皺紋盤鮑(Haliotis discus hannai)不同群體貝殼形態和相對生長變異研究*
2022-11-25 05:30:30仉佳文唐君瑋張遠帆張國范吳富村
海洋與湖沼 2022年6期
仉佳文 唐君瑋 張遠帆 魏 敏 張國范 吳富村①
(1. 江蘇海洋大學江蘇省海洋生物技術重點實驗室 江蘇連云港 222005; 2. 中國科學院海洋研究所實驗海洋生物學重點實驗室山東青島 266071; 3. 長島海洋經濟促進中心 山東煙臺 265800)
皺紋盤鮑(Haliotis discus hannaiIno)自然分布于亞洲東部海域, 包括我國遼東半島和山東半島海區,以及日本北部和朝鮮半島部分海區(呂端華, 1978)。在我國, 皺紋盤鮑是重要的漁業資源, 20 世紀50 年代捕撈量一度達100 t 以上, 而由于酷捕濫采和缺乏有效的漁業管理, 我國皺紋盤鮑資源遭到了嚴重破壞(張昭, 1979), 海珍品產量逐年下降至近枯竭。隨著我國皺紋盤鮑人工繁育技術的突破(陳木等, 1977; 劉永峰等, 1984), 以人工苗種底播為主要方式開始了皺紋盤鮑資源修復工作。在20 世紀80 年代后期至 90 年代, 山東、遼寧等海區的底播鮑的采捕量得以顯著提升(劉永峰等, 1994; 吳富村等, 2020)。之后雜交技術開始在我國皺紋盤鮑人工養殖產業成功應用, 逐漸推動福建成為苗種繁育和養殖產業中心, 然而南繁苗種因較難以適應黃渤海海區環境, 其在我國北方海區難以越冬, 底播鮑的采捕產量顯著下降, 在北方部分海區甚至已放棄開展皺紋盤鮑的底播增養殖工作。近年來, 隨著我國生態環境保護政策的制定與落實, 皺紋盤鮑棲息海區如山東長島等生態環境保護力度逐年加強, 相關海區皺紋盤鮑資源密度雖仍然較低,但也呈逐步恢復態勢。皺紋盤鮑不同地理群體也是我國重要的種質資源來源, 我國已設立包括大連圓島、煙臺欽隍島等7 處皺紋盤鮑水產種質資源保護區。近幾年隨著我國關于加強農業種質資源保護與利用、開展全國農業種質資源普查的政策推動, 皺紋盤鮑不同地理群體的種質鑒定和評價工作也得到重視。
長期以來, 人們已經認識到鮑等海洋底棲動物地理種群間差異, 以及這種差異對應對過度捕撈和局部資源枯竭等資源管理的重要性(Begget al, 1999),并引入和識別管理單位(mangement units, MU)的概念,管理單位可定義為應單獨管理和監測的獨立的地理種群(Martienet al, 2003)。而如何區分地理種群或種群組成則成為管理單位成功開發和應用的基礎(Defeoet al, 2005)。識別鮑種群結構的最常用方法是分析假設種群之間的遺傳差異如基于單基因、微衛星、SNP 等分子標記的種群遺傳分析方法(Sandoval-Castilloet al, 2018; Mares-Mayagoitiaet al, 2021; Namet al, 2021), 此類研究通常顯示出比預期更大的尺度差異(Stephenset al, 2006; Sandoval-Castilloet al,2018), 如毗鄰海區小空間尺度(~10 km)可能因環境劇烈差異而造成形態變化、種群隔離, 而分子遺傳學中單基因分析無法全面反映種群遺傳分化程度, 不能區分種間雜交個體和純種個體(Liuet al, 2021), 種群基因組學則剛興起, 往往缺乏相關研究且成本較高。