鰱微衛星標記與生長性狀的相關分析
2022-11-23 07:32:16羅相忠李曉暉梁宏偉鄒桂偉
淡水漁業 2022年6期
關鍵詞:生長
羅相忠,時 樂,沙 航,李曉暉,梁宏偉,鄒桂偉,崔 峰
(1.中國水產科學研究院長江水產研究所,武漢 430223;2.安徽科技學院生命與健康科學學院,安徽鳳陽 233100)
微衛星DNA是廣泛存在于真核生物和原核生物基因組中的1-6個簡單重復序列[1]。微衛星標記在基因組中不同區域分布差異較大,大多數富集在非編碼區[2]。由于微衛星遺傳標記具有高度多態性、信息含量豐富、共顯性等優勢,被廣泛用于生物物種數量性狀QTL定位、遺傳連鎖圖譜構建和DNA指紋分析[3]。微衛星標記已經發展成為水產動物育種領域中的常用分子標記之一。
微衛星標記已經在眾多水產動物中開展了與生長等性狀的關聯分析,取得了良好的結果。如李燕等[4]通過分析15個微衛星標記與96尾雜交鱖(Sinipercachuatsi×S.scherzeri)的體長、體質量、體高性狀的相關性,獲得cD90、MDJ821、PY21、PY55和72217標記分別與上述3種生長性狀顯著相關。余成晨等[5]研究選育草魚(Ctenopharyngodonidella)群體生長性狀與微衛星標記的關系,發現CID1512、CID973_1 和CID254_1 在雌性或雄性個體中與體質量和體長等生長性狀顯著相關,而CID391_2 僅在雄性個體中顯著相關。劉士力等[6]對翹嘴鲌(Erythroculterilishaeformis)的2個生長激素受體GHR1 和GHR2基因的微衛星序列進行檢測,發現GHR2基因中的4個微衛星位點均與體長和體質量等生長性狀相關。這些為開展鰱(Hypophthalmichthysmolitrix)微衛星標記遺傳多樣性與生長性狀的相關分析提供了借鑒。
