基于水分遷移分析熱風-熱泵聯(lián)合干燥對蝦肉品質(zhì)的影響
2022-11-29 04:39:40梁鉆好陳海強梁鳳雪余銘
食品與發(fā)酵工業(yè) 2022年22期
關(guān)鍵詞:信號
梁鉆好,陳海強,梁鳳雪,余銘*
1(陽江職業(yè)技術(shù)學院 食品與環(huán)境工程系,廣東 陽江,529566)2(廣東省食品低溫加工工程技術(shù)研究中心,廣東 陽江,529566)
干制是水產(chǎn)品主要加工方式之一。生產(chǎn)上采用傳統(tǒng)日曬或熱風烘干為主,日曬受氣候影響,開放式環(huán)境下產(chǎn)品易受污染,最終水分含量和品質(zhì)難以控制;熱風干燥通過高溫迅速脫水,期間容易發(fā)生蛋白質(zhì)熱變性、脂質(zhì)氧化、美拉德反應等不良生化反應,或出現(xiàn)表面局部硬化現(xiàn)象,使產(chǎn)品品質(zhì)下降[1]。熱泵干燥、真空冷凍干燥、遠紅外干燥等低溫干燥方式在高值水產(chǎn)品干制中的應用研究越來越多。但這些干燥方式也存在不足,如干燥速度慢、時間長,產(chǎn)品泛白、色香味欠缺,穿透力不足,不適宜厚肉產(chǎn)品干燥等[2]。聯(lián)合干燥是指根據(jù)物料特點,兩種或兩種以上干燥方式組合干燥,實現(xiàn)多種干燥方式之間的優(yōu)勢互補,提高干燥速率,改善產(chǎn)品品質(zhì)。如熱泵-熱風聯(lián)合干燥[3]、熱風-真空冷凍聯(lián)合干燥[4]、微波-遠紅外聯(lián)合干燥[5]、微波-真空冷凍聯(lián)合干燥[5]等多種聯(lián)合干燥方式在果蔬、水產(chǎn)品、肉制品和中草藥干制中已有應用研究。
低溫熱泵干燥的特點是原料表面和內(nèi)部的水分蒸發(fā)速率接近,干燥溫度較低(通常為10~40 ℃),可有效保留食品原有營養(yǎng)和功效,節(jié)約能耗[6]。對于蝦而言,高溫加熱一方面可使蝦殼中其他色素分解,另一方面使與蛋白質(zhì)結(jié)合呈藍紫色或藍綠色的耐熱原蝦紅素游離出來,恢復原來的橘紅色,而且蝦肉蛋白質(zhì)也會因高溫加熱變性從而使蝦口感發(fā)生改變,同時提升香氣和風味。這也正是市場上高溫烘干的蝦干色澤和香氣誘人的重要原因。因此,對于蝦這類高溫加熱變色的特殊水產(chǎn)品,單獨采用低溫熱泵干燥,不能滿足市場對產(chǎn)品色澤和香氣的要求。本文采用一段熱風高溫熟制定型、上色、賦香,二段低溫熱泵干燥溫和脫水,利用熱風-熱泵聯(lián)合干燥優(yōu)勢,提高干制對蝦品質(zhì)。另外,干燥是水分蒸發(fā)的過程,食品中水分蒸發(fā)的速度、所蒸發(fā)的水分在食品中的存在狀態(tài)以及水分蒸發(fā)過程所發(fā)生的水分遷移對干燥食品的最終品質(zhì)有著關(guān)鍵影響[7]。低場核磁共振(low-field nuclear magnetic resonance, LF-NMR)可通過測定質(zhì)子在磁場中的弛豫特性,快速、無損、靈敏[8]地檢測食品中自由水、不易流動水和結(jié)合水的含量、分布、活度及遷移率[9],配合核磁共振成像(magnetic resonance imaging, MRI)技術(shù),可形象地還原加工過程中水在食品內(nèi)部的分布及變化過程[10]。如,LF-NMR聯(lián)合MRI技術(shù)可用于檢測天麻干燥過程的含水率[10];獲得熱風干燥花生仁含水率預測模型[11];從3種水態(tài)的遷移和變化規(guī)律闡明超聲波預處理促進扇貝熱泵干燥速率的機理[12];探索微干海參的復水時間與質(zhì)構(gòu)口感的相關(guān)性,從而預測最佳復水時間[13];動態(tài)監(jiān)控復水過程香菇干內(nèi)部水分以確定最佳的熱風干燥條件[14]等。因此,本研究利用LF-NMR技術(shù)監(jiān)控干燥過程以及復水過程的水分吸附動態(tài),從水分遷移角度闡明聯(lián)合干燥技術(shù)對蝦肉肌纖維結(jié)構(gòu)及品質(zhì)的影響機理。
