艱難類梭菌耐藥機制及其適應性代價的研究進展
2022-11-11 12:03:54劉曉璇王偉剛趙建宏
中國抗生素雜志 2022年9期
關鍵詞:耐藥
劉曉璇 王偉剛 趙建宏
(河北醫科大學第二醫院 河北省臨床檢驗中心,石家莊 050000)
艱難類梭菌(Clostridioides difficile,C.difficile)是一種革蘭陽性、產芽胞的專性厭氧菌,是抗生素相關性腹瀉最常見的病原菌[1]。2016年,Lawson等[2]將原屬于梭菌科、梭菌屬(Clostridium)的艱難梭菌重新歸類為消化鏈球菌科、類梭菌屬(Clostridiodes)。隨后,ICSP和CLSI相繼發布公告,將艱難梭菌更名為艱難類梭菌[3]。
艱難類梭菌感染(Clostridioides difficileinfection,CDI)患者的臨床癥狀可由自限性腹瀉發展為結腸炎、中毒性巨結腸,甚至死亡[1]。2000年,獲得氟喹諾酮耐藥性的高產毒株RT027型艱難類梭菌在歐美地區暴發流行,CDI發病率和死亡率迅速升高,引起世界范圍的廣泛關注[4]。近年來,隨著抗菌藥物的廣泛應用,艱難類梭菌的耐藥率不斷攀升[5-6]。臨床分離菌株對常規治療藥物,如:甲硝唑、萬古霉素和非達霉素等敏感性降低的現象時有發生,CDI治療無效率及復發率逐年遞增,給全球公共健康帶來了嚴重威脅[7-8]。通常,細菌獲得耐藥性后易產生適應性代價(fitness cost),即生長傳播速度、定植能力、毒力等降低[9]。當環境中缺乏抗菌藥物選擇壓力時,敏感菌株便呈現生長優勢,耐藥菌株可逐漸被清除[10]。然而多數耐藥艱難類梭菌缺乏適應性代價,這極大地增加了耐藥菌株的傳播風險[11-12]。因此,本文對艱難類梭菌的耐藥機制及其適應性代價進行綜述,旨在為減少耐藥性、開發新療法提供理論參考。
1 可誘發CDI的抗生素
濫用廣譜抗生素是CDI的主要危險因素。正常情況下,腸道共生菌能通過定植占位或競爭營養等機制,阻止病原菌在腸道定植。長期使用抗生素會導致菌群失調,艱難類梭菌產毒菌株過度增殖,引發CDI[13]。幾乎所有抗菌藥物都與CDI發生相關,尤其克林霉素、頭孢菌素、氟喹諾酮以及四環素具有更高的誘發風險[4,14-16]。
1.1 大環內酯類酰胺鏈霉素
大環內酯類酰胺鏈霉素B(macrolide-lincosamidestreptogramin B,MLSB)與細菌核糖體的50S亞基結合,干擾蛋白質的合成。MLSB家族中,與CDI密切相關的是克林霉素和紅霉素。克林霉素是第一個被證實與CDI發生相關的抗生素,它的濫用引起了美國早期CDI的大面積暴發[15]。美國縱向監測結果顯示,近年來艱難類梭菌對克林霉素的耐藥率為30.1%~56.8%,呈現上升趨勢[17]。我國各地區的克林霉素耐藥率普遍較高,介于53.7%~94.8%之間,北京耐藥率高達94.8%,見表1。
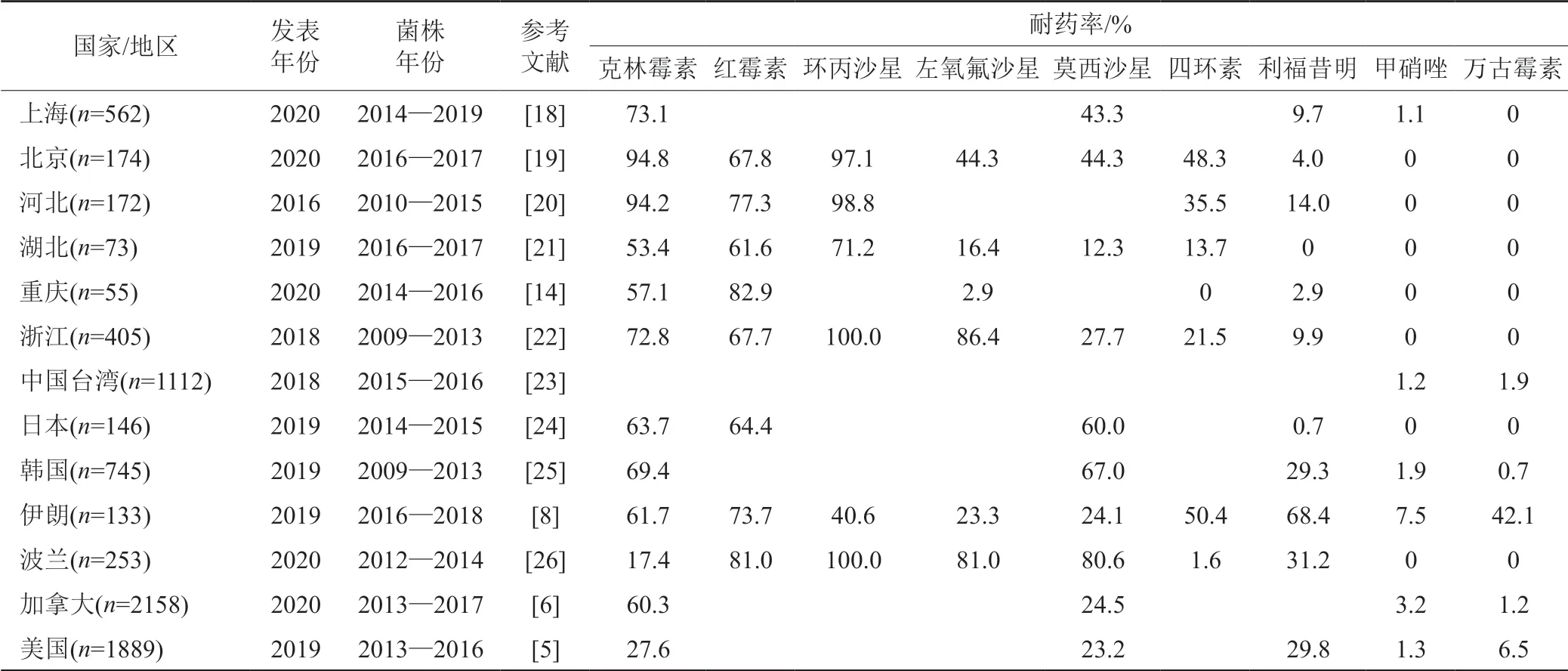