鰱屬于鯉形目鯉科鰱亞科鰱屬,是我國重要養殖魚類,為淡水養殖四大家魚之一,其營養豐富,為人類的動物蛋白來源之一[7]。同時其以水體中浮游植物為主要餌料,屬于淡水魚類中為數不多的不投餌的濾食性魚類,為典型的碳匯漁業對象,具有重要的生態價值[8]。鰱食物鏈短,易于養殖,深受養殖者和大眾消費者青睞,在我國養殖產量僅次于草魚。長豐鰱(H.molitrix)是中國水產科學研究院長江水產研究所利用綜合育種技術培育的鰱新品種,具有生長速度快、體型好和適應性強等優點[9]。基于研究鰱的遺傳多樣性以及篩選與其生長性狀相關的微衛星標記,本研究以湘江鰱(H.molitrix)和新品種長豐鰱作為研究對象,旨在篩選與鰱生長性狀相關的微衛星分子標記,為今后分子標記輔助育種提供技術支撐。
1 材料和方法
1.1 樣本采集
實驗用2齡長豐鰱和2齡湘江鰱均采自于農業農村部鰱遺傳育種中心,其中長豐鰱60尾,湘江鰱120尾。采集其胸鰭組織,置于無水乙醇的離心管中,帶回實驗室于-20 ℃冰箱中保存。
1.2 生長性狀測定
對長豐鰱和湘江鰱依次進行框架數據X1、X2、X3、X4、X5、X6、X7、X8的測量(圖1和表1)和全長(X9)、體長(X10)、頭長(X11)、體高(X12)、尾柄高(X13)、眼徑長(X14)、尾柄長(X15)、軀干長(X16)的測量,測量工具分別為直尺和游標卡尺。體質量(X17)使用電子天平測定。共計17個可量性狀。采用SPSS 22.0軟件(由美國SPSS公司開發)進行單個樣本的Kolmogorov-Smirnov(K-S),檢驗所取樣本的生長性狀是否符合正態分布(P>0.05)。
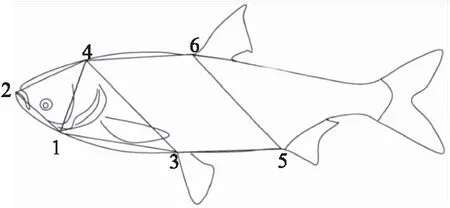
圖1 鰱框架結構示意圖

表1 鰱形態測量表
1.3 基因組DNA的提取
鰭條經超純水漂洗后,剪取少量鰭條組織,采用高鹽法提取樣本的基因組DNA。提取后,將DNA溶液經1%瓊脂糖凝膠電泳和紫外凝膠成像系統檢測后,加入超純水稀釋至50 ng/μL,-20 ℃冰箱中保存備用。
1.4 微衛星引物的篩選