1 材料與方法
1.1 材料與儀器
南美白對蝦(海水養(yǎng)殖)購于陽江市場;鹽酸、硼酸、氧化鎂、甲基紅、亞甲基藍、磷酸二氫鈉、磷酸氫二鈉、氯化鈉,均為分析純。
MesoMR23-040V-I核磁共振成像分析儀,蘇州紐邁分析儀器股份有限公司;EM ACE600真空鍍膜儀,德國萊卡儀器有限公司;EVO MA 15掃描式電子顯微鏡(scanning electron microscope,SEM),德國ZEISS公司;DDC06 TSL低溫熱泵干燥箱,上海精密科學儀器有限公司;DHG-9070A電熱恒溫鼓風干燥箱,成都圣欣科學儀器有限公司;CT3質(zhì)構(gòu)儀,美國博勒飛公司;SY204分析天平,上海佑科儀器儀表有限公司;ATN-300自動凱氏定氮儀,上海洪紀儀器設(shè)備有限公司。
1.2 實驗方法
1.2.1 原料預處理與干燥
挑選完好且大小一致的新鮮對蝦[(15±2) g],清洗干凈,去頭去殼,取蝦肉約3 cm長度(靠近蝦頭部位),根據(jù)前期試驗的工藝優(yōu)化[1],確定一段熱風干燥溫度為90 ℃時蝦肉可獲得最佳品質(zhì),故選擇最佳聯(lián)合干燥條件為:一段90 ℃熱風干燥定型,至蝦完全熟化,記錄熟化時間,取出降溫,二段轉(zhuǎn)20 ℃低溫熱泵機(風速2.0 m/s,相對濕度20%)[15]中進行二段干燥;對照組(CK組)二段轉(zhuǎn)60 ℃熱風干燥(以生產(chǎn)中熱風干燥常用的60 ℃為對照),以對蝦水分含量降至15%以下(約13%)為標準確定最終干燥時間。
CK組:90 ℃熱風干燥2 h - 60 ℃熱風干燥6 h;
聯(lián)合干燥組:90 ℃熱風干燥2 h - 20 ℃熱泵干燥20 h。
1.2.2 理化指標測定
水分含量參照GB 5009.3—2016《食品安全國家標準食品中水分的測定》中的恒重干燥法測定;總揮發(fā)性鹽基氮(total volatile base nitrogen,TVB-N)含量測定參照GB 5009.228—2016《食品安全國家標準食品中揮發(fā)性鹽基氮的測定》中的半微量滴定法。
1.2.3 電鏡掃描[16]
SEM分析加速電壓為10 kV,樣品切片大小為3 mm×3 mm×1 mm,檢測前均貼于有導電膠的檢測臺上,噴金30 s,使樣品表面鍍金約10 nm厚后進行檢測。
1.2.4 質(zhì)構(gòu)分析
干燥蝦肉于45 ℃水浴浸泡2 h,取出用濾紙吸干表面水分,待降至室溫后采用CT3質(zhì)構(gòu)儀測定。
TPA參數(shù):探頭型號TA41,測前下降速度2.00 mm/s,測試速度1.00 mm/s,測后探頭回程速度1.00 mm/s,形變50.0%,觸發(fā)值25 g,循環(huán)次數(shù)2;試驗類型:壓縮。
剪切力參數(shù):夾具型號TA-SBA,測前下降速度2.00 mm/s,測試速度2.00 mm/s,測后探頭回程速度2.00 mm/s,下壓20.0 mm,觸發(fā)值25 g;試驗類型:壓縮。
1.2.5 復水率
干燥蝦肉于45 ℃水浴復水,料水比為1∶100(g∶mL),0、5、15、30、45、60、75、90、120 min測定蝦肉重量,復水率的計算如公式(1)所示:
(1)
式中:Rf,復水率,%;m0,復水前樣品重量,g;mt,復水t時刻后樣品重量,g。
1.2.6 LF-NMR測定與成像分析[9]
干燥過程:二段干燥過程中,定期取出蝦肉,保鮮膜包好后采用低場核磁共振成像儀檢測。儀器內(nèi)1次放入1塊蝦肉進行檢測,檢測完畢后放回干燥箱中繼續(xù)干燥,用于下一個時間點的檢測,每次實驗做5個平行。