表1 不同國家/地區艱難類梭菌臨床分離株的耐藥率Tab.1 The antimicrobial resistance patterns of C.difficile isolates at different sites
紅霉素和克林霉素的耐藥機制相似,erm基因是艱難類梭菌產生耐藥的首要原因,其次是cfr基因[27-28]。二者各自編碼位于不同可移動遺傳元件的23S rRNA甲基轉移酶(前者由轉座子Tn6194、Tn6215、Tn5398和Tn5398樣攜帶;后者常由轉座子Tn916樣攜帶),通過基因水平轉移的方式傳播[29-30]。erm基因有多種亞型,最常見的是ermB,可導致艱難類梭菌對紅霉素、克林霉素高水平耐藥(MIC>256mg/L)[27]。雖然攜帶ermB基因的可移動元件會使艱難類梭菌產生適應性代價,但在傳播能力和艱難類梭菌固有遺傳特性等多種因素的共同作用下,這些遺傳元件仍然能夠在艱難類梭菌中廣泛存在[29]。
1.2 β-內酰胺類抗生素
β-內酰胺類抗生素是一類具有β-內酰胺環的抗菌藥物,通過抑制細菌細胞壁合成發揮抗菌作用,主要包括頭孢菌素、青霉素以及碳青霉素烯類抗生素[31]。艱難類梭菌對β-內酰胺類抗生素耐藥的機制主要包括產生抗生素降解酶(β-內酰胺酶)和/或產生抗生素靶點保護蛋白(青霉素結合蛋白)[32]。
頭孢菌素耐藥是20世紀80年代醫院獲得性CDI發病率上升的關鍵因素。近年來,有學者提出頭孢菌素可以抑制芽胞形成,與萬古霉素聯合用藥可預防CDI復發[33]。但頭孢菌素對腸道正常菌群有明顯的抑制作用,是CDI發生的危險因素,因此該提議存在較大爭議[14,34]。此外,艱難類梭菌對頭孢菌素耐藥的現象十分普遍,不同國家及地區的頭孢菌素耐藥率介于26.7%至100.0%之間。艱難類梭菌固有的D類β-內酰胺酶使其對頭孢菌素耐藥,耐藥水平的高低與啟動子的表達效率有關,部分學者認為艱難類梭菌對頭孢菌素天然耐藥[35]。
1.3 氟喹諾酮
氟喹諾酮類抗菌藥物(fluoroquinolones,FQs)作用于細菌的DNA回旋酶或/和拓撲異構酶Ⅳ,通過抑制細菌DNA合成發揮殺菌效應。“氟喹諾酮時代”來臨前,RT027型艱難類梭菌曾是北美地區一種非常罕見的克隆株,氟喹諾酮耐藥性的獲得使其在歐美地區暴發流行[4]。北京一項多中心研究顯示,當地優勢菌株RT369/ST81型艱難類梭菌對氟喹諾酮耐藥率為100.0%(46/46),顯著高于其他RT/ST型,推測該菌株的流行與氟喹諾酮耐藥有關。因此,亟需嚴格控制氟喹諾酮類抗菌藥物的使用,同時密切監控RT369/ST81型艱難類梭菌的出現,防止其大面積暴發流行[19]。
我國各地區艱難類梭菌分離株對氟喹諾酮普遍耐藥(表1)。其中,環丙沙星耐藥率最高,浙江地區分離株的耐藥率甚至高達100.0%,左氧氟沙星、莫西沙星次之,該差異可能與藥物的使用頻率有關。