基于本實驗室鰱基因組數據(未發表),采用Primer5.0(加拿大Primer生物公司)設計96對微衛星引物,其序列和退火溫度見表2,引物由武漢天一輝遠生物科技有限公司合成。選用湘江鰱和長豐鰱各3尾對所有引物進行初步篩選,用1.0%的瓊脂糖凝膠電泳進行檢測后毛細管電泳,進而開發多態微衛星標記。

表2 鰱13個微衛星引物序列信息
1.5 PCR擴增和條帶的檢測
對基因組DNA進行PCR擴增,總反應體系為10 μL,其中2×Taq PCR Master Mix 5 μL,基因組DNA 1 μL,上下游引物各0.5 μL,ddH2O 3 μL。PCR擴增程序為:95 ℃預變性5 min;95 ℃變性,62℃退火30 s,72 ℃延伸30 s,以后每個循環下降1 ℃,直至52 ℃,共10個循環;95 ℃變性30 s,52 ℃退火30 s,72 ℃延伸30 s,共25個循環;72 ℃延伸20 min。擴增的PCR產物在ABI-3137XL基因分析儀(美國ABI生物公司)上利用毛細管電泳法(CE)檢測。每個CE樣品中含有1 μL PCR產物、0.4 μL Genescan-500分子質量標準品和20 μL去離子甲酰胺(美國ABI生物公司)。在毛細管電泳過程中,PCR產物與Genescan-500分子質量標準品的熒光信號均由基因分析儀自動保存。
1.6 數據統計及分析
利用SPSS 22.0軟件對湘江鰱和長豐鰱兩個群體的生長性狀進行數據整理和正態分布的檢驗。利用Cervus 3.0軟件(Cervus公司)統計兩群體不同位點的等位基因數(k)、期望雜合度(He)、觀測雜合度(Ho)和多態信息含量(PIC)。利用SPSS 22.0軟件的線性回歸對兩個鰱群體的不同微衛星位點與生長性狀進行關聯分析,初步確定與生長性狀存在顯著差異的位點,再用一般線性模型(GLM)對初步確定的位點進行單變量的觀測平均值的多重比較,確定同一微衛星位點不同基因型間是否存在顯著差異,P<0.05為差異顯著。為了排除偶然性,在利用SPSS 22.0軟件分析時,每種基因型至少具有3次觀察值才被統計。
