狄氏副擬桿菌胞外多糖粗提物的免疫調(diào)節(jié)活性*
2022-11-28 12:30:14祝陽露張倩李四菊曹雪芹葉思敏謝智勇
中山大學學報(自然科學版)(中英文) 2022年6期
祝陽露,張倩,李四菊,曹雪芹,葉思敏,謝智勇
中山大學藥學院(深圳),廣東 深圳 518107
微生物胞外多糖是指微生物在代謝生長過程中分泌到細胞外或與細胞疏松連接的長鏈高分子聚合物[1]。作為活性的生物大分子,微生物胞外多糖已經(jīng)被報道具有多樣的活性,包括抗腫瘤、抗氧化、免疫調(diào)節(jié)、降血糖[2-5]等。同時相較于植物以及動物多糖,由于培養(yǎng)周期短、培養(yǎng)條件易于控制、受地理和氣候等因素的影響較少等優(yōu)點,微生物胞外多糖引起了越來越多研究者的興趣。
作為腸道共生菌,狄氏副擬桿菌Parabacteroides distasonis由于具有減輕肥胖、緩解結(jié)腸炎、結(jié)腸癌和多發(fā)性硬化[6-9]等多種生物活性近些年來受到了日益增加的關(guān)注。但目前已有的研究主要集中于對狄氏副擬桿菌整個菌體的研究,卻未曾有關(guān)于狄氏副擬桿菌胞外多糖生理活性的研究。胞外多糖作為狄氏副擬桿菌物質(zhì)基礎(chǔ)之一,對胞外多糖研究的匱乏無疑會限制該菌未來的發(fā)展利用。因此本研究首次從狄氏副擬桿菌提取得到胞外多糖粗提物,并研究其免疫調(diào)節(jié)活性,為胞外多糖在免疫增強劑方面的開發(fā)利用提供了一定的參考意義,也為從胞外多糖角度探討腸道菌群的有益功能提供了新的思路。
1 實驗部分
1.1 儀器、試劑及材料
5910R 冷凍離心機,購自德國Eppendorf公司;LightCycler 96 實時熒光定量PCR 儀,購自瑞士Roche 公司;Epoch 酶標儀,購自美國BioTek 公司;UV-2600 紫外光譜儀、高效液相色譜儀(HPLC,LC-20AT),購自日本島津公司;Spectra-Max i3x 熒光定量酶標儀,購自奧地利Molecular Devices公司。
Parabacteroides distasonis菌株,購自廣東省微生物保藏中心,編號為ATCC8503;RAW 264.7細胞株,購自美國菌種保藏中心。
噻唑藍(MTT)、脂多糖(LPS),購自Sigma-Aldrich公司;DMEM 培養(yǎng)基、胎牛血清、青霉素-鏈霉素,購自Gibco 生物科技公司;腦心浸出液培養(yǎng)基(BHI),購自廣東環(huán)凱;木瓜蛋白酶,購自北京索萊寶;BCA 蛋白定量試劑盒,購自北京康為世紀;PCR 引物,由上海生工合成;NO 試劑盒,購自上海碧云天生物技術(shù)有限公司;活性氧試劑盒,購自武漢伊萊瑞特;單糖標準品(甘露糖Man、鼠李糖Rha、半乳糖Gal、葡萄糖醛酸GluA、半乳糖醛酸GalA、葡萄糖Glu、阿拉伯糖Ara、巖藻糖Fuc),購自上海吉至公司;其他試劑均為分析純。
1.2 胞外多糖提取