艱難類梭菌沒有拓撲異構酶IV基因,DNA回旋酶亞單位GyrA或/和GyrB的突變是其對氟喹諾酮耐藥的主要原因,突變位點多發生于A亞單位67~106密碼子位點的喹諾酮耐藥決定區(quinolone-resistance determining region,QRDR)。據報道,GyrA的常見突變有D71V、D81N、T82I、T82V、F86Y、A83V、Ala118V和A118T,其中以T82I最為常見;GyrB的常見突變有S366A,R377G、S416A、D426N、D426V、R447K、E466V,其中以D426N和D426V最為常見[21,26,36-37]。然而,Lachowicz等[26]在氟喹諾酮敏感菌株中發現也了S366A、S416A、D426N和D426V,因此這些突變可能在氟喹諾酮耐藥性中并未發揮關鍵作用。此外,ABC轉運蛋白(ATP-binding cassette transporter,ABC)編碼基因cd2068的過表達可發揮外排泵作用,導致艱難類梭菌對多種抗菌藥物,如:氨芐西林、頭孢西丁、萬古霉素、左氧氟沙星和環丙沙星的敏感性降低,產生多重耐藥性[38]。
1.4 四環素
四環素(tetracycline,TET)通過特異性結合細菌核糖體30S亞基的A位,抑制細菌合成蛋白質[31]。四環素的選擇壓力是RT078型艱難類梭菌(可引起人畜共患病的流行株)通過食物鏈迅速引發國際傳播的關鍵因素[16]。目前我國已陸續發現該型菌株,耐藥率為20.7%~75.9%[23]。
艱難類梭菌對四環素的耐藥機制主要與tet基因有關。該基因簇有多種亞型,分別通過不同的機制介導細菌對四環素的耐藥[16,39]:①TET特異性外排蛋白(Tet40、TetA(P));②核糖體保護蛋白(TetM、TetW、TetB(P)、TetO)。其中TetM是艱難類梭菌對四環素耐藥的最常見原因,其次是TetW。在產氣莢膜梭菌中,tetA(P)和tetB(P)通常由質粒pCW3攜帶,而在艱難類梭菌中,該基因則位于基因組DNA[40]。其余基因由多個轉座子,如:Tn5397、Tn916、Tn916-like和Tn6164)攜帶,可在艱難類梭菌間或艱難類梭菌與枯草芽胞桿菌、腸球菌間水平傳播[41-42]。tetX編碼黃素依賴性單加氧酶,該酶可使包括替加環素在內所有的四環素修飾失活。近期He等[43]通過數據庫挖掘和回顧性篩選分析發現攜帶TetX3或TetX4的質粒在大腸埃希菌臨床菌株中廣泛存在。艱難類梭菌尚未見該基因的相關報道。
2 常用的CDI治療藥物
近30年來,甲硝唑和萬古霉素一直是治療CDI的主要藥物,而新型窄譜抗生素——非達霉素(fidaxomicin,FDX)的出現改變了這一局面。2017年,美國臨床實踐指南將萬古霉素和非達霉素作為CDI的一線治療藥物[1]。甲硝唑因其療效遜于萬古霉素且存在神經毒性,不再推薦使用[44]。但對于輕度CDI患者,甲硝唑的治療效果與萬古霉素無差異,仍可將其作為治療首次CDI的備選方案。此外,非達霉素在許多國家(包括中國)尚未獲得監管部門的批準,因此并未得到廣泛應用,利福昔明作為其替代方案,可用于治療復發性艱難類梭菌感染(recurrenceClostridioides difficileinfection,rCDI)[1]。