2 結果與分析
2.1 長豐鰱和湘江鰱生長性狀的統計
長豐鰱中體質量的變異系數最大為15.67%,湘江鰱中體質量的變異系數最大為15.68%(表3),體質量在鰱群體中的變異系數均較大,相對于其他生長性狀,體質量具有更大的選擇潛力。湘江鰱和長豐鰱的生長性狀均服從正態分布,測量的所有性狀都具有連續變異的特點,可進行后續的關聯分析。
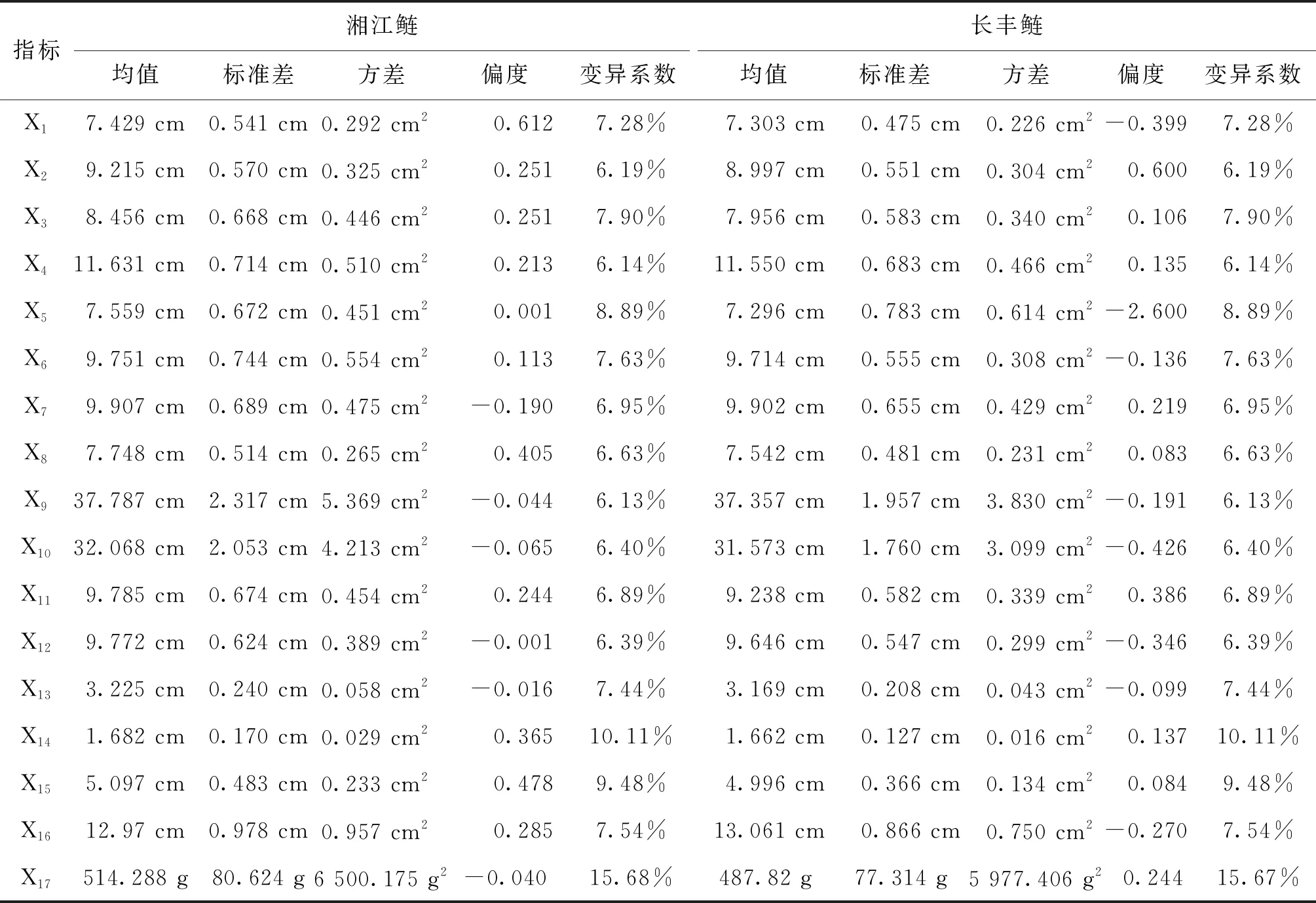
表3 長豐鰱和湘江鰱各生長性狀統計
2.2 微衛星多態性標記分析
96個微衛星位點中有13個多態性位點均能在長豐鰱和湘江鰱中穩定擴增,這13個多態性位點具體分布為:位點P058位于染色體LG1上,P030、P036位于LG2上,P086位于LG5,P034位于LG9上,P021、P062位于LG12上,P056位于LG13上, P037、P043位于LG16上,P054位于LG17上,P041位于LG23上,P020位于LG24上。長豐鰱和湘江鰱兩個群體的遺傳多樣性信息見表4。長豐鰱共檢測到56個等位基因,平均k為4.31個,Ho為0.017~0.683,He為0.049~0.800,PIC范圍為0.048~0.768。湘江鰱群體共檢測到60個等位基因,平均k為4.62個,平均Ho為0.050~0.867,平均He為0.126~0.822,PIC為0.177~0.796。
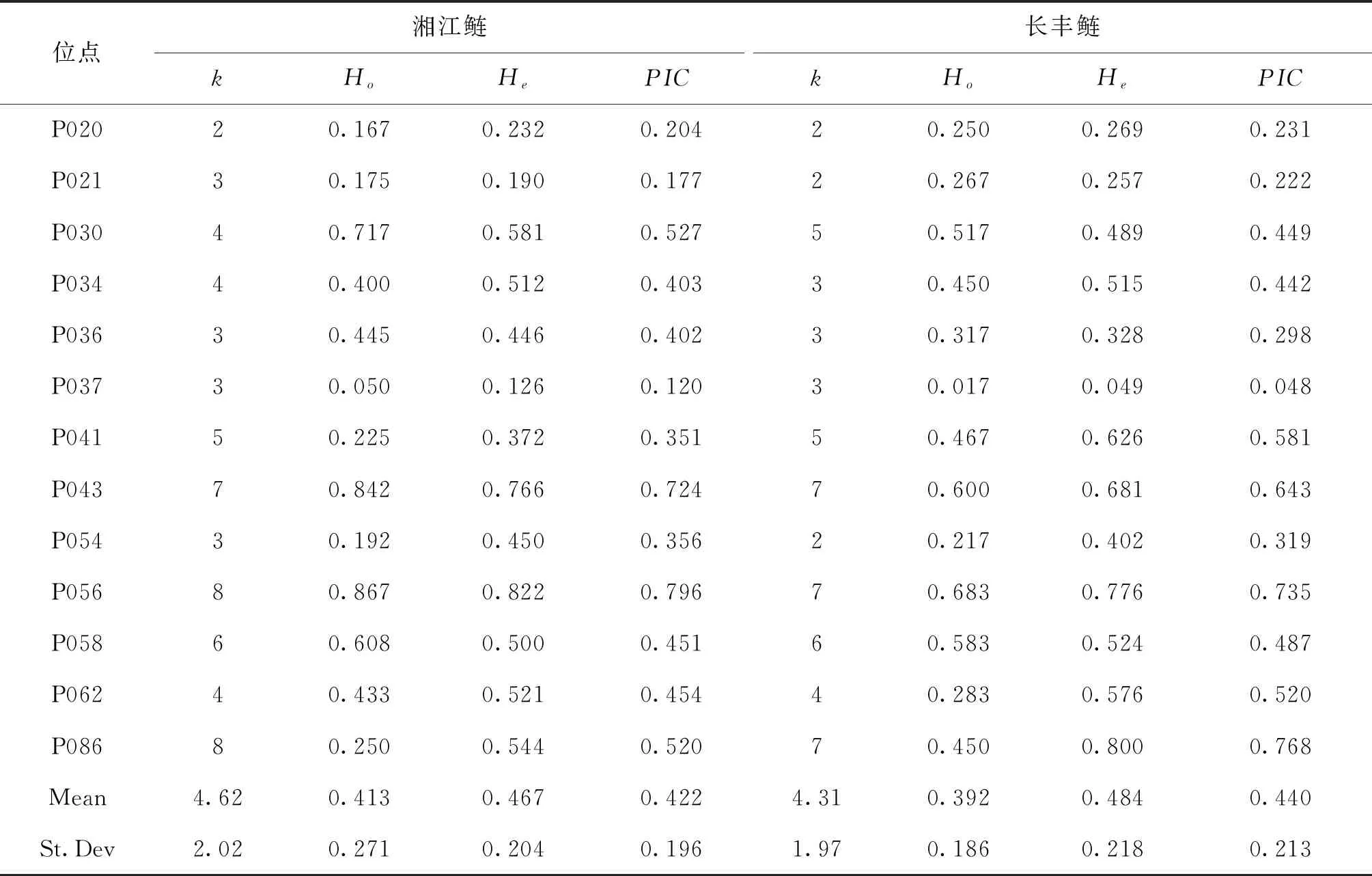
表4 鰱生長性狀有關的SSR多態位點信息
2.3 微衛星標記與鰱生長性狀的相關性分析
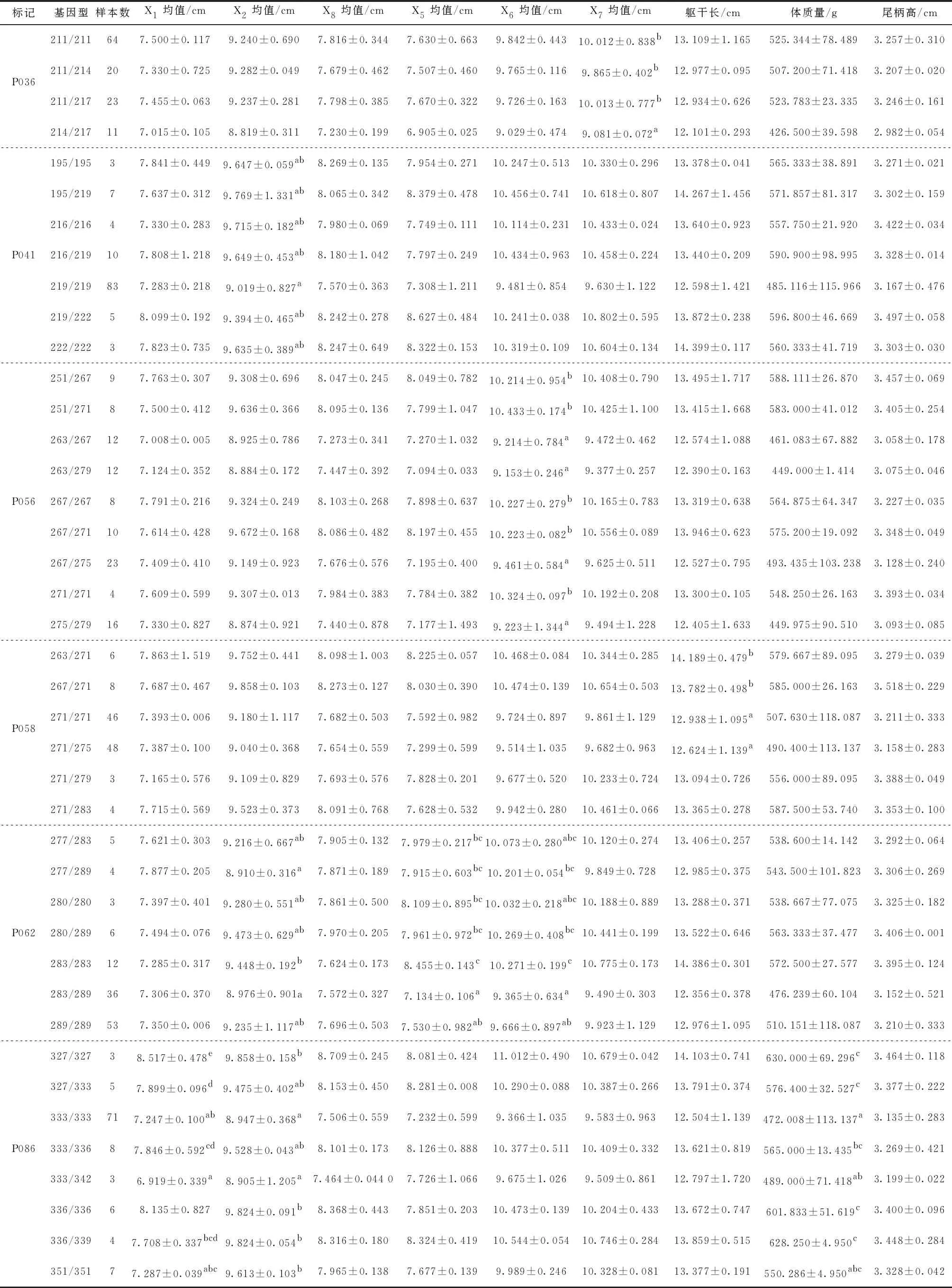
13個微衛星位點中10個微衛星位點與生長性狀顯著相關,見表5。對具有關聯性的9個生長性狀進行不同位點的均值或多重比較。其中,篩選出湘江鰱8個與生長性狀顯著相關的位點(表5),長豐鰱2個顯著相關位點(表6)。
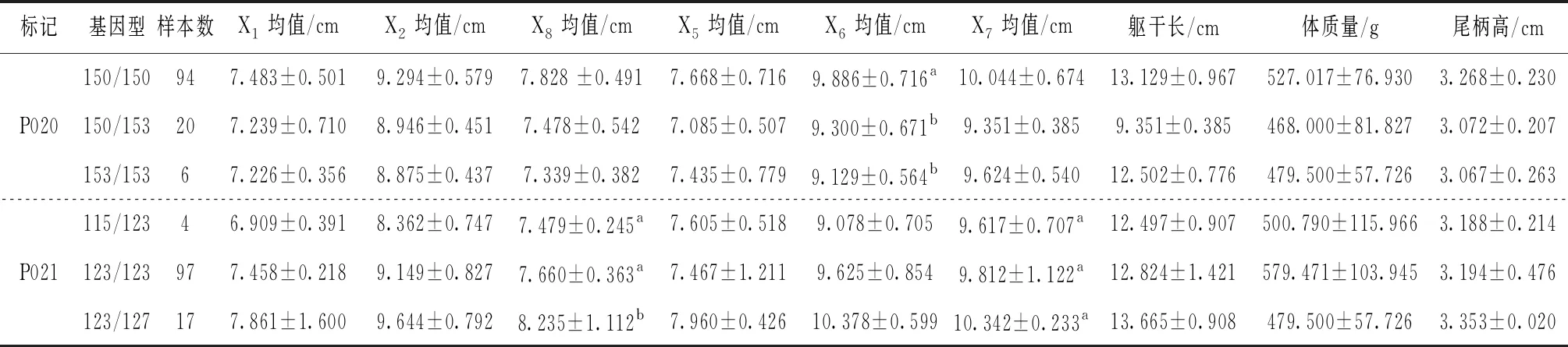
