武昌湖退化濕地菰草根際水體細菌多樣性
2022-11-08 05:14:54劉趙文汪雨羊禮敏葉姍程起航
高師理科學刊 2022年10期
關鍵詞:環境
劉趙文,汪雨,羊禮敏,葉姍,程起航
武昌湖退化濕地菰草根際水體細菌多樣性
劉趙文1,汪雨2,羊禮敏1,葉姍1,程起航1
(池州學院 1. 材料與環境工程學院,2. 地理與規劃學院,安徽 池州 247000)
為研究濕地細菌群落結構及功能與退化濕地環境之間的關系,選取武昌湖濕地菰草()根際水體環境樣本,提取樣本細菌基因組DNA,并利用16S rDNA基因進行454高通量測序,共得到優質序列17 051條,產生3 032條OUT.結果表明,武昌湖退化濕地菰草根際水體細菌群落結構涉及15門50屬,其中以變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、放線細菌門(Actinobacteria)、藍細菌(Cyanobacteria)為主要門類;菰草根際不同水體環境細菌群落結構存在顯著差異,生物多樣性指數Shannon,Chao,ACE指數及OTUs的變化特征均表明菰草介入導致其根際水體細菌群落多樣性降低;濕地微環境下菌種組成與環境功能之間有密切的聯系.探究濕地退化過程中植被菰草根際細菌群落結構特征,對揭示長江中下游淺水湖泊濕地退化機理具有重要意義.
濕地;菰草;細菌多樣性;高通量測序;武昌湖
濕地作為同時兼顧水生和陸生環境的一種特殊生境,面積雖然不到陸地面積的1/10,卻承載了地球上1/5已知生物的生存環境[1].濕地生態系統具有調節生物多樣性、氣候和土壤碳儲存等重要的生態系統服務功能[2].武昌湖濕地位于長江中下游北岸,近年來湖面大面積菰草()繁殖導致濕地退化現象非常嚴重,植物群落數量均極大降低,使植物群落趨向同質化,生物多樣性遭到破壞[3-4].
微生物作為濕地中的主要分解者,參與了根際微環境N,P元素的循環和利用,直接影響著植物生長發育等[5],根際微生物可有效提高植物對土壤氮素的吸收和利用[6].植物入侵會對濕地土壤理化性質和土壤微生物群落產生顯著影響.研究表明,植物入侵會改變土壤性質以及土壤微生物群落結構等[7].鄭潔[8]等研究發現互花米草入侵使紅樹林土壤質量發生明顯退化,土壤微生物群落多樣性指數明顯下降.Cui[9]等在研究互花米草入侵沿海濕地時發現植物入侵使硫酸鹽還原菌群落的豐富度和多樣性顯著提高.Liu[10]等在研究紅樹林濕地植物入侵時發現,植物入侵改變了土壤細菌群落組成和功能.YANG[11]等研究發現,植物入侵使土壤真菌功能類群發生了改變,腐生真菌的相對豐度顯著增加,共生真菌和病原性真菌的相對豐度顯著降低.
根際微生物進行代謝不斷向體外分泌養分和低分子有機物等,極大地提高了植物對養分的利用效率,因此微生物多樣性對植物的生長、發育、群落結構演替具有重要的作用[12].但從研究對象來看,目前學者研究植物根際土壤微生物較多,而關于植物根際水體微生物的研究鮮有報道.通過研究入侵植物菰草根際水體細菌群落結構多樣性,探究濕地退化過程中植被根際微生物群落結構特征,對揭示長江中下游淺水湖泊濕地退化機理具有一定的借鑒意義.