2.1 萬古霉素
萬古霉素(vancomycin,VAN)是一種糖肽抗生素,作用于脂質Ⅱ的D-丙氨酸-D-丙氨酸(D-Ala-DAla),可干擾細菌肽聚糖細胞壁的合成,發揮殺菌效應[31]。近年來,全球各地陸續出現艱難類梭菌對萬古霉素耐藥的相關報道(表1)。該耐藥菌在伊朗發生大規模流行暴發,重災區耐藥率可達42.1%,我國目前僅在臺灣地區發現對萬古霉素耐藥的艱難類梭菌。
νan基因簇是常見的萬古霉素耐藥機制[45]。該基因簇編碼合成D-丙氨酸-D-乳酸(D-Ala-D-Lac)或D-丙氨酸-D-絲氨酸(D-Ala-D-Ser)的連接酶,可取代D-Ala-D-Ala,使萬古霉素喪失作用靶點。其中部分νan基因簇(νanA,νanB,νanD,νanG,νanL,νanM)的表達受調控基因νanR和νanS的控制。當環境中存在萬古霉素時,位于膜上的傳感器激酶VanS檢測到該信號,發生自我磷酸化,并將信號傳遞給細胞質中的調節蛋白VanR,VanR~P激活νanG的表達,繼而使細菌產生耐藥。2006年,研究人員在艱難類梭菌630中發現一個與糞腸球菌BM4518的νanG具有高度一致性的類νanG基因(νanGCd),而后證實85%的艱難類梭菌中存在該基因[46]。與腸球菌不同,萬古霉素誘導νanSCd表達產生的D-Ala-D-Ser連接酶的濃度對敏感性降低的影響十分微弱,不足以使艱難類梭菌對萬古霉素產生耐藥性[47]。但是當VanSRCd發生氨基酸替換突變(R314L、G319D、S313F和T349I)時,其磷酸酶活性增強,使νanGCd表達上調,導致耐藥[48]。
酰胺酶(Ami1)可以降低結核分枝桿菌對靶向細胞壁的抗生素的敏感性[49]。質粒pX18-498攜帶有一個推定的酰胺酶基因pX18-498_006,其編碼蛋白與Ami1有23%的序列一致性。艱難類梭菌獲得質粒pX18-498后,對萬古霉素的敏感性降低了8倍,并且在小鼠體內實驗中表現出更強的致病力[49]。
Leeds等[50]在兩株耐藥菌中分別發現RpoC突變(D244Y);MurG/CD2725突變(P108L)、CD3659突變(E327stop)以及SdaB/CD3222缺失(A292-295)。MurG參與肽聚糖細胞壁的合成,其突變可能會導致細菌對萬古霉素耐藥。此外,萬古霉素的選擇壓力能促進艱難類梭菌形成生物膜,膜屏障的物理保護作用及膜內細菌生理狀態的改變,也可能導致艱難類梭菌對藥物敏感性降低[51]。
2.2 非達霉素