表5 湘江鰱8個微衛星位點各生長性狀的多重比較
續表5
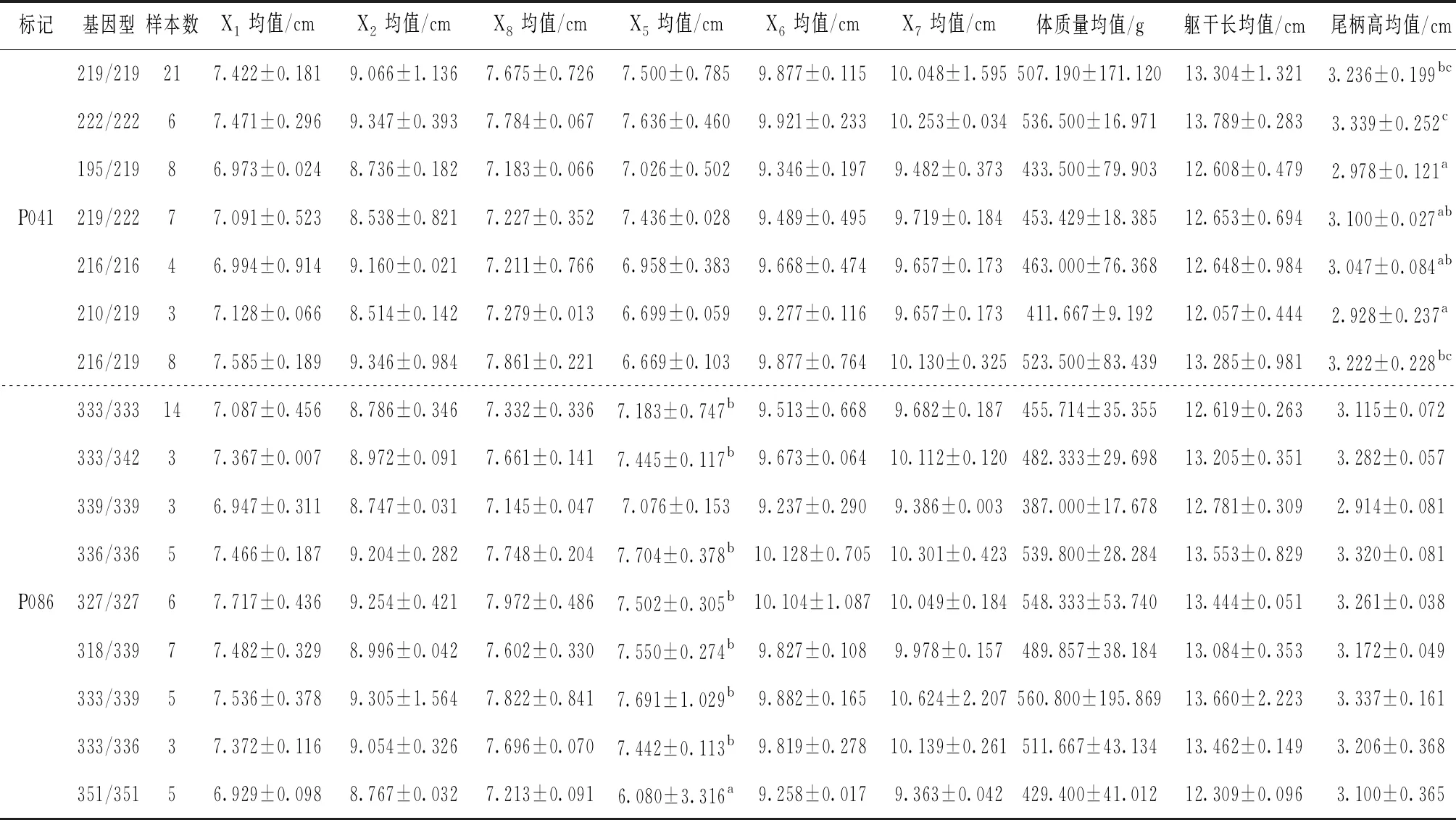
表6 長豐鰱2個微衛星位點各生長性狀的多重比較
2.3.1 微衛星位點與湘江鰱生長性狀的相關性分析
湘江鰱群體中,位點P020、P056、P062均與框架數據X6顯著相關;位點P041、P062、P086均與框架數據X2顯著相關。位點P021與框架數據X8均值顯著相關。位點P058與軀干長顯著相關。
對不同基因型的不同性狀進行多重比較。位點P020中,150/150基因型個體各生長性狀均值均高于其他基因型;個體150/150基因型的框架數據X6的均值顯著高于150/153、153/153基因型,而基因型150/153與153/153個體各生長性狀之間差異不顯著,由此得出150/150基因型是生長性狀的優勢基因型。在位點P021中,基因型123/127個體各生長性狀均值均優于其他兩種基因型,框架數據X8上123/127基因型個體顯著高于115/123、123/123基因型。位點P036,共檢測到4種基因型,基因型214/217各生長性狀均值均低于其他基因型,且框架數據X7顯著低于211/211、211/214、211/217和217/217基因型,由此證實214/217基因型是生長性狀的劣勢基因型。位點P041中,基因型219/219個體各生長性狀均值均低于其他基因型,且框架數據X2均值顯著低于其他基因型,該基因型也被證實為生長性狀的劣勢基因型。在位點P058中共檢測到6種基因型,基因型263/271、267/271個體的軀干長均值分別為14.189 cm和13.782 cm,顯著高于基因型271/271和271/275,但基因型263/271和基因型267/271的軀干長均值沒有明顯差異。位點P062,283/283基因型個體的框架數據X2、X5和X6顯著高于其他基因型,283/289框架數據X5顯著低于基因型277/283、280/289、283/283,X6顯著低于280/289、283/283基因型。