我國不同海區的皺紋盤鮑群體是否可以以獨立的地理種群作為管理單位, 一直以來缺乏相關數據支持。海水貝類由于貝殼形態的可塑性, 殼形態測量及變異研究在物種鑒定和種群分化中得到了廣泛的應用。形態學標記作為最直接的遺傳標記, 在鮑科動物中也廣泛的應用于鮑種間的分類和判別分析(陳世杰,1995), 如吳玉云(2001)利用鮑殼長、殼寬、殼高間的比率數值參數可基本將我國自然分布的7 種鮑科動物進行分類; 張儀方(2019)利用線性測量法分別研究不同鮑種及雜交種的形態差異, 結果顯示不同種間鮑的形態差異存在明顯區別, 可以通過形態差異區分開。除此之外, 同時張儀方(2019)對皺紋盤鮑6 個養殖群體采用形態學聚類等分析得出僅靠形態差異很難區分開所有的養殖群體的結論。而利用鮑形態學及相關測量參數間的相對生長來開展我國不同海區自然群體的變異和判別分析則未見報道。
本研究通過采集和比較我國不同海區皺紋盤鮑樣本, 利用貝殼外部形態特征和樣本重量的測量, 對皺紋盤鮑不同群體的形態和相對生長變異進行了研究。所獲得的數據對進一步了解地理群體的可塑性具有重要意義, 同時對判別種群來源提供參考。
1 材料與方法
1.1 樣本采集
在我國黃渤海自然海區位于大連的圓島(YD)、老偏島(LPD)、獐子島大蠔島(ZZD), 煙臺長島的大欽(DQ)、南隍城(NHC), 青島的靈山島(LS)以及榮成成山(XXK) 7 個地點隨機收集了700 余只皺紋盤鮑個體(圖1)。為防采樣季節對數據分析的影響, 所有樣本均來自2020 年及2021 年7 月份皺紋盤鮑資源調查時采集樣本, 潛水于2~6 m 深的潮下帶海區采集。資源調查時, 根據所屬島嶼或海區管理單位歷史記錄, 盡量選擇未采用皺紋盤鮑人工繁育苗種開展底播放流的海區, 或至少10 年內未曾開展相關底播工作的海區作為采樣點。樣本收集后, 根據殼頂顏色判別是否為人工苗種底播的個體(Gallardoet al, 2003; Marchaiset al, 2017), 如有綠殼色或者在距殼頂2~5 cm 外殼處有顯著生長殼紋, 則該個體不作為樣本分析。最終本研究共對自然海區采集的452 個皺紋盤鮑樣本進行形態學測量, 每個地點取樣的樣本數在31~153 不等。
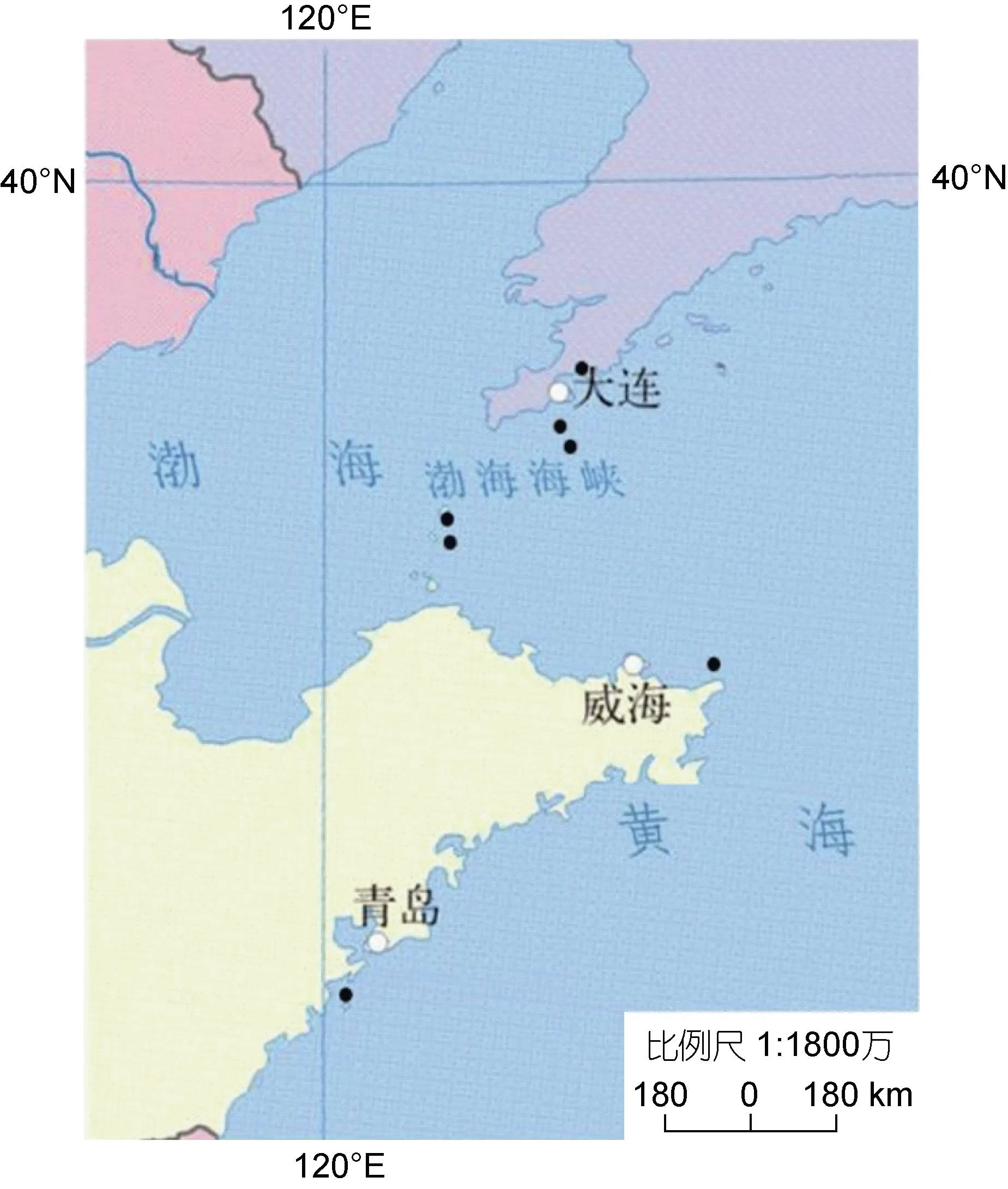
圖1 本研究沿海皺紋盤鮑采樣點分布圖Fig.1 Distribution of sampling sites for the Pacific abalone along coast of China in this study
1.2 形態測量
每個樣本采樣后立即記錄每個個體的形態特征:外殼的最大殼長(SL, 單位: mm)、殼寬(SW, 單位:mm)、殼高(SH, 單位: mm)、總濕重(TWW, 單位: g);取回實驗室解剖所有樣本, 稱量記錄軟體部重(SPW,單位: g)以及干殼重(SHELLW, 單位: g)。長度測量使用數字卡尺(精度為0.01 mm), 重量測量使用電動數字天平(精度為0.001 g), 其中干殼重SHELLW 的測定為解剖后取鮑殼暴曬72 h 后稱量。
