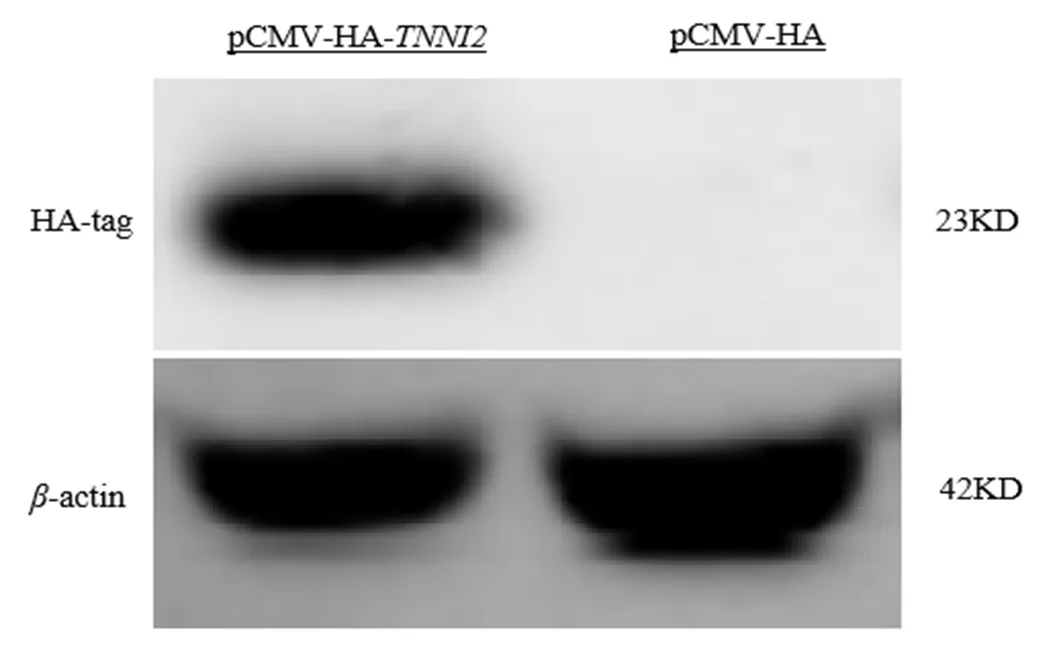
圖2 western blot分析ICP1細胞中TNNI2蛋白的表達
2.3 過表達TNNI2對雞前脂肪細胞增殖的影響
CCK-8實驗結果表明,與轉染pCMV-HA的細胞相比,轉染pCMV-HA-的細胞在450 nm處的吸光值明顯上升,48 h差異顯著(<0.05,見圖3a).EdU實驗結果表明,與轉染pCMV-HA質粒的細胞相比,轉染pCMV-HA-質粒的細胞EdU摻入的細胞數明顯上升(<0.01,見圖3b).qRT-PCR實驗結果表明,轉染了pCMV-HA-質粒后,ICP1細胞的增殖標志因子,,,的mRNA水平雖有上升,但均無顯著變化(>0.05,見圖3c).實驗結果表明,過表達TNNI2對雞前脂肪細胞的增殖有促進作用.