復水過程:干燥蝦肉于45 ℃水浴浸泡,0、5、15、30、45、60、75、90、120 min分別取出蝦肉用濾紙吸干表面水分,保鮮膜包好后采用低場核磁共振成像儀檢測。儀器內(nèi)1次放入1塊蝦肉進行檢測,檢測完畢后放回水中繼續(xù)復水,用于下一個時間點的檢測,每次實驗做5個平行。
核磁共振弛豫時間T2測試方法:放入玻璃試管(口徑40 mm),然后將樣品管置于核磁探頭中,使用CPMG序列測試橫向弛豫時間T2。其中參數(shù)設(shè)置如下:磁體溫度32 ℃;重復采樣等待時間TW=3 000.00 ms;重復采樣次數(shù)NS=16;回波時間TE=0.15 ms;回波個數(shù)NECH=9 000;采樣帶寬SW=200 kHz。模擬增益RG1=20.0 db;數(shù)字增益DRG1=3;干燥過程的前置放大器增益PRG=2,復水過程的前置放大器增益PRG=1。
MPI成像分析,參數(shù)設(shè)置:層數(shù)=1;層厚=5.0 mm;視野FOV=100 mm×100 mm;重復采樣次數(shù)Average=2;重復采樣等待時間TR=900.00 ms;回波時間TE=18.125 ms。
1.2.7 數(shù)據(jù)處理
試驗數(shù)據(jù)采用Execl 2010進行處理,并用Origin 9.1作圖,SPSS 17進行統(tǒng)計學分析。
2 結(jié)果與分析
2.1 干燥對蝦的基本特性
2種干燥方式得到的蝦肉水分含量基本一致,均為21%左右,無顯著差異。即用這2種干燥工藝得到的對蝦做比較,具有較好的可比性。但二者的TVB-N含量卻有顯著差異,聯(lián)合干燥組的TVB-N含量顯著降低43.50%(P<0.05),即干燥到相同水分含量的情況下,聯(lián)合干燥方式更有利于延緩蝦肉氧化變質(zhì)。
哦,是這樣的啊!我趕在父親回來之前到家了,把我看到的情況告訴了母親,我發(fā)現(xiàn)母親的臉頰上有兩行淚珠在淌。
熱風干燥和熱泵干燥二者原理不同。熱風干燥通常是通過加熱器加熱空氣實現(xiàn)烘干的,配有風機排除濕空氣,艙體為非密閉空間,艙內(nèi)空氣高溫高濕。而熱泵干燥機是利用逆卡諾循環(huán)原理來對物料進行干燥的,艙內(nèi)密閉,保持低溫低濕,在此條件下,有四大優(yōu)點:(1)微生物繁殖速率減慢;(2)各種生化反應速率受到抑制;(3)在前兩點實現(xiàn)的基礎(chǔ)上,可長時間緩慢脫水,同時保持食品不腐敗;(4)食品內(nèi)外水分散失速率相對一致,溫和脫水,避免熱風干燥情況下快速不均勻的脫水對肌肉組織結(jié)構(gòu)的嚴重破壞。
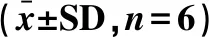
表1 干燥對蝦的基本特性Table 1 Basic characteristics of dried prawn
2.2 干燥方式對蝦肉微觀結(jié)構(gòu)的影響
圖1 為干燥對蝦分別放大400、1 000和4 000倍數(shù)的電鏡掃描圖。如圖所示,CK組蝦肉肌纖維模糊,排序錯亂(400 ×),在放大1 000倍和4 000倍下可見蝦肉肌纖維嚴重收縮,多處斷裂,被擠壓成團。相比之下,聯(lián)合干燥組蝦肉仍可見排列有序的肌纖維紋路(400 ×),在更大的放大倍數(shù)下也能看到部分排序錯亂(1 000 ×)、肌纖維溶脹(4 000 ×),但溶脹程度明顯小于CK組,整體紋路清晰,仍然保持原有排列方向。表明聯(lián)合干燥組的蝦肉肌纖維結(jié)構(gòu)破壞程度小于CK組。
2種處理方式的對蝦都是前期經(jīng)過90 ℃熱風干燥熟化2 h的,肌纖維蛋白變性收縮,水分由內(nèi)向表面擴散脫水,肌纖維間相互依靠,進而形成緊密的結(jié)構(gòu)[17]。持續(xù)高溫處理,肌纖維會進一步喪失完整性,發(fā)生斷裂,甚至溶解形成膠狀[18]。聯(lián)合干燥組采用的二段干燥方式是低溫熱泵干燥,雖然干燥時間偏長,但卻能有效避免持續(xù)高溫對肌纖維的進一步破壞作用。董志儉等[19]也得出了冷風干燥南美白對蝦的組織結(jié)構(gòu)破壞成程度明顯小于熱風干燥的。