周長、面積、彎曲度的識別和測量: 將曬干后的鮑殼樣本通過數字相機(Cannon EOS 5D4, 100 微鏡頭)拍攝以每個鮑殼樣本的圖像, 樣本拍攝記錄鏡頭與樣本距離、焦距等參數以保持一致; 然后通過軟件ImageJ (美國國立衛生研究院)計算殼口輪廓的周長(CIRC, 單位: mm)和面積(AREA, 單位: mm2); 通過Matlab 軟件計算殼口邊緣的粗糙度(CURV), 該指標定義為: (1 - 凸包周長與周長之比)×100, 其中凸包(convex hull)是一個圖形學分析中用于描述邊緣粗糙/光滑的參數, 可以用MATLAB 軟件來實現相關計算。
1.3 相對生長
通過回歸分析, 建立了所有種群和每個種群的殼形態特征(殼長SL, 殼寬SW, 殼高SH 分別與總濕重TWW、軟體部重SPW 和干殼重SHELLW)之間的異速生長關系。對各參數的自然對數變換數據采用線性最小二乘法估計相關參數a和b。lnY=lna+b×lnX(Y=aXb), (1)式中, 參數b通常用作生長比率與等距的偏差的指標(Rizhinashvili, 2008), 采用Student’st檢驗以確定異速生長的類型, 及在每組異速生長關系中, 以整合所有種群擬合得到的b值為期望, 分別以各個群體內所有個體為樣本進行Student’st檢驗。
1.4 數據處理
用所有記錄的形態測量值計算出17 個性狀比率指標(SPW/TWW、SHELLW/TWW、SHELLW/SPW、SW/SL、SH/SL、SL/CURV、SW/CURV、SH/CURV、SL/SUM、SW/SUM、SH/SUM、TWW/DSUM、SPW/DSUM、SHELLW/DSUM、SL/CIRC、SW/CIRC、SLSW/AREA), 盡管兩兩測量性狀間可獲得更多的比率指標, 本研究的17 個比率指標的選取主要依據其生物學意義, 如 SW/SL、SL/CIRC、SW/CIRC、SLSW/(100×CIRC)分別為樣本殼寬與殼長、殼長與殼口周長、殼寬與殼口周長、殼長殼寬乘積與殼口周長的比值, 可以用來表示殼型偏長或偏寬的形態;SL/SUM、SW/SUM、SH/SUM 與 TWW/DSUM、SPW/DSUM、SHELLW/DSUM 比率指標主要參考Kobayashi 等(1996)文獻, 分別表示皺紋盤鮑殼長SL、殼寬SW、殼高SH 與這三個測量指標(SL、SW 和SH)之和的比率, 以及全濕重TWW、軟體部重SPW、干殼重SHELLW 與這三個測量指標(SL、SW 和SH)之積的比率。
基于(協)方差分析(ANCOVA)對這17 個比率變量進行群體間變異顯著性比較; 進而在比率變量差異顯著下, 對 17 個比率變量采用聚類分析(CLUSTER)和非度量多維尺度(NMDS)開展群體間多元統計分析。本研究中因樣本采集難度的原因, 樣本規格(以殼長SL 反映)在種群內、種群間均差異較大。為了減少樣本規格的影響, 形態學數據按照Reist(1986), 對種群間每個樣本殼長(SL)進行常用對數(lg)轉換并作為協變量進行ANCOVA 分析(Sokalet al, 1981)。用9 個測量指標變量進行種群的線性判別分析(LDA), 同樣為了減少樣本尺寸的影響, 對每個測量指標變量(x)使用lg(x)轉換后進行LDA 分析。
2 結果
2.1 種群的描述