將狄氏副擬桿菌甘油凍存管于37 ℃融化以后,用接種環(huán)蘸取少量菌液于固體平板劃線并于37 ℃厭氧培養(yǎng)數(shù)日,待狄氏副擬桿菌復蘇后,選取單克隆菌落接種于腦心浸出液液體培養(yǎng)基中于37 ℃厭氧培養(yǎng)48 h,并以3%的比例擴大培養(yǎng)2 代以后收集發(fā)酵液。將發(fā)酵液離心(4 000 r/min,4 ℃,30 min)后,取上清于100 ℃水浴加熱10 min 以滅活其中的酶,然后將離心后的上清液減壓濃縮至原體積的1/3 后,加4 倍體積的冰乙醇攪拌混合,并于4 ℃靜置過夜,離心收集沉淀,即得胞外多糖粗提取物。將沉淀加少量去離子水溶解后加木瓜蛋白酶(終質(zhì)量濃度為1 mg/mL)于65 ℃攪拌反應(yīng)3 h后,100 ℃滅酶10 min以后,離心收集上清液,再用Sevag 試劑(V三氯甲烷∶V正丁醇=4∶1)反復多次除去蛋白,待中間白色蛋白層消失以后,收集上層液體經(jīng)減壓濃縮、透析、冷凍干燥得到除蛋白后的胞外多糖粗提取物。由于胞外多糖粗提取物中含有大量的色素可能會影響其純度及生物活性,繼而采用AB-8 大孔吸附樹脂[10]對除蛋白后的胞外多糖粗提取物進行脫色,并將脫色后的胞外多糖粗提取物,命名為PEP。
1.3 總糖含量、蛋白質(zhì)含量、紫外光譜測定
以葡萄糖為標準,用苯酚硫酸法[11]對PEP 的總糖含量進行測定。
以牛血清白蛋白為標準,按照BCA 試劑盒的操作對PEP的蛋白質(zhì)含量進行測定。
用紫外分光光度計在200~400 nm 范圍內(nèi)對PEP 溶液(1 mg/mL)進行掃描,并繪制紫外光譜圖。
1.4 單糖組成測定
采用1-苯基-3-甲基-5-吡唑啉酮(PMP)柱前衍生化高效液相色譜法[12]對PEP 的單糖組成進行測定。
PEP 的水解:稱取5 mg PEP 于反應(yīng)管中,加入2 mL 的3 mol/L TFA 溶液于135 ℃油浴3 h 進 行水解。水解完成待冷卻后,重復3 次加入5 mL甲醇洗滌溶液并減壓濃縮至干,以除去殘留的三氟乙酸。
PEP及單糖標準品的衍生化:將水解完全并減壓蒸干的PEP 糖聚物用800 μL 去離子水溶解,取100 μL 溶 液,加 入0.5 mol/L 的PMP 甲 醇 溶 液 和0.3 mol/L 的NaOH 溶液各100 μL,并于70 ℃放置30 min,待恢復室溫后加入105 μL 0.3 mol/L 的HCl溶液進行中和,并加入200 μL去離子水進行稀釋,而后重復3次加入600 μL三氯甲烷進行萃取以除去溶液中殘留的PMP,最后水層過0.22 μm濾膜過濾后于HPLC進行分析。各單糖標準品按照相同的方法進行衍生化最后進樣HPLC進行分析。
HPLC 條件為,色譜柱:C18柱(4.6 mm×250 mm,5 μm,Waters 公司);流動相(體積比):0.05 mol/L 磷酸鹽緩沖液(pH6.7)∶乙腈=83∶17;流速:1 mL/min;檢測器:二極管陣列檢測器;檢測波長:250 nm;進樣量:10 μL。
1.5 PEP的免疫調(diào)節(jié)活性
本實驗以小鼠單核巨噬細胞RAW 264.7 為模型,對PEP的免疫調(diào)節(jié)活性進行研究。
1.5.1 細胞培養(yǎng)RAW 264.7 細胞用含有φ=10%胎牛血清和φ=1%的雙抗(含10 000 U/mL 的青霉素和10 000 μg/mL的鏈霉素)的DMEM完全培養(yǎng)基在37 ℃培養(yǎng)箱(φ=5%CO2)培養(yǎng),每隔2 d傳代。