位點P086中共檢測到8種基因型,個體327/327基因型的框架數據X1均值顯著高于基因型333/333、333/336/、333/342、336/339、351/351,高于基因型327/333和336/336但無顯著差異,327/327基因型為框架數據X1的優勢基因型。
2.3.2 微衛星位點與長豐鰱生長性狀的相關分析
長豐鰱群體中,位點P041與尾柄高顯著相關,位點P041共檢測到7種基因型,基因型195/219、210/219個體的尾柄高均值分別為2.978 cm和2.928 cm,均低于基因型219/219、222/222、216/219的個體,差異顯著,但基因型195/219和210/219之間無明顯差異。個體222/222基因型的尾柄高均值為3.339 cm,顯著高于基因型219/222、216/216、210/219的個體,高于其他基因型但無顯著差異。說明基因型222/222為尾柄高的優勢基因型,222 bp片段的等位基因與尾柄高呈正相關。
長豐鰱群體中,位點P086中,檢測到9種基因型,基因型351/351個體的框架數據X5均值為6.080 cm,顯著低于基因型333/339、333/342、336/336、327/327、318/339、333/339、333/336,低于339/339基因型但差異不明顯;基因型351/351為框架數據X5的劣勢基因型。
2.4 微衛星標記在鰱群體的適用性分析
2.4.1 微衛星標記P041與湘江鰱、長豐鰱生長性狀的相關分析
P041位點在兩群體中共檢測到8種基因型,湘江鰱和長豐鰱中均檢測到7種基因型,兩群體共有基因型6種,分別是基因型195/291、216/216、216/219、219/219、222/222、219/222。其中,基因型222/222個體在湘江鰱、長豐鰱中的軀干長均值分別為14.399 cm和13.789 cm,均高于其群體的其他基因型,但無顯著差異,說明222 bp片段的等位基因個體與軀干長呈正相關。