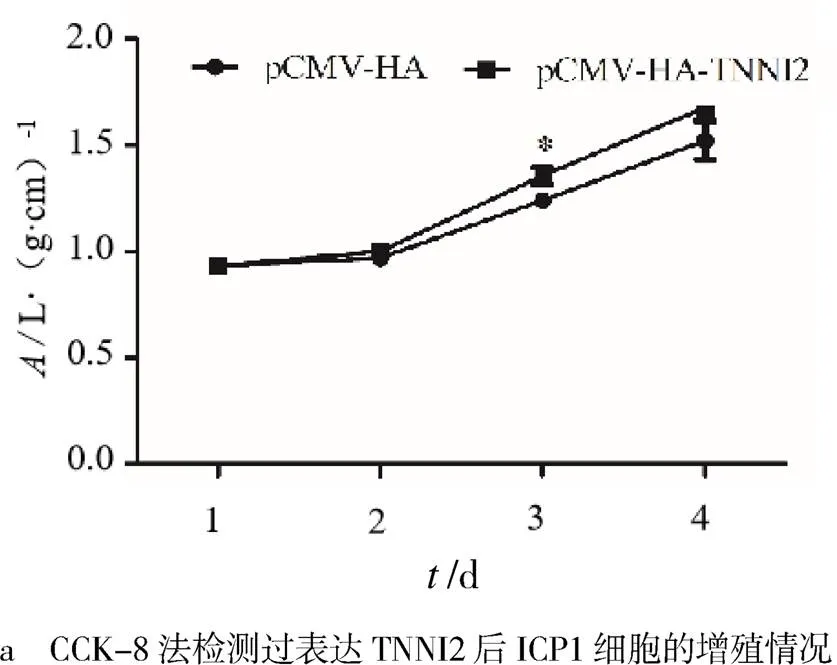
3 結論與討論
肌鈣蛋白I(TnI)是一種可以在肌鈣蛋白復合物橫紋肌細纖維中起到抑制作用的亞基,并且在鈣調節的肌肉收縮和松弛中起核心作用.脊椎動物TnI分為3個亞型,TNNI1(編碼慢骨骼肌)、TNNI2(編碼快骨骼肌)、TNNI3(編碼心肌),目前TNNI2在畜類動物肌肉組織中的作用已被證實,但其在禽類脂肪組織形成過程的功能還不清楚.
KLF7是脂肪細胞分化的負調控因子,實驗室前期以雞前脂肪細胞為研究對象,進行染色質免疫共沉淀結合測序(ChIP-seq)分析,并首次發現在基因5′側翼區存在一個KLF7結合位點,提示可能是KLF7的一個潛在靶基因[18],KLF7促進哺乳動物和禽類前脂肪細胞增殖,所以推測TNNI2對細胞增殖具有促進作用.本研究首先檢測了基因的本底表達水平,結果顯示,48 h前的表達量呈逐漸降低的趨勢,這可能是細胞剛剛復蘇狀態不佳,未達到理想的增殖狀態所導致,48 h后的表達水平逐步上升,提示TNNI2可能對雞前脂肪細胞的增殖有促進作用.為了驗證這一假設,在ICP1細胞中進行了過表達實驗,并在蛋白質水平驗證了過表達效果,結果表明,TNNI2過表達載體構建成功.進一步采用CCK-8和EdU實驗檢測細胞增殖,實驗結果顯示,過表達組的細胞增殖水平都顯著高于空白對照組,這些結果說明TNNI2促進雞前脂肪細胞增殖.合成了3條干擾片段,但是沒有成功干擾TNNI2基因的表達.今后有必要利用Crispr cas9技術在體內和體外開展TNNI2在雞前脂肪細胞中功能的研究.
雞前脂肪細胞增殖會直接影響到雞脂肪細胞的數量,過多的脂肪細胞會導致脂肪組織過度蓄積,雞體內脂肪過度蓄積會影響雞的產蛋率、受精率等一系列問題.前脂肪細胞增殖受復雜的轉錄因子網絡調控.BMP4作為前脂肪細胞增殖的正調控因子通過促進G1/S的轉化來促進前脂肪細胞增殖,另外PPAR[19],RB1[20],HOPX[21],TCF21[22]等因子對前脂肪細胞增殖起到負調控作用.然而,本研究在過表達TNNI2之后,檢測增殖標志基因的表達發現,等增殖標志基因的表達量略有升高,但均無顯著性,這可能是由于轉染效率不高或基因并不通過這些標志基因來影響細胞增殖所導致.有研究發現,編碼的fsTnI蛋白的亞細胞定位于細胞核.也有研究認為,在哺乳動物體內fsTnI蛋白可以于核內與雌激素受體相關受體a(estrogen receptor related receptor a,ERRa)結合,以共激活劑的形式增強其在乳腺細胞中的交互作用[23].因此,本研究推測TNNI2可能作為轉錄因子在核內對ICP1細胞的增殖起到促進作用.基因在雞體內脂肪組織發育中的作用仍有待研究,在后續的研究中有必要開展基因干擾實驗以及流式細胞術檢測細胞周期以確定TNNI2以何種方式促進ICP1增殖,繼續驗證TNNI2在體內和體外的功能.