1 材料與方法
1.1 研究區概況
武昌湖濕地位于安徽省西南部安慶市望江縣東北部,東經116°26′~116°53′,北緯30°9′~30°24′,屬北亞熱帶季風濕潤氣候區,這里四季分明,全年平均氣溫約為16.5 ℃,雨水充足,全年平均有130~150 d為降水日,降水量可達到1 400 mm,全年日照可達到2 052~2 065 h.該濕地系斷裂帶局部凹陷洼地,經長江泥沙長期淤塞、積水而成的淡水湖濕地是長江中下游北岸古雷池遺跡的重要組成部分.景區水域面積達102.5 km2,水生資源極為豐富,其生態作用主要體現在調節氣流、改善氣候、區域旅游、漁業養殖等[13].武昌湖濕地區位與菰草種群空間分布情況見圖1.武昌湖濕地是以草本沼澤和湖泊為主的一塊濕地,其東西長約20 km,南北長約7.5 km,1993年修建的跨湖公路將武昌湖分割為上湖和下湖2個區域.濕地植被建群種主要以挺水植物——菰草為主,高度為0.8~2.1 m,生長茂盛,蓋度常可達60%~70%,其它伴生種主要有蓮子草、愧葉萍、丘角菱、水鱉、浮萍等.

圖1 武昌湖濕地區位與菰草種群空間分布圖
1.2 樣品采集
采樣前期通過查閱相關文獻及實地多次觀察發現,武昌湖濕地菰草部分群落在每年的豐水期會大面積漂浮在水上,隨著水位下降,種群會漂到不同的環境下重新扎根生長.本文選取武昌湖濕地菰草群落重新扎根在不同的微環境下生長后的菰草根際水體作為研究對象.結合現實可操作性以及該區域的地形地勢,選取了武昌湖濕地4個樣點作為釆樣區域,采樣點的基本概況見表1.采樣時使用GPS衛星定位,記錄采樣區域的地理位置,各樣點優先選擇采集水樣.采集水樣時先將藍蓋瓶用采樣點水樣潤洗幾遍后再取水樣1 000 mL.

表1 采樣點基本概況
1.3 樣本理化參數測定
水體濁度的測定(GB 13200—91)、水體總氮的測定(GB HJ 636—2012)、水體氨氮的測定(GB HJ 536—2009)、水體總磷的測定(GB 11893—89)、水體化學需氧量(CODcr)的測定(GB 11914—89).
1.4 樣品基因組DNA的提取及高通量測序
采用PowerSoil?DNAIsolation試劑盒分別提取菰草根際水體環境樣本總DNA,樣本經0.22 μm濾膜負壓過濾收集水體細菌后,用干冰保存樣品,并寄至上海美吉生物公司完成DNA抽取和測序.利用Nanodrop-2000檢測總DNA濃度,并利用1%瓊脂糖凝膠電泳檢測DNA質量.分別對各環境樣本總DNA進行16S rRNA基因PCR擴增,所采用的引物為338F:5′-ACTCCTACGGGAGGCAGCAG-3′和806R:5′-GGACTACHVGGGTWTCTAAT-3′.PCR擴增體系包含2 μL dNTP(2.5 mmol/L)、0.8 μL正/反向引物(5 μmol/L)、0.4 μL 5×FastPfu緩沖液、10 ng模板DNA、0.2 μL BSA、0.4 μL FastPfu聚合酶,并加ddH2O至反應體積為20 μL.擴增條件:95 ℃預變性3 min,95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸45 s,28次循環,最后72 ℃延伸10 min.利用2%瓊脂糖凝膠電泳檢測PCR產物,并用純化試劑盒純化,然后對目標產物用Illumina進行高通量測序并分析.
2 結果與討論
2.1 武昌湖退化濕地菰草生長水環境
菰草根際水體各樣點的理化性質見表2.由表2可見,武昌湖濕地菰草根際水體基本偏弱酸性.菰草根際水體樣品間濁度大小為W3>W4>W2>W1,這可能是由于采樣點間的微環境不同,從而導致各水樣間濁度存在一定差異.