非達霉素是一種作用于σ因子的窄譜抗生素,可在轉錄的早期階段抑制細菌RNA聚合酶活性,達到殺菌效果[31]。相比于萬古霉素和甲硝唑,非達霉素對腸道菌群作用溫和,同時還能夠破壞艱難類梭菌的生物膜,抑制艱難類梭菌產芽胞、產毒,降低CDI復發率,是當前治療rCDI的首選藥物[52-53]。艱難類梭菌對非達霉素耐藥的現象十分罕見,但敏感性降低的現象時有發生,為抗生素治療埋下了隱患[9]。
艱難類梭菌在體外對非達霉素敏感性降低主要與rpoB突變相關[54]。Leeds等[50]通過體外誘導實驗獲得非達霉素耐藥突變株,并在其中檢測到rpoB或CD22120(多重耐藥相關轉錄調控因子marR的同源物)突變。隨后Kuehne[54]和Schwanbeck[9]分別利用定點突變技術證實RpoB氨基酸替換突變V1143D、V1143G、V1143F會造成艱難類梭菌對非達霉素耐藥。攜帶V1143D突變的臨床分離株Goe-91(MIC>64 mg/L),是目前非達霉素耐藥水平最高的突變菌株[9]。
2.3 甲硝唑
甲硝唑(metronidazole,MTZ)為硝基咪唑衍生物,其硝基在無氧環境中被還原成氨基,產生細胞毒性,發揮抗厭氧菌作用。丙酮酸-鐵/黃素氧還蛋白氧化還原酶(PFOR)、鐵氧還蛋白、氫化酶等幾種參與電子傳遞的蛋白質在甲硝唑激活過程中發揮重要作用[55]。甲硝唑治療CDI失敗與甲硝唑異質性耐藥有關[7]。該耐藥性不穩定,具有介質依賴性,傳代或低溫貯藏會導致耐藥性喪失[56]。最近的兩項研究表明,血紅素對甲硝唑中介艱難類梭菌的鑒定至關重要,在培養基中添加新鮮血紅素能使具有介質依賴性的甲硝唑耐藥性保持穩定[57-58]。此外,甲硝唑藥代動力學不足,口服后未能在腸腔達到有效抑菌濃度也會導致治療失敗,并促進耐藥性的發展[55]。不同國家/地區艱難類梭菌對甲硝唑的耐藥率差異較大,介于0~7.5%(表1)。甲硝唑耐藥艱難類梭菌在我國較為罕見。
艱難梭菌對甲硝唑耐藥的機制十分復雜,可能由多因素共同作用,主要涉及硝基還原酶、鐵代謝、DNA修復能力、電子傳遞鏈和能量代謝的改變[56,59-60]。研究表明,5-硝基咪唑還原酶Nim、鐵攝取調節劑基因(fur)突變或過表達、DNA修復蛋白RecA過表達、氧化還原過程和電子轉移鏈的破壞可造成厭氧菌對甲硝唑耐藥[56,59]。Lynch[56]和Chong[59]分別通過全基因組測序及蛋白質組學分析,發現耐甲硝唑的艱難類梭菌除上述改變外還存厭氧糞卟啉氧化酶基因(hemN)和及硫胺素生物合成相關基因(thH)突變。其中Nim作為艱難類梭菌對甲硝唑耐藥的機制尚存爭議。雖然耐甲硝唑的艱難類梭菌中有47.5%存在nimA[61],但是在敏感菌株中也發現了該基因[62]。此外,伊朗某醫院81.5%(53/65)的艱難類梭菌對甲硝唑高水平耐藥(MIC≥256 mg/L),但均未檢測到nim[41]。因此,nim并非甲硝唑耐藥的決定因素,高水平的Nim可以降低艱難類梭菌對甲硝唑的敏感性,但無法使其產生高水平耐藥[55,63]。