綜上所述,過表達基因可以有效促進ICP1細胞增殖.
致謝:感謝東北農業大學農業農村部雞遺傳育種重點實驗室王寧教授提供永生化雞前脂肪細胞系.
[1] 樊紅平,侯水生.家禽體內脂肪沉積調控的研究進展[J].動物營養學報,2004(4):1-6,12.
[2] 李云雷,孫研研,陳繼蘭.脂肪沉積對家禽繁殖性狀影響的研究進展[J].中國家禽,2016,38(13):36-41.
[3] 鞠大鵬,詹麗杏.脂肪細胞分化及其調控的研究進展[J].中國細胞生物學學報,2010,32(5):690-695.
[4] SHENG J J,JIN J P.TNNI1,TNNI2 and TNNI3:Evolution,regulation,and protein structure-function relationships[J].Gene,2016,576(1 Pt 3):385-394.
[5] 李伯江,李平華,吳望軍,等.骨骼肌肌纖維形成機制的研究進展[J].中國農業科學,2014,47(6):1200-1207.
[6] BERSON G,SAMUEL J L,SWYNGHEDAUW B.A comparative study of the cardiac troponin inhibitory factor(TNI)from mammalians[J].Pflugers Arch,1978,374(3):277-283.
[7] MOSES M A,WIEDERSCHAIN D,WU I,et al.Troponin I is present in human cartilage and inhibits angiogenesis[J].Proc Natl Acad Sci U S A,1999,96(6):2645-2650.
[8] FU Z,LIANG X,SHI L,et al.SYT8 promotes pancreatic cancer progression via the TNNI2/ERRalpha/SIRT1 signaling pathway[J].Cell Death Discov,2021,7(1):390.
[9] 鄭昭璟,傅啟華.應用全外顯子組測序技術進行遺傳性骨病分子缺陷機制的研究[D].上海:上海交通大學,2014.
[10] ZHU X,WANG F,ZHAO Y,et al.A gain-of-function mutation in Tnni2 impeded bone development through increasing Hif3a expression in DA2B mice[J].PLoS Genet,2014,10(10):e1004589.
[11] WANG B,ZHENG Z,WANG Z,et al.A novel missense mutation of TNNI2 in a Chinese family cause distal arthrogryposis type 1[J].Am J Med Genet A,2016,170A(1):135-141.
[12] PARK B S,OH Y K,KIM M J,et al.Skeletal Muscle Troponin I(TnI)in Animal Fat Tissues to Be Used as Biomarker for the Identification of Fat Adulteration[J].Korean J Food Sci Anim Resour,2014,34(6):822-828.
[13] DUPONT J,METAYER-COUSTARD S,JI B,et al.Characterization of major elements of insulin signaling cascade in chicken adipose tissue:apparent insulin refractoriness[J].Gen Comp Endocrinol,2012,176(1):86-93.
[14] JI B,ERNEST B,GOODING J R,et al.Transcriptomic and metabolomic profiling of chicken adipose tissue in response to insulin neutralization and fasting[J].BMC Genomics,2012,13(441):1-16.
[15] MELLOUK N,RAME C,BARBE A,et al.Chicken Is a Useful Model to Investigate the Role of Adipokines in Metabolic and Reproductive Diseases[J].Int J Endocrinol,2018,19:1-19.