表2 武昌湖濕地菰草根際水體理化性質
水中總氮、總磷和氨氮的含量是水生植物最主要的營養物質來源,其含量的多少是直接導致水質污染現象的最主要原因之一.菰草根際水體總氮含量的大小為W3>W2>W1>W4,總磷含量的大小為W3>W1>W2>W4,氨氮含量的大小為W3>W4>W2>W1.根據我國地表水環境質量標準(GB 3838—2002)發現,總氮含量W1,W4水樣符合三類水(≤1.0 mg/L)的標準,W2,W3水樣符合四類水(≤1.5 mg/L)標準;氨氮含量除了W3水樣屬于四類水(≤1.5 mg/L)標準,其它3組水樣屬于三類水(≤1.0 mg/L)標準;總磷含量只有W4符合三類水(≤0.05 mg/L)標準,其它3組屬于四類水(≤0.1 mg/L)標準;化學需氧量除了W2外,其它3組屬于四類水(≤30 mg/L)標準.W3水樣中的營養元素含量高于其它組,這可能與該環境中菰草常年漂浮生長在水面上有關,水體中的營養元素基本上成為該環境下菰草生長的唯一來源,使菰草根際某些微生物可能對營養元素具有一定的富集作用.這一發現與相關學者研究結果具有一定的相似性,如Eileen[14]等在研究斑點矢車菊過度繁殖時發現,其根際的菌根菌通過地下菌絲網絡將碳水化合物等由本土植物向斑點矢車菊根際運輸,從而抑制了本土植物的生長,促進了斑點矢車菊在河漫灘地的過度生長.Fumanal[15]等在研究植物豚草時發現,接種其根際的叢枝菌根真菌能顯著增加豚草的葉片數、株高以及總生物量干質量等,促進了其種群的進一步擴大.植物紫莖澤蘭在入侵我國西南部時,重度入侵區土壤中總磷、硝態氮的含量及土壤中自生固氮菌的數量和種類的多樣性都顯著高于輕度入侵區和未入侵區[16-17].而水樣其它的理化性質具有一定的差異性,很可能跟采樣點的選取及采樣點微環境的不同存在一定聯系.
2.2 菰草根際水環境中微生物群落多樣性分析
隨機抽樣測試序列,并基于所抽到獲得的序列數表征其可代表的數目,構建稀釋性曲線(見圖2).就稀釋曲線分析4個水環境樣本基本達到0.97%,總體處于平坦的狀態,但是尚未能飽和.這說明樣品中包含很多與數據樣本有關的信息,能夠總體說明濕地細菌群落情況.Rank-abundance曲線的作用主要體現在說明物種的豐富程度及其均勻程度,一般可用曲線形狀說明,如果曲線較為平坦則說明其所對應的物種組成具有較高的均勻度;通過橫軸上曲線的長度可說明物種的豐富程度,如果該曲線越寬,則表征具有越為豐富的物種組成.通過W4水樣的曲線說明采樣點具有較高的物種豐富度,其中W2的物種豐富程度最小,這一結果與上面根據多樣性指數所得到的結果基本保持一致,存在小的差異可能是由于不同計算方法之間存在一定的誤差所導致.

圖2 武昌湖濕地水樣Rank-abundance曲線
運用Mothur軟件基于97%分類,計算出其多樣性生物指數(見表3).由表3可見,不同水體微環境下,OTUs的大小為W4>W1>W3>W2,細菌多樣性Chao指數大小為W4>W1>W2>W3,ACE指數大小為W4>W2>W1>W3,Shannon指數大小為W4>W3>W1>W2,Simpson指數大小為W2>W1>W3>W4.利用16S rDNA基因進行454高通量測序,在97%相似度水平下,利用Mothur軟件對各環境樣本進行分析發現,武昌湖濕地水體細菌菌屬按照W4>W3>W1>W2規律依次遞減.退化濕地菰草水環境中細菌多樣性十分豐富,共涉及15門,50個屬.

表3 武昌湖濕地菰草水環境細菌群落豐富度及多樣性
OTUs分布維恩圖見圖3.由圖3可見,3種微環境下相同的OTUs為155個,約占OTUs總數的9.5%;從OTUs分布情況來看,W1和W2的相似性最大,共有389個相同OTUs,占兩者OTUs總數的54%;其次是W1和W4兩者共有483個相同OTUs,占兩者OTUs總數的34%;W2和W3共有274個相同OTUs,占兩者OTUs總數的32%;W1和W3共有309個相同OTUs,占兩者OTUs總數的31%;W2和W4共有357個相同OTUs,占兩者OTUs總數的26%;W3和W4為4組樣品兩兩之間相似性最低的,共有327個相同OTUs,占兩者OTUs總數的23%.