2.4.2 微衛星標記P086與湘江鰱、長豐鰱生長性狀的相關分析
P086位點在湘江鰱中檢測到8種基因型,在長豐鰱中檢測到9種基因型,其中6種基因型為兩群體共有:327/327、333/333、333/336、333/342、336/336、351/351。湘江鰱中333/333基因型個體的各生長性狀均值均低于333/342外的其他基因型個體但無顯著差異;長豐鰱中基因型333/333個體各生長性狀的均值低于其他基因型,除基因型339/339、351/351外,但無顯著差異,333 bp片段的等位基因與生長性狀呈負相關。
3 討論
3.1 長豐鰱與湘江鰱的遺傳多樣性
遺傳多樣性是物種多樣性的基礎,也是生命適應環境和進化的基礎[10]。遺傳雜合度是衡量群體遺傳變異的參數之一,種群遺傳多樣性越豐富,該物種對環境的適應能力越強[11]。本實驗中篩選得到的13個微衛星標記,在湘江鰱、長豐鰱中的平均觀測雜合度分別為0.413、0.392;在湘江鰱、長豐鰱中的平均期望雜合度分別為0.467、0.484。兩群體的雜合度都處于中度水平,遺傳多樣性處于中等多態水平。用于基因變異程度高低的多態信息含量(PIC)>0.5,則該位點是高度多態的;0.25 分子標記輔助育種,就是通過與基因連鎖的分子標記,直接篩選目標區域的DNA[13]。其可以避免環境的影響,提高選擇的準確性,因而廣泛運用于許多水產生物的遺傳連鎖圖譜構建、性狀關聯分析和改良品種的輔助育種等研究。湘江鰱群體中,位點P020、P056、P062均與框架數據X6顯著相關(P<0.05);位點P041、P062、P086均與框架數據X2顯著相關,這些位點中存在著“多因一效”的現象。同時發現,一個標記位點也可能與多個性狀密切相關,位點P062與湘江鰱的框架數據X2、X5、X6顯著相關;位點P086與湘江鰱的框架數據X1、X2和體質量顯著相關,位點中存在著“一因多效”的現象,在對其他一些水產動物群體的遺傳多樣性研究也發現有類似的現象[14-15]。 