各海區采集的皺紋盤鮑樣本的測量指標變量的均值和標準偏差如表1 所示, 群體間及群體內均有很大的變異性。個體總平均殼長為85.17 mm, 種群的平均殼高度差異較大。本次調查記錄的最大樣本位于大連獐子島(ZZD), 殼長SL 為120.25 mm, 最小的在老偏島(LPD), 高度為2.15 mm。我們發現, 大于100 mm的樣本主要來自獐子島(ZZD, 44.14%)、大欽島(23.42%)。另一方面, 發現所有群體在所測量指標中均存在重要的異質性, 且緯度靠北的大個群體變異系數更大, 如老偏島和獐子島變異系數分別為24.18%和21.12%。
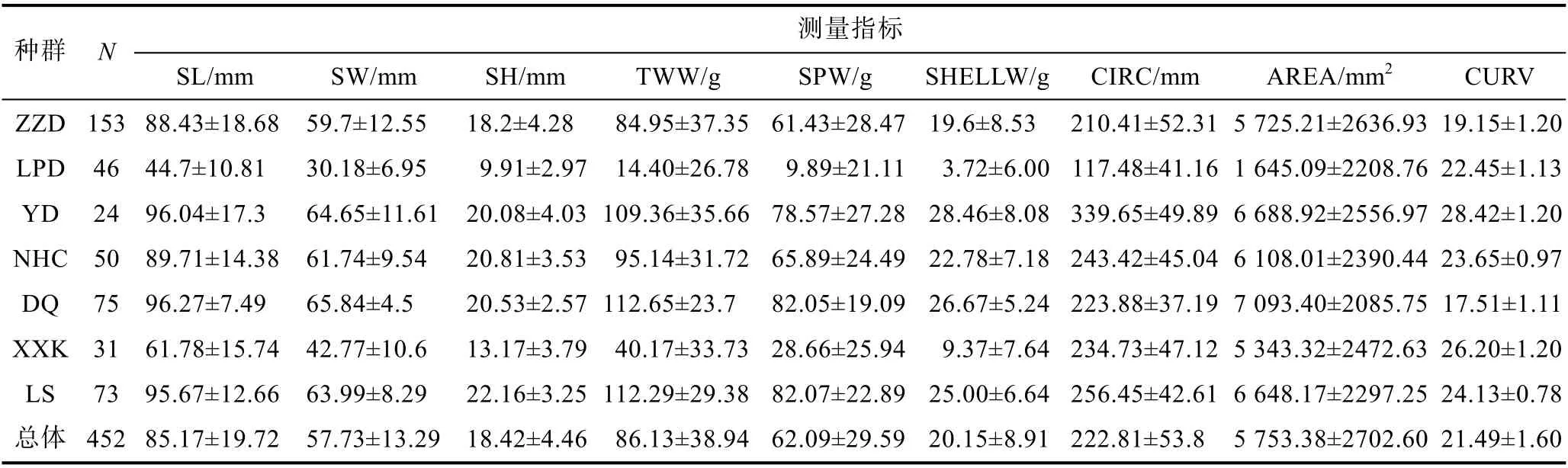
表1 皺紋盤鮑各采樣點樣本量及形態和重量相關測量指標描述性統計Tab.1 Descriptive statistics of sample size, morphological and weight-related characteristics of Pacific abalone at each sample site
2.2 相對殼體生長分析
本研究測定了所有種群樣本稱量指標變量(TWW、SPW 和SHELLW)與殼生長變量(SL、SW 和SH)之間的異速生長關系。整合所有種群異速生長線性方程和相關系數如圖2 所示。Student’st檢驗結果表明, 所有三種異速生長類型(等速生長和正向、負向異速生長)均可在群體內觀察到。因此, 為了更好地解釋數據, 我們將種群的異速生長類型分為如下組:I 組顯示負向異速生長(negative allometry); II 組顯示等速生長(isometry); III 組顯示正向異速生長(positive allometry)。
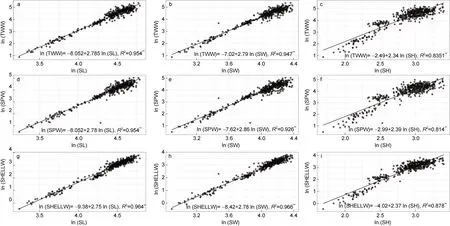
圖2 皺紋盤鮑所有采樣點樣本貝殼形態測量指標和重量相關指標的異速生長關系Fig.2 Allometric relationships between shell morphometric and body weight characters of all samples
2.2.1 全濕重TWW 與殼形態指標(SL、SW 和SH)關系 回歸分析表明, 所采集皺紋盤鮑樣本的全濕重TWW 和3 個殼形態指標(SL、SW 和SH)之間存在顯著的線性相關關系(P<0.01)。在全濕重TWW 和殼長 SL 關系方面, 在合并群體的決定系數為0.954 (P<0.01, 圖 2a), 種群間的決定系數為0.553 (DQ)~0.987 (XXK)。b值在2.07~2.933 之間波動。在全濕重TWW 和殼長SL 關系方面, I 組由DQ 和LS種群組成, II 組由LPD、YD 和ZZD 種群組成, III 組由NHC 和XXK 組成。在全濕重TWW 和殼寬SW 關系方面, 在合并群體的決定系數為0.947 (P<0.01, 圖2a),種群間的決定系數為0.557 (DQ)~0.991 (XXK)。b值在2.18~3.05 之間波動。經Student’st檢驗, I 組、II 組、III 分組與上述全濕重TWW 和殼長SL 關系結論相同。然而, 由全濕重TWW 和殼高SH 關系擬合b值經Student’st檢驗, III 組由LPD 和XXK 組成, 其余群體均為I 組。