鐵代謝與氧化還原反應酶之間的上位效應導致艱難類梭菌對甲硝唑耐藥。Deshpande等[60]通過敲除DNA錯配修復基因,構建了一株具有高突變率的艱難類梭菌,以此研究甲硝唑耐藥性在體外的演變過程。研究結果顯示,鐵轉運體FeoB1最先發生突變,接著PFOR、黃嘌呤脫氫酶(Xdh)、鐵硫調節劑(IscR)相繼突變,MIC從最初的0.25 mg/L增加至32 mg/L[60]。Xdh和IscR突變雖然突變無法單獨介導甲硝唑耐藥性,但其與PFOR和FeoB1突變之間可發揮協同作用,進一步提高艱難類梭菌的耐藥水平[60]。
Boekhoud等[63]在耐甲硝唑的艱難類梭菌(4.1%,23/563)中,發現一個可水平傳播的質粒pCDMETRO,該質粒介導了艱難梭菌對甲硝唑耐藥,可能與甲硝唑治療失敗有關。隨后,Arcay等[64]在對甲硝唑高水平穩定耐藥(MIC>256mg/L)的艱難類梭菌中,檢測到上述質粒以及HemN(E317K)和NimA(Y130S)點突變,并發現該質粒在甲硝唑耐藥中發揮主導作用。然而,目前尚不清楚pCDMETRO(包含8個開放閱讀框,分別編碼與代謝相關的蛋白)中哪些基因是造成耐藥的直接原因,需繼續深入研究。此外,與萬古霉素類似,生物膜也可以降低艱難梭菌對甲硝唑的敏感性,促使CDI治療失敗或復發[65-66]。
2.4 利福霉素
利福霉素(rifamycins,RFs)能夠特異性結合rpoB基因編碼的RNA聚合酶β亞基,抑制細菌增殖。用于治療CDI的利福霉素主要是利福平和利福昔明,尤其利福昔明口服不易吸收,可在腸腔達到高濃度,與萬古霉素聯合用藥能有效防治rCDI[1]。利福昔明與利福平存在交叉耐藥,艱難類梭菌對二者的敏感性存在相關性,利福平的藥敏結果能有效預測艱難類梭菌對利福昔明的耐藥性[67]。
利福霉素耐藥性多見于RT017型艱難類梭菌,該型別為亞太地區的流行型菌株[20,68]。在美國,臨床上約有29.8%(563/1889)的艱難類梭菌對利福霉素耐藥[5]。2017—2018年石家莊市的耐利福昔明艱難類梭菌的檢出率為19.0%(38/200),較過去5年增長了5個百分點[20,69]。我國各地區艱難類梭菌對利福霉素的耐藥率在0~19.0%之間[20-21,69],整體耐藥率略高于亞太地區平均水平(6.7%)[70]。
RpoB突變是艱難類梭菌對利福霉素產生耐藥的主要原因。如圖1所示,此類突變多集中于507-533密碼子位點的81bp核心區域,即利福平耐藥決定區(rifamycin resistance-determining region,RRDR),其中以R505K最為常見。
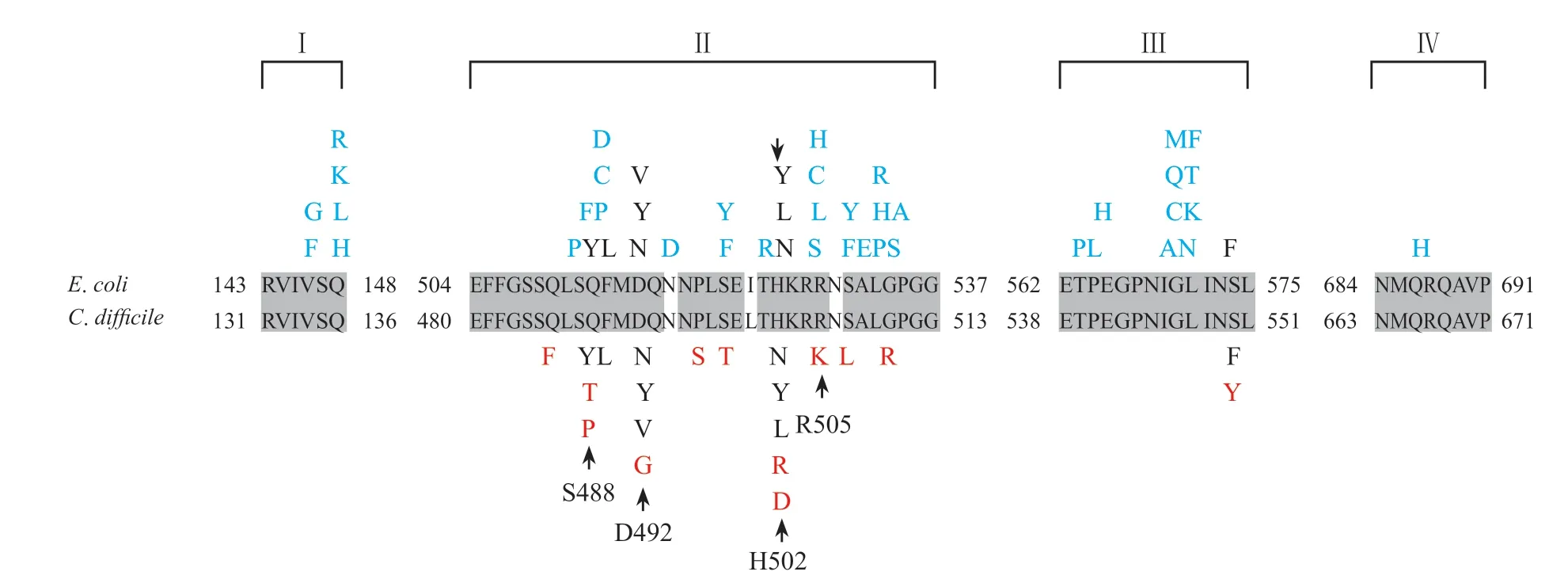