圖3 武昌湖濕地菰草水環境樣本細菌群落OTUs分布維恩圖
通過測試分析表明,武昌湖菰草水環境細菌在門分類水平上(見圖4)絕對優勢菌門為變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、放線細菌門(Actinobacteria)、藍細菌(Cyanobacteria).但是各菌門豐度在各樣品間存在一定差異,變形菌門在各樣本中相對豐度為W2(75.19%)>W4(66.03%)>W1(48.45%)>W3(21.02%),擬桿菌門在各樣品中相對豐度為W1(28.91%)>W3(17.38%)>W4(8.42%)>W2(7.98%),放線細菌門在樣品中相對豐度為W3(28.93%)>W1(13.36%)>W2(11.56%)>W4(7.85%),藍細菌在樣品中相對豐度為W3(22.37%)>W1(3.36%)>W4(2.25%)>W2(2.06%).

圖4 武昌湖濕地菰草水環境樣本細菌群落結構(門水平上)組分圖
2.3 菰草根際水體優勢菌種環境功能分析
菰草根際水體樣本細菌群落Heatmap圖見圖5.由圖5可見,菰草根際水體在不同微環境下細菌在屬水平上的優勢菌種作為W1文庫中第一類優勢菌屬,其比例高達26.69%,而且Hahn[18]等發現該菌屬于化學有機營養的兼性厭氧微生物,廣泛存在于淡水和湖泊中,具有同化乙酸鹽、甘油酸鹽和葡萄糖酸鹽等功能.第二類是廣泛分布于土壤和水中的典型有機化能營養型的好氧反硝化細菌——黃桿菌屬,該菌在文庫中的比例為13.67%.而第三大類叢毛單胞菌屬在文庫中比列為6.69%,研究表明該菌屬可用于活性污泥處理廢水的脫氮除磷處理工藝,不僅能夠將蛋白質分解成氨基酸,而且還能分解苯酚、有機酸和氨基酸[19].作為W2文庫中第一類優勢菌屬(29.91%),Kim[20]等發現其廣泛存在于湖泊、淡水沉積物和沼澤中,該細菌嚴格好氧且主要以甲烷、甲醇等含甲基單碳化合物作為碳源和能源.(10.80%)兼性甲基營養菌細胞呈橢圓形或桿狀.除了甲胺外,還可以利用果糖、甜菜堿、甲醇、丙酮酸和乙醇作為碳源,利用銨、硝酸鹽和尿素可作為氮的來源[21].是一種兼性厭氧化自養菌,以硫化物、元素硫、硫代硫酸鹽和氫作為電子供體,能夠利用不同的硫種類和硝酸鹽[22].W3文庫中最主要優勢菌屬是.W4文庫中優勢菌屬為(14.25%),紅環菌在文庫中的比例為9.59%,該類微生物能夠分解不同有機物獲得碳源進行生存,其在環境有機污染物降解方面具有較好的應用前景[23].革蘭氏陰性需氧細菌(6.95%)在結構和功能上都專門利用甲烷作為唯一的能量和碳源,該細菌能夠適應自然生態位中的顯著溫度變化[24].