近年來,分子標記與性狀間的關聯分析已經成為當前水產動物遺傳育種的研究熱點之一。不同位點的基因型與性狀的關聯分析,可直接通過對基因型和表型變異程度的差異分析,確定標記與數量性狀之間的相關性,得到數量性狀與一個或多個標記的遺傳相關,從而篩選出控制數量性狀的基因型[16]。但是,目前國內外文獻有關鰱群體的研究主要集中在人工繁育和遺傳多樣性[9,17-18],有限的研究報道了有關鰱基因組上的QTL分析[19-20]。WANG等[21]對鰱的生長性狀與微衛星標記的研究,鑒定了20個生長性狀QTL相關標記,發現兩個標記ref-16553和ref-46998_8均與體重體長等多個生長性狀顯著相關。本研究中標記P041在湘江鰱和長豐鰱兩個群體中均存在222 bp片段的等位基因,且該位點在湘江鰱、長豐鰱中分別與框架數據X2、尾柄高有顯著相關性。同一標記在長豐鰱和湘江鰱上關聯的生長性狀不同,可能是由于長豐鰱和普通鰱的形態略有不同[22],顯著相關的位點在兩群體中所表現的性狀有所差異。兩群體在標記P086上的333/333基因型個體各生長性狀均值相對于其他基因型普遍偏低,該基因型在鰱群體中可能對生長性狀有負面影響。由于樣品量有限,本研究得到的結果尚需大群體樣本的進一步驗證。這些位點的某些基因型具有顯著的生長優勢,這些基因型可在未來的選育工作中有意識的富集[23],從而用于后續的分子標記輔助育種工作。 在水產生物育種研究中,關于鰱群體的生長性狀與標記輔助育種的基因的連鎖分析報道很少,本研究獲得的與生長性狀相關的標記能否在湘江鰱和長豐鰱的標記輔助選擇實踐中得到應用,仍需進一步深入研究與驗證,以便達到其能合理地應用該標記指導育種實踐,篩選具有優勢生長性狀的良種,提高育種速度,改良物種重要經濟性狀的目標。3.2 分子標記與性狀之間的關聯效果
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