圖1 大腸埃希菌與艱難類梭菌的利福平耐藥決定區序列比對Fig.1 Sequence alignment spanning RIF resistance-determining regions(RRDRs)of the E.coli and C.difficile RpoB of RNAP
此外,臨床分離耐藥菌株還涉及H502N&R505K、H502D&D590V、H502Y&L487F、R505K&I548M、R505K&S498T、R505K&D492N等雙替換突變[20,72-73,76]。此類突變未提高艱難類梭菌的耐藥水平,推測其可能與細菌的適應性有關,即聯合突變菌株的適應性高于單獨突變。研究人員在耐藥決定區之外陸續發現新的突變,如E573A、L584F、E603N,但其與耐藥性的關聯仍需進一步驗證[11,36]。
3 耐藥艱難類梭菌的適應性代價
細菌耐藥的主要機制包括:獲得攜帶耐藥基因的移動遺傳元件,染色體基因突變。尤其細菌的關鍵基因發生耐藥突變后會影響其正常的生理功能,即產生適應性代價(fitness cost)。主要體現在突變菌株的生長傳播速度、定植及產毒能力降低[9]。當環境中缺乏抗菌藥物選擇壓力時,耐藥菌株的生存能力下降,敏感菌株便呈現生長優勢,耐藥菌株可逐漸被清除[10]。因此,適應性代價的高低在一定程度上決定了耐藥菌的傳播能力。當耐藥突變株具有較高的適應性代價時,可在其發生補償性進化(compensatory evolution)前,通過停藥控制耐藥菌株的傳播,消除耐藥性。然而多數耐藥艱難類梭菌缺乏適應性代價(表2),極大地增加了耐藥菌株的傳播風險和治療難度。

表2 艱難類梭菌染色體耐藥突變的適應性代價Tab.2 The fitness cost of antimicrobial resistance C.difficile with target-altering chromosomal mutations
DNA回旋酶是一種Ⅱ型拓撲異構酶,由GyrA、GyrB兩個亞單位組成,在基因復制和轉錄過程中發揮重要作用。GyrA和/或GyrB的氨基酸替換突變可使艱難類梭菌對氟喹諾酮耐藥。2000年,氟喹諾酮耐藥性的獲得使RT027型艱難類梭菌在歐美地區暴發流行[15]。2007年,英國限制使用氟喹諾酮類藥物以期降低CDI發病率。該措施的實施取得顯著成效,在隨后的幾年里CDI發病率明顯降低[77]。然而,艱難類梭菌目前已進化出GyrA T82I突變型,該突變可使其對氟喹諾酮高水平耐藥但不會產生適應性代價,即便環境中沒有抗菌藥物選擇壓力,艱難類梭菌依然可以維持耐藥性狀[12]。此外,GyrB分別發生D426N和D426V突變的耐藥菌株的適應性有所提高,但相較于GyrA突變該突變型在臨床并不常見[37]。