a-400 ×;b-1 000 ×;c-4 000 ×圖1 干燥蝦肉電鏡掃描圖Fig.1 SEM chart of dried prawn
2.3 干燥方式對蝦肉質(zhì)構(gòu)的影響
蝦肉復水2 h后的質(zhì)構(gòu)評價如表2所示。2種干燥方式得到的蝦肉干復水后的剪切力、硬度和咀嚼性沒有顯著差異,但聯(lián)合干燥組蝦肉的內(nèi)聚性、彈性和膠著性顯著(P<0.05)高于CK組,分別高出21%、16%和42%。較高的內(nèi)聚性、彈性和膠著性表明對蝦肉質(zhì)保持較好。膠原蛋白變性、嚴重收縮以及肌原纖維的聚積、短縮會使得肌肉的持水性、恢復能力降低[20]。聯(lián)合干燥過程中,一段高溫熱風干燥熟制,二段低溫脫水,相比CK組的長時間高溫(60 ℃以上)處理,聯(lián)合干燥對蝦肉肌肉纖維組織破壞較小(圖1),復水后肌肉蛋白能保持一定的彈性和持水能力,表現(xiàn)為具有較好的質(zhì)構(gòu)特性。
表2 對蝦復水后的質(zhì)構(gòu)評價Table 2 Texture evaluation of prawn after rehydration
2.4 干燥過程的水分遷移分析
2.4.1 干燥過程蝦肉的水分遷移
圖2為利用低場核磁共振儀檢測蝦肉二段干燥過程的橫向弛豫時間變化,其反演圖大體上顯示了3個峰,T21、T22、T23分別代表蝦肉中的結(jié)合水、不易流動水和自由水。其中不易流動水含量和比例變化最明顯,整個干燥過程中以不易流動水減少為主。不易流動水是蝦肉的最主要水分存在狀態(tài),其含量變化及減少速度可反映干燥效果。因此,此處單獨列出不易流動水所在峰頂點和峰面積隨干燥時間的變化(表3)。
CK組的T22峰在二段干燥過程中變矮變寬,峰面積在干燥前期急速減小,后期減小速率相對緩慢。而聯(lián)合干燥組的T22峰隨時間的增加,峰面積減小速率相對緩慢,在二段干燥前10 h,T22峰面積減小的平均速率為每小時150信號幅度,面積減小速率相對一致。但同等峰面積情況下,相比CK組,聯(lián)合干燥組的峰頂點左移速度加快,即不易流動水的弛豫時間更小(表3),束縛力更大。這表明聯(lián)合干燥技術(shù)在去除蝦肉水分的同時,也在促使蝦肉部分不易流動水向結(jié)合水方向遷移,蝦肉對不易流動水的束縛力加強,所以熱泵干燥后期水分更難去除。如果與蛋白結(jié)合的水被強硬去除,蛋白也會發(fā)生變性。由于部分不易流動水向結(jié)合水遷移,與蛋白牢固結(jié)合,這部分水分得以保留,聯(lián)合干燥的蝦肉結(jié)構(gòu)破壞相對較少,復水后質(zhì)構(gòu)特性表現(xiàn)相對較好(表2)。而CK組不易流動水快速損失,肌肉組織快速脫水必然對肌纖維造成不可逆影響,復水后仍表現(xiàn)為肌肉斷層。

a-CK;b-聯(lián)合干燥圖2 蝦肉干燥過程中橫向弛豫時間T2反演圖Fig.2 Refutation graph of transverse relaxation timeT2 of prawn during the drying process

表3 蝦肉干燥過程的不易流動水所在峰的峰信息Table 3 T22 Peak information of prawn during
圖3為蝦肉干燥過程中的MRI圖,圖中的顏色深淺反映了質(zhì)子密度大小引起的弛豫時信號強弱差別[21],直觀地還原了干燥過程蝦肉內(nèi)部水分分布情況的變化。CK組的蝦肉在二段干燥3、4 h后,蝦肉內(nèi)部信號值仍然是很深的紅色,而四周是明顯的綠色,蝦肉從內(nèi)到外的信號值顏色區(qū)別很大,表明蝦肉表面或蝦肉內(nèi)部顏色較淺部位脫水嚴重,而此刻的中心部位水分含量仍較高,這種情況很容易造成成品表面焦殼,進而增加中心水分向外蒸發(fā)擴散的阻力[22]。相比之下,聯(lián)合干燥組的蝦肉信號值顏色變化較緩慢,蝦肉整體信號值顏色差別比CK組小,屬于溫和脫水。