1.5.2 細胞活性測定取對數(shù)生長期的細胞,以1×106個/mL 的密度接種于96 孔板(100 μL/孔),過夜培養(yǎng)待細胞貼壁后,棄培養(yǎng)液,空白對照組加100 μL 完全培養(yǎng)基,陽性對照組加100 μL 1 μg/mL 的LPS 溶液,實驗組加100 μL 質(zhì)量濃度分別為50、100、200 μg/mL 的PEP 溶液,培養(yǎng)24 h 后,加入10 μL MTT 溶液(5 mg/mL)避光培養(yǎng)4 h 后,吸棄上清加入200 μL DMSO 溶液于搖床振搖10 min 后于490 nm 測定每孔吸光值。將空白對照組細胞的存活率視為100%。
1.5.3 NO和活性氧測定 按照1.5.2的接種和給藥方式培養(yǎng)24 h 后,用NO 試劑盒測定細胞產(chǎn)生的NO 濃度同時細胞的活性氧產(chǎn)生參照活性氧試劑盒測定。
1.5.4 中性紅吞噬實驗按照1.5.2 的接種和給藥方式培養(yǎng)24 h 后,棄培養(yǎng)液,加入0.075%的中性紅溶液避光孵育45 min 后,棄上清,細胞用PBS溶液洗3次后,加入醇酸裂解液(V乙醇∶V冰醋酸=1∶1)后于培養(yǎng)箱避光放置2 h 后于540 nm 處測定每孔吸光度。
1.5.5 相關(guān)細胞因子表達水平測定細胞以5×105個/mL 接種于6 孔板(2 mL/孔),待細胞貼壁后,吸棄培養(yǎng)液,每孔分別加入2 mL 的DMEM完全培養(yǎng)基,LPS溶液(1 μg/mL)和各質(zhì)量濃度梯度的PEP 溶液(50、100、200 μg/mL)培養(yǎng)24 h 后,參考伍芳芳[13]的方法提取細胞總RNA,通過逆轉(zhuǎn)錄試劑盒合成cDNA 并經(jīng)過37 ℃15 min,50 ℃5 min,98 ℃5 min 反應(yīng)后,利用熒光定量PCR 儀檢測各細胞因子(引物序列見表1)mRNA的相對表達,反應(yīng)體系為95 ℃預(yù)變性30 s;95 ℃5 s,60 ℃10 s,75 ℃20 s,重 復40 個循環(huán);95 ℃1 min,40 ℃1 min,65 ℃1 s,95 ℃25 個采集點;40 ℃冷卻30 s。以GAPDH 為內(nèi)參基因,用2-△△Ct方法計算各細胞因子的相對表達量。

表1 PCR引物序列Table 1 PCR primer sequences
1.6 數(shù)據(jù)分析
實驗重復3 次,實驗中數(shù)據(jù)以均值±標準偏差的形式表示,采用IBM SPSS 20.0對數(shù)據(jù)進行單因素方差分析,P<0.05表示存在顯著性差異。
2 結(jié) 果
2.1 PEP化學組成、紫外光譜分析
以葡萄糖的質(zhì)量濃度(mg/mL)為橫坐標(x),各質(zhì)量濃度葡萄糖在490 nm 處的吸光值為縱坐標(y),擬合的回歸方程為y= 4.166 3x+0.064 6,r2=0.996 6,表明具有良好的線性關(guān)系。將PEP 在490 nm 處的吸光值代入方程,由此計算出PEP 的總糖含量為(62.34±1.99)%。以牛血清白蛋白的濃度為橫坐標(x),各濃度牛血清白蛋白在562 nm處的吸光值為縱坐標(y),擬合的回歸方程為y= 0.842 4x+0.110 8,r2=0.997 3,表明具有良好的線性關(guān)系。將PEP 在562 nm 處的吸光值代入方程,由此計算出PEP 的蛋白質(zhì)含量為(5.66 ±0.44)%。如圖1a 所示,PEP 在260 和280 nm 均沒有出現(xiàn)明顯的吸收峰,說明PEP 不含或含有較少的核酸和蛋白質(zhì),該結(jié)果與蛋白質(zhì)含量測定結(jié)果一致。