rpoB基因編碼細菌RNA聚合酶β亞基,是高度保守的單拷貝基因。該基因的關鍵位點突變會賦予細菌利福霉素或非達霉素耐藥性,同時rpoB突變會降低細菌的轉錄效率,使其產生適應性代價[78]。為提高適應性,耐藥菌株通常會伴隨rpoA、rpoB或rpoC的補償性突變[79]。然而Dang等[11]發現,在rpoB突變的耐利福昔明艱難類梭菌中,包括最常見的R505K突變株,大部分突變株缺乏適應性代價。由于該研究是通過體外誘導實驗獲得耐藥菌株,并且僅檢測利福霉素作用靶點RpoB的突變情況,不排除耐藥菌在連續傳代過程中發生補償性突變的可能。并且,在該研究的體外競爭實驗中,適應性分別降低20%和33%的D492Y和S550Y突變菌株在小鼠體內并未表現出適應性代價,研究人員認為可能是由于黃金地鼠對艱難類梭菌的易感染性使得耐藥菌株的適應性代價的細微差異無法在體內環境中得到體現[11]。
Kuehne等[54]利用定點突變技術構建了對非達霉素耐藥的RpoB突變株(V1143D、V1143G、V1143F)并通過體內、外的競爭實驗分別證實這些突變給艱難類梭菌造成適應性代價,使其生長傳播速度減慢、芽胞生成和產毒能力降低,這可能是臨床環境中非達霉素耐藥株少見的原因之一。然而,同樣具有RpoB V1143D突變的臨床分離株Goe-91并未出現生長速度減緩的現象,僅毒力和生成芽胞的能力降低,適應性代價低于預期[9]。造成這一現象的原因可能是艱難類梭菌的遺傳背景彌補了突變引起的生長減緩,亦或是在小鼠體內增殖過程中發生了補償性突變,但具體機制仍需進一步研究。
Leónidas等[80]發現具有鏈霉素(RpsL K43T),利福平(RpoB H526Y)耐藥突變以及雙取代突變(RpsL K43T&RpoB H526Y)的大腸埃希菌在無菌小鼠中均具有較高的適應性代價,而在單獨飼養的SPF小鼠中,由于每只小鼠的腸道菌群具有個體差異,耐藥菌株表現出不同程度的適應性代價,甚至出現適應性提高的現象。因此,體外競爭實驗無法代替動物實驗,而在哺乳動物復雜的腸道微生態中,耐藥菌的適應性代價并未研究透徹,仍需深入研究。
4 總結與展望
綜上所述,艱難類梭菌的耐藥率在不同國家及地區之間存在較大差異(表1)。總體來說,艱難類梭菌對克林霉素、氟喹諾酮、頭孢菌素普遍耐藥,并且這些抗菌藥物具有更高的誘發CDI的風險,臨床使用時應警惕腹瀉的發生,一旦出現相關癥狀應立即停用。對于CDI一線治療藥物(甲硝唑、萬古霉素和非達霉素等),絕大多數菌株敏感性較高,部分菌株出現敏感性降低的現象應引起高度重視。對此,我國亟需在全國范圍內進行大規模的CDI流行病學調查以及艱難類梭菌耐藥檢測;同時嚴格規范抗菌藥物的使用,預防多重耐藥菌的出現及流行。艱難類梭菌的耐藥機制十分復雜,目前的研究雖已取得一些新的進展,但仍有許多耐藥機制至今尚未明確,需進一步研究。此外,耐藥突變產生的適應性代價較低也是耐藥菌株暴發流行的原因之一。對此,應當對低適應性代價的機制進行深入研究,設計降低細菌適應性、抑制耐藥性進化的抗菌藥物,與一線治療藥物聯合應用,增加細菌對藥物的敏感性,延長藥物的使用壽命。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