a-CK;b-聯(lián)合干燥圖3 蝦肉干燥過程中MRI圖Fig.3 MRI images of prawn during the drying process
2.5 復水過程的水分遷移分析
2.5.1 蝦肉復水率分析
由圖4可知,0~15 min CK組蝦肉復水率幾乎呈直線式上升,15 min后復水率的增加有所減緩,30 min 后復水率的增加大大減緩,復水120 min后,復水率只有66.56%。聯(lián)合干燥組蝦肉的復水率在15 min后就開始大于CK組,30 min以后的復水率就遠超CK組,30~90 min的復水率增加速率仍然可以達到每15 min增加6%~10%。復水120 min,聯(lián)合干燥組的復水率高達91.79%。

圖4 蝦肉復水曲線Fig.4 Rehydration curve of prawn
復水率在一定程度上可反映干制品的品質(zhì)。干燥過程中產(chǎn)品的肌肉組織結(jié)構(gòu)破壞越小,復水效果就越好[23],干制品復水的口感就越接近新鮮樣品。相比CK組,聯(lián)合干燥組二段溫和干燥方式對蝦肉肌纖維結(jié)構(gòu)影響較小,蝦肉干復水率較高,復水后表現(xiàn)為持有較高的內(nèi)聚性、彈性和膠著性。肌纖維軸的橫向收縮主要發(fā)生在40~60 ℃,60~70 ℃時結(jié)締組織網(wǎng)絡(luò)和肌纖維縱向協(xié)同收縮[18, 24],這種收縮使得大量水分排出。高溫下蝦肉肌纖維蛋白質(zhì)疏水區(qū)域的局部結(jié)構(gòu)和蛋白質(zhì)的三級結(jié)構(gòu)都會發(fā)生變化,蛋白發(fā)生降解聚集[25],肌纖維結(jié)構(gòu)發(fā)生明顯的不可逆變化,失去持水性[26]。CK 組的二段干燥正處于60 ℃ ,因此復水后無法截留充足水分,復水率低下。而且長時間的高溫,蝦肉表面也容易形成硬殼,復水時阻礙水分快速滲透到內(nèi)部組織。
2.5.2 復水過程蝦肉的水分遷移
由橫向馳豫時間T2反演圖(圖5)可看出,圖譜中主要有3峰,從左到右分別代表結(jié)合水、不易流動水和自由水。從圖譜中可以看到,復水過程中,蝦肉的不易流動水峰面積在增大,峰整體在向右移。表明蝦肌肉組織在吸水。

a-CK;b-聯(lián)合干燥圖5 蝦肉復水過程中橫向弛豫時間T2反演圖Fig.5 Refutation graph of transverse relaxation time T2 of prawn during the rehydration process
干制品復水過程增加的主要是不易流動水和自由水,其中不易流動水對產(chǎn)品品質(zhì)影響最大。表4中可看到規(guī)律,復水前,聯(lián)合干燥的蝦肉不易流動水所在峰的峰頂點橫向弛豫時間T2比CK組小的,單位峰面積也更小。即復水前,相比CK組,聯(lián)合干燥組的不易流動水束縛力更大。但復水5 min后,聯(lián)合干燥組的蝦肉不易流動水所在峰的峰頂點橫向弛豫時間T2開始大于CK組,單位質(zhì)量峰面積接近CK組。復水15 min以上,聯(lián)合干燥組的蝦肉不易流動水所在峰的峰頂點橫向弛豫時間T2明顯大于CK組,平均大4ms,同時,單位峰面積也表現(xiàn)為明顯更大。這表明復水后,聯(lián)合干燥蝦肉這部分水的束縛力明顯更小,含量更高。即聯(lián)合干燥的蝦肉復水速率增加得更快,復水效果更佳。結(jié)果與2.5.1復水率分析結(jié)果一致。此外,復水后蝦肉的內(nèi)聚性、彈性和膠著性也顯著較好。程沙沙等[27]的研究也表明,蝦仁復水過程中LF-NMR參數(shù)與部分質(zhì)構(gòu)特性和復水率有極顯著的相關(guān)性。