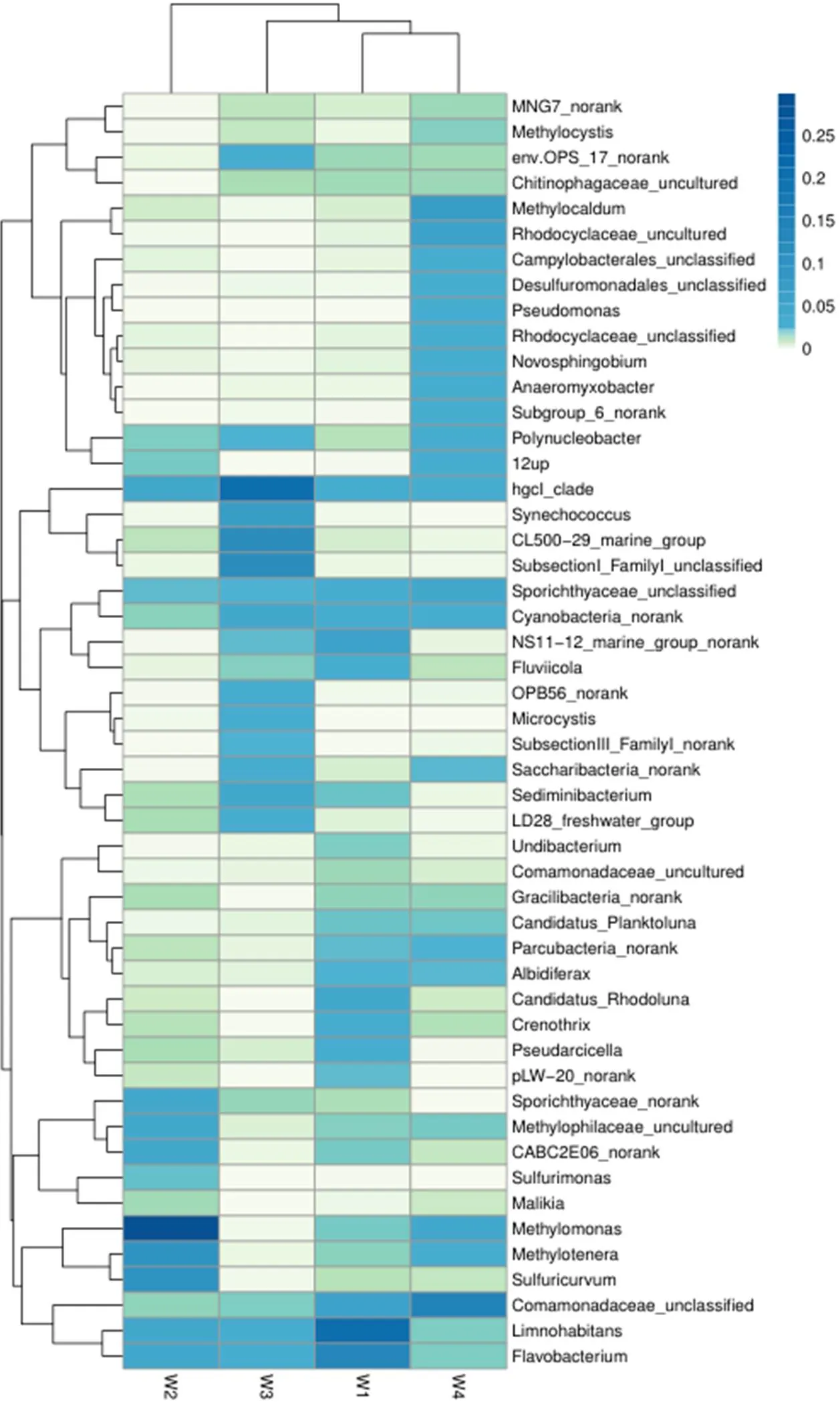
圖5 武昌湖濕地水環境樣本細菌群落Heatmap圖
菰草根際水體不同微環境細菌群落結構存在明顯差異.W1優勢菌種為Proteobacteria,Actinobacteria,Bacteroidetes,3種之和占檢測總門數的94.08%;W2優勢菌種為Proteobacteria,Actinobacteria,2種之和占檢測總門數的86.78%;W3優勢菌種為Proteobacteria,Bacteroidetes,Actinobacteria,Cyanobacteria,4種之和占檢測總門數的89.7%;W4優勢菌種為Proteobacteria,該菌門占檢測總門數的66.03%.盡管Proteobacteria在4種水體微環境中都是絕對優勢菌種,但在不同的環境中所占的比例各不相同,在W3(21.02%),W1(48.45%),W4(66.03%),W2(75.19%)中的比例逐漸增大;4組水體樣品生物多樣性指數表明了各樣本的細菌間存在差異性豐度,這一點通過OTU聚類即可進一步驗證說明,W4樣品的Shannon指數最大,W3和W1樣品的Shannon指數大小差別不大,W2的Shannon指數較小,這說明W4樣本多樣性最高,這一點通過Simpson指數也能夠得到驗證4組樣品細菌的多樣性高低為W4>W3>W1>W2.