2.2.2 軟體部重SPW 與殼形態指標(SL、SW 和SH)關系 回歸分析表明, 所采集皺紋盤鮑樣本的軟體部重SPW 和3 個殼形態指標(SL、SW 和SH)之間存在顯著的線性相關關系(P<0.01)。在軟體部重SPW和殼長 SL 關系方面, 在合并群體的決定系數為0.933 (P<0.01, 圖 2d), 種群間的決定系數為0.475 (DQ)~0.989 (XXK)。b值在2.44~3.07 之間波動。在軟體部重SPW 和殼長SL 關系方面, I 組由LS、ZZD、YD 和DQ 種群組成, II 組由LPD 種群組成, III組由NHC、XXK 組成。在全濕重TWW 和殼寬SW關系方面, 與TWW 和SL 關系類似, 經Student’st檢驗, I 組、II 組、III 分組與上述全濕重TWW 和殼長SL 關系結論相同。然而, 由全濕重TWW 和殼高SH關系擬合b值經經Student’st檢驗, III 組由LPD 和XXK 組成, 其余群體均為I 組。
2.2.3 干殼重SHELLW 與殼形態指標(SL、SW 和SH)關系 回歸分析表明, 所采集皺紋盤鮑樣本的干殼重SHELLW 和3 個殼形態指標(SL、SW 和SH)之間存在顯著的線性相關關系(P<0.01)。在干殼重SHELLW 和殼長SL 關系方面, 在合并群體的決定系數為 0.964 (P<0.01, 圖 2g), 種群間的決定系數為0.546 (LS)~0.987 (LPD)。b值在1.95~2.94 之間波動。在軟體部重SPW 和殼長SL 關系方面, I 組由LS、NHC、YD 和DQ 種群組成, II 組由LPD、ZZD 種群組成, III 組由XXK 組成。在干殼重SHELLW 和殼寬SW 關系方面, 與TWW 和SL 關系類似, 經Student’st檢驗, I 組、II 組、III 分組與上述全濕重TWW 和殼長SL 關系結論相同。然而, 由干殼重SHELLW 和殼高SH 關系擬合b值經經Student’st檢驗, III 組由LPD和XXK 組成, 其余群體均為I 組。
2.3 殼特征的比較分析
各海區采集的皺紋盤鮑樣本的測量指標間17 個比率變量的均值和標準偏差如表2 所示, 與測量指標變量相似, 比率變量在群體間及群體內均有很大的變異性。采用不同的分類或判別方法研究了所采樣點皺紋盤鮑貝殼形態特征的變異性。測量指標比率變量的協方差分析表明, 除SHELLW/SPW 變量外其他變量表示的殼形態間有顯著差異(P<0.05), 這一結果揭示了各群體之間存在重要的表型變異。通過測量指標間比率變量的馬氏距離(Mahalanobis distance,D2)對種群進行了區分, 結果顯示不同地點的種群的殼形態存在顯著差異, 聚類分析CLUSTER 可以分為2 個不同的生物地理群體(圖3): 由XXK、NHC 和LPD所采集的群體組成的一組, 以及由剩余的ZZD、YD、DQ 和LS 組成的另一組。對基于比率多變量的NMDS分析基本證實了馬氏距離聚類的結果, 從圖4 可以大體看出XXK、NHC 和LPD 集中于坐標軸的右中位置。基于Fisher 判別法, 建立了線性判別標準化典型判別函數系數矩陣, 同時基于該系數矩陣構建線性判別函數圖(圖5), 未能很好地區分各個群樣本, 其中僅62.6%的原始分組觀察值已正確地分類到各群體內,針對分析中的各樣本觀察值進行交叉驗證。在交叉驗證中, 每一個觀察值都會依據從該觀察值之外的所有觀察值衍生的函數進行分類, 結果顯示僅58.0%的交叉驗證已分組觀察值已正確地分類。