2.5.3 復水過程中蝦肉MRI分析
圖6為蝦肉復水過程中的MRI圖。由圖6可知,隨著復水時間的延長,核磁共振信號值從邊緣往內(nèi)部增強,表明水分從外部逐漸滲透到內(nèi)部。復水5 min,CK組的蝦肉邊緣核磁共振信號值較高,復水較快,但后續(xù)復水過程中,聯(lián)合干燥蝦肉的整體信號值會比CK組的高,蝦肉內(nèi)部信號值覆蓋速度會快于CK組,表明聯(lián)合干燥的蝦肉內(nèi)部復水速度更快,復水效果更好。

表4 蝦肉復水過程的不易流動水所在峰的峰信息Table 4 T22 peak information of prawn during the rehydration process
復水120 min后,聯(lián)合干燥組整只蝦肉都覆蓋有核磁共振信號,邊緣信號值偏高(黃紅色),內(nèi)部信號值略低(綠色為主),但信號均勻,最中間部位信號值最低(淺藍色),表明聯(lián)合干燥的蝦肉120 min后大體復水完畢,蝦肉邊緣水分含量最高,內(nèi)部水分分配均勻,只是最中間有一點點部位還沒有完全復水。相比之下,CK組蝦肉復水75~120 min期間,蝦肉內(nèi)部有3個部位明顯是完全沒有信號值的(深藍色),猜測可能是這3個部位的蝦肉斷層。長時間的高溫熱風干燥使蝦肉變干變脆,肌肉纖維斷裂,彈性變差(表2),表現(xiàn)為蝦肉內(nèi)部組織斷層,復水時斷層部位沒有肌肉組織,所以沒有水分子信號表征。

圖6 蝦肉復水過程中MRI圖Fig.6 MRI images of prawn during the rehydration process
3 結(jié)論
本研究采用一段高溫熱風(熟化、上色、定型)、二段低溫熱泵(進一步脫水干燥)聯(lián)合干燥南美白對蝦,探究干燥過程及復水過程蝦肉的水分分布及遷移規(guī)律,結(jié)合微觀結(jié)構(gòu)分析、質(zhì)構(gòu)評價、復水率和MRI圖分析,并與分段熱風干燥作對比(CK組:一段90 ℃ 熱風干燥,二段60 ℃熱風干燥),闡明低溫熱泵干燥對蝦肉品質(zhì)保持的機理。結(jié)論如下:
(1)在干燥品最終水分含量一致的情況下,相比CK組,熱風-熱泵聯(lián)合干燥對蝦的TVB-N含量顯著降低43.50%;復水后表現(xiàn)出更佳的質(zhì)構(gòu)特性,內(nèi)聚性、彈性和膠著性分別高出21%、16%和42%;復水率和復水效果遠超CK組,2 h后的復水率高達91.79%。電鏡掃描結(jié)果顯示,聯(lián)合干燥組的蝦肉肌纖維結(jié)構(gòu)破壞程度較小。
(2)基于LF-NMR分析干燥過程中蝦肉的水分分布和遷移,干燥過程以不易流動水減少為主。二段干燥過程前10 h,聯(lián)合干燥的蝦肉不易流動水以每小時150信號幅度的平均速率緩慢減少,同時T22峰頂點左移速度加快,促使蝦肉部分不易流動水向結(jié)合水方向遷移,因此熱泵干燥后期水分更難去除。由MRI圖可見,CK組蝦肉二段干燥3~4 h后,蝦肉整體信號值顏色區(qū)別很大,容易造成表面焦殼,而聯(lián)合干燥的蝦肉整體信號值顏色差別較小,屬于溫和脫水。
(3)基于LF-NMR分析復水過程中蝦肉的水分分布和遷移,以不易流動水和自由水的增加為主,T22峰面積逐漸增大,峰整體右移。復水過程,聯(lián)合干燥的蝦肉T22峰頂點快速右移,15 min后其峰頂點弛豫時間和峰面積明顯大于CK組,進一步證實聯(lián)合干燥的蝦肉復水更快,復水效果更好。聯(lián)合干燥蝦肉復水過程的MRI圖信號值更高,蝦肉內(nèi)部信號值覆蓋速度更快,也證實了這一結(jié)論。而CK組的復水蝦肉MRI圖表明蝦肉內(nèi)部組織有3個部位斷層,與干燥過程對蝦肉肌纖維結(jié)構(gòu)的嚴重破壞有關(guān)。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