3 結論
不同環境下根際水體細菌Shannon指數大小為3.78~5.78,根際水體的細菌絕對優勢菌門為變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、放線細菌門(Actinobacteria)、藍細菌(Cyanobacteria);退化濕地菰草水環境中細菌多樣性十分豐富,共涉及15門,50個屬.菰草根際不同水體環境細菌群落結構存在顯著差異,生物多樣性指數Shannon,Chao,ACE指數及OTUs的變化特征均表明菰草介入導致其根際水體細菌群落多樣性降低,濕地微環境下菌種組成與環境功能之間有密切的聯系.不同微環境下菰草根際細菌多樣性存在一定差異,濕地微環境下細菌組成與環境功能之間有密切的聯系,它們主要參與脫氮除磷,降解大分子有機物和重金屬以及為植物群落的演替提供了良好的生境.
[1] 周雅心,陳鈺,鄭毅,等.中國典型濱海濕地轉變為養殖塘對土壤細菌多樣性及群落結構的影響[J].環境科學學報,2021,41(6):2402-2413.
[2] 肖濤,石強勝,聞熠,等.濕地生態系統服務研究進展[J]. 生態學雜志,2022,41(6):1205-1212.
[3] 周葆華,尹劍,金寶石,等.30年來武昌湖濕地退化過程與原因[J].地理學報,2014,69(11):1697-1706.
[4] 劉趙文,周葆華,寇樂勇,等.武昌湖典型退化濕地菰草根際微生物群落結構[J].地球與環境,2018,46(4):339-347.
[5] Nico E,Stefan S,Alexandre J,et al.Bacterial Diversity Stabilizes Community Productivity[J].PLoS ONE,2012,7(3):e34517.
[6] Cole L,Staddon P L,Sleep D,et al.Soil animals influence microbial abundance,but not plant microbial competition for soil organic nitrogen[J].Functional Ecology,2004,18(5):631-640.
[7] Montserrat Vilà,José L Espinar,Hejda M,et al.Ecological impacts of invasive alien plants:A meta-analysis of their effects on species,communities and ecosystems[J].Ecology Letters,2011,14(7):702-708.
[8] 鄭潔,劉金福,吳則焰,等.閩江河口紅樹林土壤微生物群落對互花米草入侵的響應[J].生態學報,2017,37(21):7293-7303.
[9] Cui J,Chen X,Nie M,et al.Effects of Spartina Alterniflora Invasion on the Abundance,Diversity,and Community Structure of ulfate Reducing Bacteria along a Successional Gradient of Coastal Salt Marshes in China[J].Wetlands,2017,37(2):221-232.
[10] Liu M,Yu Z,Yu X,et al.Invasion by cordgrass increases microbial diversity and alters community composition in a mangrove nature reserve[J].Frontiers in Microbiology,2017,8:2503.
[11] YANG W,JEELANI N,XIA L,et al.Soil fungal communities vary with invasion by the exotic Spartina alternifolia Loisel in coastal salt marshes of eastern China[J].Plant and Soil,2019,442(1/2):215-232.
[12] 趙官成,梁健,淡靜雅,等.土壤微生物與植物關系研究進展[J].西南林業大學學報,2011,31(1):83-88.
[13] 周葆華,操璟璟,朱超平,等.安慶沿江湖泊濕地生態系統服務功能價值評估[J].地理研究,2011,30(12):2296-2304.
[14] Eileen V Carey,Marilyn J Marler,Ragan M Callaway.Mycorrhizae transfer carbon from a native grass to an invasiveweed:evidence from stable isotopes and physiology[J].Plant Ecology,2004,172(1):133-141.
[15] Fumanal B,Plenchette C,Chauvel B,et al.Which role can arbuscular mycorrhizal fungi play in the facilitation of Ambrosia artemisiifolia L. invasion in France?[J].Mycorrhiza,2006,17(1):25-35.