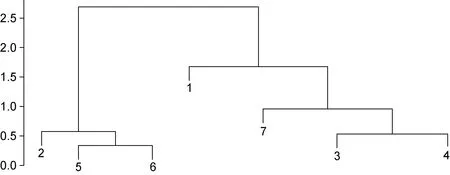
圖3 皺紋盤鮑各采樣點群體基于測量指標間比率變量馬氏距離的聚類分析Fig.3 The Mahalanobis-distance-based cluster analysis for ratio variates from morphological and weight related characters of Pacific abalone populations
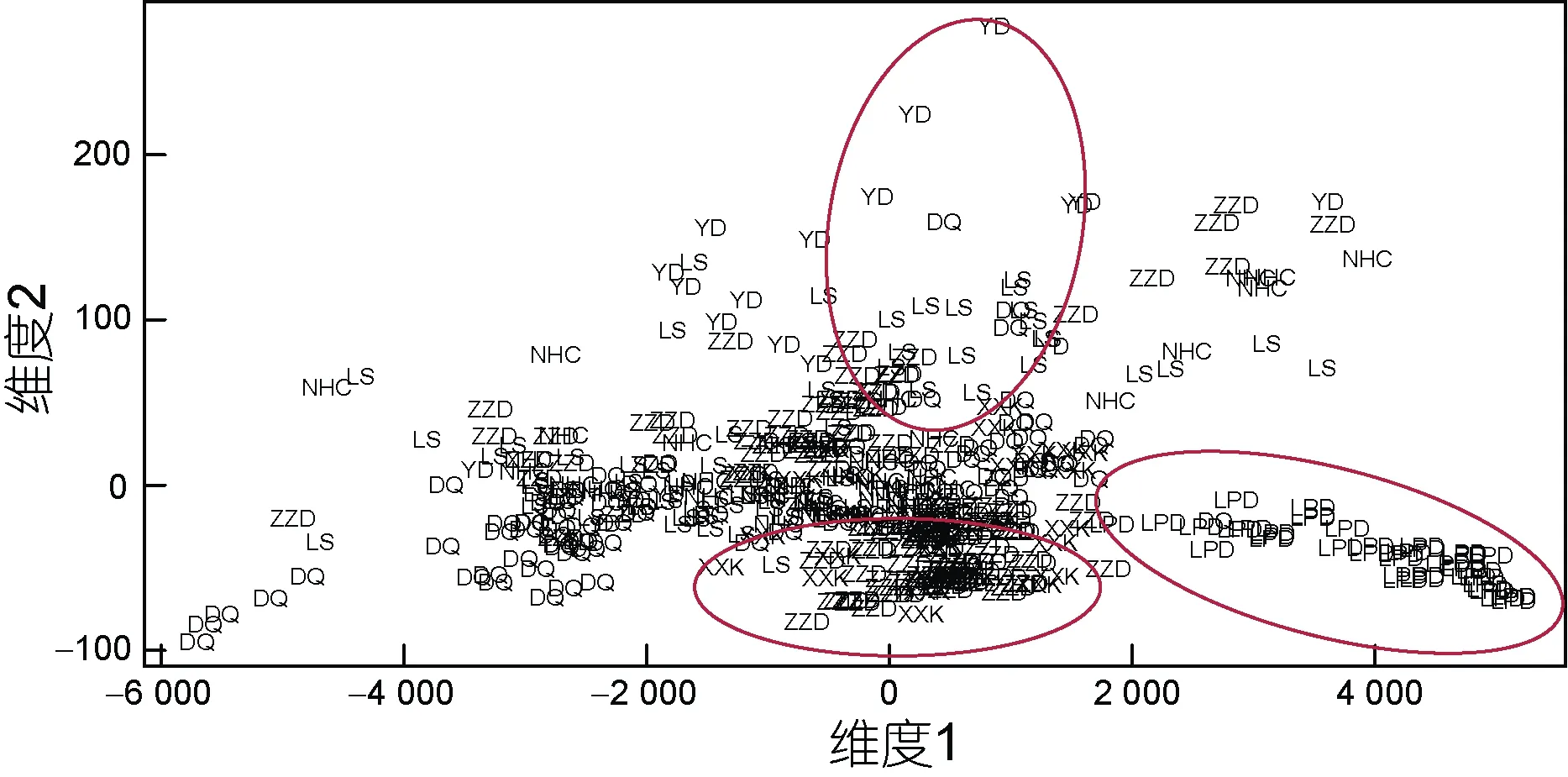
圖4 皺紋盤鮑所有采樣點樣本非度量多維尺度分析Fig.4 Multidimensional scaling plots of weight ratios of Pacific abalone samples at all sample sites
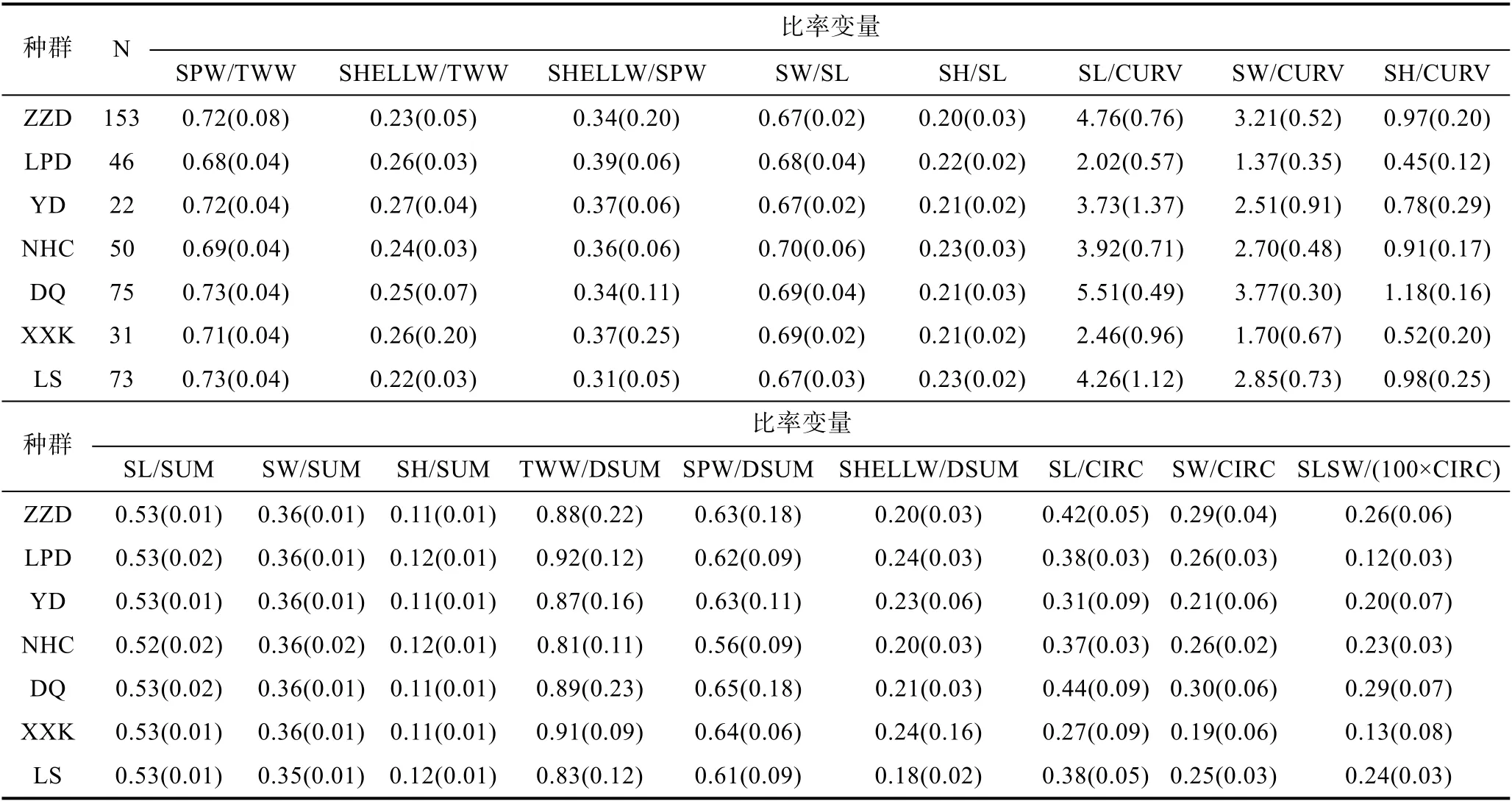
表2 皺紋盤鮑各采樣點測量指標間比率變量的描述性統計Tab.2 Descriptive statistics of sample size and ratios from morphological and weight-related characters of Pacific abalone at each sample site
3 討論
形態測量分析在鮑物種鑒定中應用較廣(Geigeret al, 2012)。在我國, 因鮑種間雜交的產業化應用, 形態測量分析也在鮑純種和雜交種區分中有報道, 相關研究結果表明該分析方法具有重要應用價值(吳玉云, 2001; 張儀方, 2019)。與上述研究的樣本來源不同,本研究樣本來自我國沿海7 個自然海區采樣點的皺紋盤鮑, 分別從貝殼形態變化和相對生長情況進行了調查和測定分析, 從結果看雖然未能完全將所有樣本按采樣區分開, 但大致能分為2 個組, 且該結果能分別從各多元統計方法的結果中相互印證。
有關鮑的生長研究多集中于絕對生長的研究,如Day 等(1992)綜述分析了18 種經濟鮑種不同的生長曲線方程。貽貝、珠母貝等雙殼貝類以及海水魚類中, 異速生長方面的研究較多(Gardner, 1996; Nurul Aminet al, 2004; Bellaaj-Zouariet al, 2012), 其中體重和體長的異速生長系數由線性回歸方程的指數b表示; 在魚類和貝類研究表明當b值接近3 時, 表示等速生長, 而與b=3 顯示顯著不同的值表示異速生長(Nurul Aminet al, 2008; Najmudeen, 2015)。在鮑科動物中, 不同性狀間的異速生長報道相對較少。在本研究中, 我國皺紋盤鮑不同自然海區采樣點所有樣本體重相關的測量指標(TWW、SPW 和SHELLW)與殼形態(SL、SW 和SH)間的線性回歸方程指數b大多在2.34~2.89 之間。