2.2 PEP的單糖組成分析
如圖1b 所示,通過將PEP 的液相色譜圖與混合單糖標準品衍生化后的液相色譜圖進行對比,根據(jù)保留時間和峰面積計算可知,PEP主要由甘露糖、鼠李糖、葡萄糖、半乳糖及少量的巖藻糖組成,其組成比例為19.35∶8.70∶5.24∶8.17∶1.00,表明PEP是一個由5種單糖組成的雜多糖。
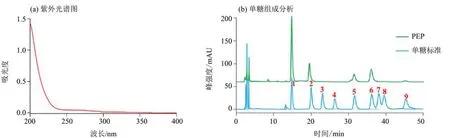
圖1 PEP的紫外光譜圖及單糖組成分析Fig.1 UV spectrum and monosaccharide composition analysis of PEP
2.3 PEP對RAW 264.7細胞增殖的影響
采用MTT 實驗考察PEP 對巨噬細胞的毒性。如圖2a 所示,在50~200 μg/mL 范圍內(nèi),PEP 對細胞沒有毒性,相反可以促進細胞的生長,表明該濃度范圍可用于后續(xù)的實驗。
2.4 PEP 對RAW 264.7 細胞釋放NO、活性氧的影響
NO、活性氧是巨噬細胞在活化過程中產(chǎn)生的一些信號分子,在宿主防御病原體侵害等方面占據(jù)重要地位[14-15]。因此我們運用試劑盒檢測了細胞釋放NO、活性氧的情況。如圖2b、2c 所示,與空白對照組比較,PEP可以呈濃度依賴地促進巨噬細胞分泌NO、活性氧,上述結(jié)果表明PEP 可以通過增加巨噬細胞釋放NO、活性氧等信號分子行使其免疫調(diào)節(jié)功能。
2.5 PEP對RAW 264.7細胞吞噬活性的影響
吞噬活性提高是巨噬細胞被激活后的顯著特征,巨噬細胞的吞噬作用可以幫助機體有效抵抗和清除病原體[16]。因此我們采用中性紅實驗測定了RAW 264.7 的吞噬功能。如圖2d 所示,與空白對照組比較,PEP干預(yù)后,巨噬細胞的吞噬活性明顯增加,并且呈現(xiàn)濃度依賴關(guān)系,表明PEP 可以增強巨噬細胞的吞噬功能。
2.6 PEP對RAW 264.7細胞中細胞因子的影響
3 個典型的細胞因子IL-1β、IL-6、TNF-α 在免疫調(diào)節(jié)中發(fā)揮重要作用,可以介導和調(diào)節(jié)免疫反應(yīng)[17]。IL-1β和IL-6是免疫應(yīng)答過渡階段的重要信號分子,對維持免疫穩(wěn)態(tài)至關(guān)重要[18]。同時作為一種多功能細胞因子,TNF-α可以參與對外來感染的防御和清除,促進受損組織的修復,對巨噬細胞的生長和功能起決定性作用[19]。因此,我們采用實時熒光定量PCR 檢測了IL-1β、IL-6、TNF-α 3個細胞因子在基因水平的相對表達量。如圖3a~c所示,與空白對照組比較,細胞中IL-1β、IL-6、TNF-α 表達量與PEP 的質(zhì)量濃度呈正相關(guān),表明PEP可以通過提高細胞因子基因的表達水平增強細胞的免疫調(diào)節(jié)能力。