[16] 李金煒,史洪巖,譚茗,等.雞基因的真核表達載體構建及功能生物信息學分析[J].云南農業大學學報:自然科學,2021,36(3):430-437.
[17] 靳文姣,翟彬,苑鵬濤,等.miR-215-5p通過靶向NCOA3基因抑制固始雞腹部前脂肪細胞的增殖和分化[J].畜牧與獸醫,2021,53(7):69-77.
[18] 孫嬰寧.KLF7 在雞前脂肪細胞中的靶基因篩選和鑒定[D].哈爾濱:東北農業大學,2016.
[19] WANG L,NA W,WANG Y X,et al.Characterization of chicken PPARgamma expression and its impact on adipocyte proliferation and differentiation[J].Yi Chuan,2012,34(4):454-464.
[20] 張琦,黃嬌嬌,楊彩俠,等.CRISPR/Cas9介導基因敲除及其在雞前脂肪細胞分化、增殖中的功能研究[J].畜牧獸醫學報,2016,47(9):1775-1784.
[21] SHI H Y,QI H E,CHENG M,et al.Effect of HOPX Gene Overexpression on Chicken Preadipocyte Proliferation[J].Scientia Agricultura Sinica,2015,48:1624-1631.
[22] ZHANG X,CHENG B,LIU C,et al.A Novel Regulator of Preadipocyte Differentiation,Transcription Factor TCF21,Functions Partially Through Promoting LPL Expression[J].Front Physiol,2019,10:458.
[23] LI Y,CHEN B,CHEN J,et al.Fast skeletal muscle troponin I is a co-activator of estrogen receptor-related receptor alpha[J].Biochem Biophys Res Commun,2008,369(4):1034-1040.
Effects of fast skeletal muscle troponinⅠon proliferation of chicken preadipocytes
KONG Lingzhe,LI Jinwei,ZHANG Xinyu,DANZENG Jinmei,SUN Ze,SUN Yingning
(School of Life Sciences,Agriculture and Forestry,Qiqihar University,Qiqihar 161006,China)
Our previous studies showed that KLF7 promotes the proliferation of chicken preadipocytes and regulates the expression ofgene,but the role of TNNI2 in proliferation of preadipocytes remains unclear.Hence,the present study investigated the effect ofon the proliferation of chicken preadipocytes.The expression ofgene during the proliferation of immortalized chicken preadipocytes(ICP1)was detected by qRT-PCR.TNNI2 overexpression vector PCMV-HA-was transfected into ICP1 cells.Western blot assay was used to detect the overexpression effect.Cell proliferation was detected by CCK-8 and EdU methods.The expression of proliferation marker gene was detected by qRT-PCR.Thegene decreased first and then increased gradually during ICP1 cells proliferation.Western blot results showed thatgene was overexpressed successfully.CCK-8 results showed that the absorbance value of ICP1 cells at 450 nm increased significantly after TNNI2 overexpression(<0.05).EdU results showed that after TNNI2 overexpression,the percentage of EdU incorporation in the total number of cells in the experimental group was significantly higher than that in the control group(<0.01).qRT-PCR results showed that the expression levels of proliferative marker genes were up-regulated but the difference was not significant(>0.05).gene can promote the proliferation of chicken preadipocytes.
chicken;preadipocyte;overpression;TNNI2;proliferation
1007-9831(2022)10-0048-06
Q955
A
10.3969/j.issn.1007-9831.2022.10.010
2022-05-23
國家自然科學基金項目(31402061);2022年度黑龍江省省屬高等學校基本科研業務費科研項目(145109211);齊齊哈爾大學創新創業訓練計劃項目(202220232304)
孔令喆(1999-),男,黑龍江大慶人,在讀碩士研究生,從事動物分子遺傳研究.E-mail:konglz121@126.com
孫嬰寧(1981-),女,遼寧撫順人,教授,博士,從事肥胖及脂肪組織發育相關研究.E-mail:yingningsun@163.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