[16] Xu Chanwen,Yang Mingzhi,Chen Yunjiao,et al.Changes in non-symbiotic nitrogen-fixing bacteria inhabiting rhizosphere soils of an invasive plant Ageratina adenophora[J].Applied Soil Ecology,2012,54:32-38.
[17] Xin S,Cheng G,Guo L D.Changes in soil microbial community and enzyme activity along an exotic plant Eupatorium adenophorum invasion in a Chinese secondary forest[J].Science Bulletin,2013,58(33):4101-4108.
[18] Hahn M W,Kasalicky V,Jezbera J,et al.Limnohabitans australis sp. nov. isolated from a freshwater pond,and emended description of the genus Limnohabitans[J].International Journal of Systematic and Evolutionary Microbiology,2010,60(12):2946-2950.
[19] Ge H,Batstone D J,Keller J.Biological phosphorus removal from abattoirWastewater at very short sludge ages mediated by novel PAO clade Comamonadaceae[J].Water Research,2015,69:173-182.
[20] Kim S,Lee W,Song I,et al.Production of uracil from methane by a newly isolated Methylomonas sp. SW1[J].Journal of Biotechnology,2016,240:43-47.
[21] Vorobev A,Beck D,Kalyuzhnaya M G,et al.Comparative transcriptomics in three Methylophilaceae species uncover different strategies for environmental adaptation[J].Peer J,2013,1(1):e115.
[22] Han C,Kotsyurbenko O,Chertkov O,et al.Complete genome sequence of the sulfur compounds oxidizing chemolithoautotroph Sulfuricurvum kujiense type strain(YK-1T)[J].Standards in Genomic Sciences,2012,6(1):94-103.
[23] Jinwook C,Sunghoon S,Jeill O.Characterization of a microbial community capable of reducing perchlorate and nitrate in high salinity[J].Biotechnology Letters,2009,31:959-966.
[24] Medvedkova K A,Khmelenina V N,Suzina N E,et al.Antioxidant systems of moderately thermophilic methanotrophs Methylocaldum szegediense and Methylococcus capsulatus[J].Microbiology,2009,78(6):670-677.
Bacterial diversity in rhizosphere water ofin degenerating wetland of Wuchang Lake
LIU Zhaowen1,WANG Yu2,YANG Limin1,YE Shan1,CHENG Qihang1
(1. School of Materials and Environmental Engineering,2. School of Geography and Planning,Chizhou University,Chizhou 247000,China)
In order to study the relationship between wetland bacterial community structure and function and degraded wetland environment,selects the rhizosphere water environment samples ofin Wuchang Lake wetland,extracts the bacterial genome DNA,and uses 16S rDNA gene for 454 high-throughput sequencing.A total of 17 051 high-quality sequences and 3 032 OUT were obtained.The results showed that the bacterial community structure ofrhizosphere water in the degraded wetland of Wuchang Lake involved 15 phyla and 50 genera,among which Proteobacteria,Bacteroides,Actinobacteria and Cyanobacteria were the main phyla.There are obvious differences in bacterial community structure in different water environments in the rhizosphere of.The variation characteristics of biodiversity indexes Shannon,Chao,ACE index and OTUs all show that the intervention ofleads to the decrease of bacterial community diversity in the rhizosphere of.There is a close relationship between species composition and environmental function in wetland microenvironment.Exploring the structural characteristics of rhizosphere bacterial community ofin the process of wetland degradation is of great significance to reveal the mechanism of wetland degradation in shallow lakes in the middle and lower reaches of the Yangtze River.
wetland;;bacterial diversity;high throughput sequencing;Wuchang Lake
1007-9831(2022)10-0059-08
X826
A
10.3969/j.issn.1007-9831.2022.10.012
2022-05-31
池州學院校級重點項目(CZ2019ZRZ02,CZ2021ZRZ12);大學生創新創業項目(202011306157);材料加工實驗實訓中心項目(2021XSXZX01);材料與化工一流本科人才示范引領基地項目(2020rcsfjd28)
劉趙文(1992-),男,安徽銅陵人,助理實驗師,碩士,從事環境污染微生物防治研究.E-mail:liuzhaowen92@163.com
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