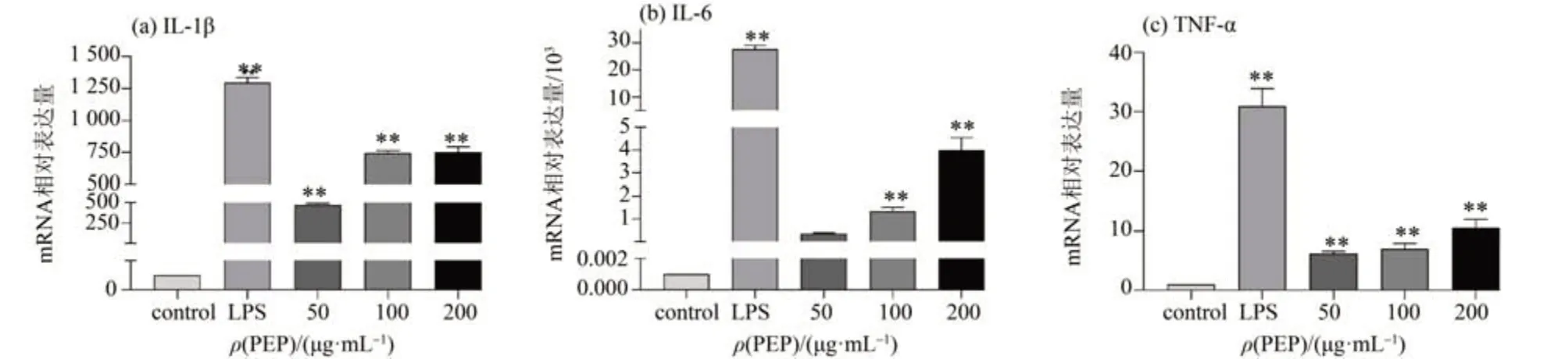
圖3 PEP對RAW 264.7細胞的細胞因子mRNA表達的影響Fig.3 Effects of PEP on the mRNA expression of cytokines in RAW 264.7 cells
3 討論與小結(jié)
微生物胞外多糖,尤其是來自于腸道微生物的多糖,由于其屬于微生物重要的活性代謝產(chǎn)物,近年來已經(jīng)成為研究熱點之一。目前對胞外多糖的提取主要采用水提取,高濃度乙醇沉淀的方法,本研究也采用該方法對狄氏副擬桿菌胞外多糖進行了初步提取。初步提取得到的胞外多糖往往含有大量的蛋白質(zhì)和色素等雜質(zhì)會干擾到多糖的純度以及活性,因此會采用一系列除蛋白以及脫色的操作對胞外多糖進行下一步的純化。本研究采用酶法與Sevag 試劑相結(jié)合的方法對胞外多糖粗提物進行脫蛋白處理,一方面由于采用木瓜蛋白酶從而大大提高了除蛋白的效率,另一方面也減少了Sevag 試劑的用量,這對于日后胞外多糖的擴大生產(chǎn)具有良好的借鑒意義。樹脂脫色具有條件溫和,對多糖的保留率高等優(yōu)點,因此我們采用了AB-8 大孔吸附樹脂對胞外多糖進行脫色處理,從而避免出現(xiàn)脫色之余多糖損失過多以及降解等問題。
先天免疫反應(yīng)是機體抵抗病原體入侵和外來物質(zhì)的第一個屏障[20]。一些細胞,如巨噬細胞不僅在先天性免疫中發(fā)揮著重要作用,同時也作為橋梁,將非特異性免疫與特異性免疫連接在一起[21]。當巨噬細胞被激活以后,可以通過釋放出NO、活性氧、細胞因子以及增強吞噬等途徑來實現(xiàn)保護機體免受病原體侵害的作用,因此巨噬細胞常常被用于體外免疫活性的研究[22]。相關(guān)研究表明,增強免疫系統(tǒng)對免疫抑制、微生物感染、惡性腫瘤等疾病有良好的治療效果[23]。作為生物反應(yīng)調(diào)節(jié)劑,微生物胞外多糖顯示出良好的免疫調(diào)節(jié)活性[24-25]。而狄氏副擬桿菌又是一株被證明具有多種益生活性的腸道共生菌,因此我們提取其胞外多糖,并對其免疫調(diào)節(jié)活性進行探究,我們發(fā)現(xiàn)該胞外多糖是一個純度較高的多糖,并且可以通過促進巨噬細胞增殖、增加NO、活性氧的釋放,增強吞噬功能以及促進細胞因子的表達等多種途徑從而發(fā)揮其良好的免疫調(diào)節(jié)活性。
通過提高免疫功能的免疫療法已成為治療癌癥的臨床方法之一[26]。某些多糖可通過增強免疫力達到抗癌活性。例如,人參多糖RGRP-1b 本身對Huh7 肝癌細胞沒有作用,但可以通過作用于巨噬細胞增強免疫功能,從而在癌癥治療中發(fā)揮作用[27]。此外,柴胡多糖BCAP-1 也具有類似的抗癌作用[28]。由于PEP 具有良好的免疫增強活性,有望在未來作為抗癌藥物的佐劑使用。
綜上所述,本研究首次從狄氏副擬桿菌中提取得到胞外多糖并證明其具有免疫調(diào)節(jié)活性。對狄氏副擬桿菌胞外多糖活性的研究一方面推動了胞外多糖的進一步發(fā)展,為狄氏副擬桿菌胞外多糖的商業(yè)化轉(zhuǎn)化提供了可能,另一方面也為從胞外多糖的角度探討腸道菌群的益生作用具有借鑒作用。