為比較群體間相關指標異速生長的變異, 本研究以整合所有群體樣本的b值作為理論值,通過 Student’st檢驗發現正向異速生長(positive allometry)的群體集中在LPD、NHC 和XXK, 一般認為關于總重和殼長的相對增長反映了生態因素對貝類生長的影響, 即環境條件有利于物種的生長和發展(Bellaaj-Zouariet al, 2012), 本研究結果也反映了三個海區的環境條件有利于皺紋盤鮑生長, 比如據有關海藻調查報告顯示山東南隍城島(NHC)大型海藻豐度是國內最高的海區, 不僅調查點大型海藻平均生物量達18 000 g/m2, 且從海藻組成上以皺紋盤鮑喜食的裙帶菜、海帶為主(章守宇等, 2020); 而大連老偏島LPD 和榮成XXK 是最近兩年新發現皺紋盤鮑自然繁殖幼鮑和成體的海區, 由于皺紋盤鮑自然海區繁殖生態條件苛刻, 如孵化后幼體需舔食珊瑚礁微藻(crustose coralline algae, CAA), 稚幼鮑攝食分布較淺的藻類(turf algae)以及成體需大型海藻床(Takamiet al, 2018), 皺紋盤鮑在自然海區繁殖個體的發現也一定程度證明這兩個海區生態的改良。同時,本研究嘗試以各采樣點群體樣本 TWW、SPW、SHELLW 與SL、SW、SH 間的9 個線性回歸方程系數組成的矩陣進行聚類和判別分析, 但可能由于各系數相差較小、變異度較低, 未能區分本研究中皺紋盤鮑各采樣點地理群體。
形態比率變量的ANCOVA 分析表明, 本研究各采樣點皺紋盤鮑種群間形態具有重要的異質性。在采樣中本著隨機采樣原則, 各群體均由皺紋盤鮑大型成年和幼年個體組成的, 這表明皺紋盤鮑在這些采樣點中已經找到了一個適合生長和繁殖的環境。我們的研究結果進一步證明了皺紋盤鮑自然群體中表現出殼形態變異, 這與Kobayashi 等(1996)的研究結果一致。皺紋盤鮑種群的殼形態和異速生長的變異可能是由于當地條件和環境因素的差異, 特別是波浪作用、溫度、海水酸化影響了軟體組織的生長和發育,這在其他腹足綱貝類報道較多(Bouldinget al, 1999;Solaset al, 2013; Carroet al, 2019)。它也可能與種群密度和食物資源的可獲得性有關。事實上, 許多調查已經得出結論, 因為對食物的競爭, 低密度種群的增長比高密度種群的增長更快, 可能更為重要的是越來越多的報道中受全球變暖的影響, 皺紋盤鮑的生長和繁殖受海膽種群的擴張而影響更大(Strainet al,2013; Rogers-Bennettet al, 2019)。
本研究結果表明皺紋盤鮑各采樣點樣本的形態特征區分與其地理分布不相吻合。本研究中利用形態比率變量(17 個)的馬氏距離對各采樣點皺紋盤鮑群體進行聚類分析, 馬氏距離可以克服變量之間的相關性干擾, 并且消除各變量量綱的影響, 結果顯示大連的老偏島(LPD)、圓島(YD)以及榮成(XXK)聚為一組, 剩余4 個群體聚為另一組, 而且基于標準化后的測量變量(9 個)的線性判別分析(LDA)也印證了這一結果, LDA 分析采用Fisher 判別法, 該方法的優點是與主成分分析有關, 而對各變量的分布、方差等都無限制。同時, 非度量多維尺度NMDS 分析雖然不能將所有個體完全區分, 但LPD、XXK 和YD 群體基本上可單獨區分, 剩余群體的樣本則混為一組。造成這一結果的原因, 根據相關單位的生產記錄, LPD、YD和XXK 雖曾大規模底播皺紋盤鮑人工苗種, 但后一組的青島LS、大連ZZD、煙臺NHC、DQ 作為20世紀90 年代皺紋盤鮑底播鮑的主產區, 其底播鮑的數量還是面積都遠遠高于前者。雖然本研究可以確定采樣皺紋盤鮑樣本均為各采樣點的自然繁殖的個體,但這些樣本的親本遺傳來源不確定(未經人工繁育的野生個體繁育而來, 或底播人工苗種繁育而來)可能顯著影響了采樣樣本的形態變異。貝類由于個體繁殖力強, 人工苗種往往來源于幾個雄性親本, 再加上海區間底播投苗來源不清晰, 這可能造成雖然地理相隔很遠(如LS 和ZZD 地理距離遠), 但形態比率變量仍聚類為一組。這也可能是LDA 未能將所有個體完全清晰按采樣點分開的原因(成功率僅62.6%)。
樣本質量和測量方法的準確性是采用形態特征區分皺紋盤鮑群體的關鍵。無論在本研究的采樣過程還是樣本處理、測定中采用了嚴格的質量控制。比如為防止季節對樣本測量指標的影響, 特別是重量相關指標的影響, 本研究均采用夏季7 月份采樣的樣本;采樣地點的選擇是至少近10 年未由人工苗種底播的海區, 并且采樣后嚴格觀察是否有因海流等遷徙過來的人工底播鮑, 本實驗中所有采樣點雖有700 余個標本采集, 但實際分析中僅有452 個樣本進入數據分析。在實驗中測量時發現皺紋盤鮑殼高和粗糙度的測量會受到測量人和拍照獲取殼輪廓影響很大, 實驗中殼高SH 的測定采用同一測量人多次測量(n>5)取平均的方法, 而為獲取鮑殼外緣輪廓的凸包值, 實驗采用同一天暗室光源拍照獲取所有貝類輪廓的方式進行。此外, 在數據分析中, 為減少樣本規格差異對結果的影響, 本研究采用數據對數轉化, 并以對數轉化后的殼長作為協變量來進行比率變量的協方差分析。盡管如此, 樣本間規格差異大、樣本量較小, 以及人工苗種的混雜或者遺傳影響可能是本研究缺陷,未來研究需加強該方面工作。
4 結論
研究以我國沿海采集的7 個皺紋盤鮑群體作為研究材料, 分析了群體內和群體間貝殼形態和體重相對生長的變異。協方差分析揭示形態特征變量(比率指標)在群體間存在顯著變異; 非度量多維尺度分析和馬氏距離將皺紋盤鮑群體分為2 個組; 異速生長研究揭示了殼形態特征與皺紋盤鮑群體中異速生長差異之間的高度線性相關性, 異速生長程度呈現一定的群體區別; 然而線性判別分析未能較好地將各種群樣本分類。該研究初步表明皺紋盤鮑表現出與我國沿海環境和/或生態條件差異相關的表型可塑性,然而也可能受到人工底播影響造成的遺傳影響。本研究將對于皺紋盤鮑漁業資源管理如管理單位的界定,以及為今后全面評價不同地理群體的遺傳變異等提供參考。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
中學生數理化·八年級物理人教版(2019年9期)2019-11-25 07:33:02
華人時刊(2019年13期)2019-11-17 14:59:54
中學生數理化·八年級物理人教版(2019年3期)2019-04-25 06:20:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
中學生數理化·八年級物理人教版(2018年3期)2018-05-31 08:52:45
數學小靈通(1-2年級)(2017年10期)2017-11-08 08:39:45
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
