酒酒球菌酸脅迫應答機制研究進展
2022-11-29 04:40:28劉龍祥張克穎趙紅玉宋煒鈺
食品與發酵工業 2022年22期
劉龍祥,張克穎,趙紅玉,宋煒鈺
1(濱州學院 生物與環境工程學院,山東省黃河三角洲野生植物資源開發利用工程技術研究中心,山東 濱州,256600) 2(西北農林科技大學 葡萄酒學院,陜西 楊凌,712100)
蘋果酸乳酸發酵(malolactic fermentation, MLF)可以將葡萄汁中尖銳的二元羧酸蘋果酸轉化為柔和的一元羧酸乳酸和二氧化碳,是葡萄酒釀造中一個重要的過程[1]。MLF的順利進行使葡萄酒的酸度降低,隨著碳源的消耗微生物穩定性得以提高,此外,MLF會改變葡萄酒的香氣結構,增加葡萄酒的果香味,豐富葡萄酒的結構感等。因此被認為是釀造優質紅葡萄酒必需的步驟[2]。
酒酒球菌(Oenococcusoeni)作為蘋果酸乳酸發酵的主要啟動者,其生長繁殖和蘋果酸乳酸發酵的進行都會受到葡萄酒多種生理生化特性的抑制[3]。葡萄酒中影響蘋果酸乳酸發酵進行的4種主要因素分別為乙醇(10%~16%,體積分數)、低pH(3.0~3.5)、SO2(超過10 mg/L)和低溫(可能低于12 ℃)[4]。在這些脅迫因素中,低pH是限制微生物在葡萄酒中生長的關鍵因素。目前許多研究都集中在酒酒球菌脅迫應答機制方面,提出了多種相關機制,例如膜成分和流動性、pH內外平衡、氧化應激反應、DNA和蛋白質損傷修復機制[5]。但酒酒球菌脅迫適應性的具體機制仍需進一步研究。本文主要從酒酒球菌酸脅迫應答機制及提高酒酒球菌抗脅迫能力的方法進行綜述。
1 酒酒球菌脅迫應答與防御機制
1.1 阻隔氫離子
在脅迫條件下微生物需要維持細胞膜的主要功能,以控制離子通透性,調節細胞與外界環境間的物質交換。酒酒球菌是革蘭氏陽性細菌,沒有完整的細胞結構,直接由細胞膜包被,所以細胞膜是其抵御不良環境的第一道屏障。酒酒球菌主要通過調節細胞膜的流動性和組成成分來阻隔外界脅迫環境中的氫離子。
pH作為影響微生物生長的重要因素之一,也是影響MLF順利進行的重要因素。在較高pH處理(pH 3.2)酸脅迫瞬時處理后,細胞膜軸向流動性發生微弱的增大,隨著培養時間的延長,細胞膜流動性會逐漸恢復[6]。較低pH處理(pH 3.0)后,會引發細胞膜軸向擴散速率降低,隨著pH的降低這種影響越顯著,當細胞培養環境pH恢復正常時,這種狀態不會恢復,這說明酸脅迫對細胞膜的影響是不可逆的[7-8]。
細胞質膜脂肪酸組成的修飾是微生物對逆境最重要的適應性反應之一,其中細菌的反應包括飽和度、碳鏈長度、分支位置、順式/反式異構化,以及不飽和脂肪酸轉化為環狀脂肪酸等[9]。pH 3.5和pH 3.2處理對細胞生物量的影響不大,但可以明顯提高細胞的接種活力和冷凍干燥活力,隨著pH的降低,不飽和脂肪酸(C18∶1)轉化為環狀脂肪酸(C19cyc11),短時間酸脅迫處理會顯著降低不飽和脂肪酸與飽和脂肪酸比值,但長時間的酸脅迫培養會增加細胞膜脂中不飽和脂肪酸與飽和脂肪酸的比值,這說明細胞在應對短時間與長時間酸脅迫處理時會采用不同的應對策略,短時間通過降低細胞膜流動性阻止胞外氫離子對細胞的侵害,隨著酸脅迫處理時間的延長,細胞通過調整胞內生化反應狀態,逐漸適應低pH侵害,細胞膜流動性恢復,加速胞內外物質交換[10-11]。
近期有研究表明,在脅迫環境中,酒酒球菌的細胞壁合成相關的基因的表達和蛋白含量也有顯著的變化,說明細胞壁可能同樣具有阻礙環境脅迫的屏障作用[12]。
1.2 排出氫離子
通常微生物調控胞內pH的方式有:(1)轉運質子的ATP酶;(2)胞內生成堿;(3)消耗胞內質子等[13-14]。其中,H+-ATP酶在維持胞內pH穩定方面起著最為重要的作用,它通過消耗ATP將胞內的H+泵出胞外。H+-ATP酶是酒酒球菌脅迫適應性機制研究的一個重要方面。研究發現,ATP酶的缺失會導致蘋果酸-乳酸操縱子也不發生轉錄,使細胞無法進行MLF[15],且在低pH條件下,H+-ATP酶活性會顯著提高[16],在酸脅迫處理1 h時,H+-ATP酶活性顯著低于對照組,在脅迫處理3 h后,酸脅迫處理組H+-ATP酶活性與對照組無顯著差異,說明在酸脅迫處理較短時間內,H+-ATP酶的活性可以恢復到正常水平[8],這一機制可以在短時間內通過消耗能量的方式將大量氫離子排出胞外,從而使細胞內pH穩定在較高水平,降低氫離子對細胞內酶與DNA等分子及生化反應的影響,維持細胞的正常功能,為細胞做出脅迫響應爭取時間。
與生理數據報道相比,關于轉運基因的報道要多一些。在O.oeni的酸脅迫轉錄組研究中發現了121個與ABC轉運體相關差異表達基因,說明ABC轉運體發揮了重要作用[4]。ATP酶基因在酸脅迫處理1 h和3 h時均發生上調表達,但在模擬酒環境中,該基因呈現先下降后上升的趨勢,在長時間模擬酒環境下,隨著蘋果酸的消耗,ATP酶基因表達逐漸被激活[12]。
1.3 中和氫離子
另一種抵抗酸脅迫的方法是細胞質堿化。在酒酒球菌中消耗氫離子的代謝途徑主要有氨基酸合成和代謝、蘋果酸乳酸發酵、檸檬酸代謝等途徑。蘋果酸乳酸發酵是酒酒球菌維持細胞內外pH穩定的重要途徑,蘋果酸乳酸發酵主要由蘋果酸滲透酶和蘋果酸乳酸酶完成,蘋果酸滲透酶負責將胞外的蘋果酸轉運進入細胞,二元羧酸蘋果酸在蘋果酸乳酸酶的催化作用下轉換成一元酸乳酸,這一過程可以降低胞外氫離子含量。在酸脅迫耐受酒酒球菌突變菌株中,發現在酸脅迫培養條件下,蘋果酸乳酸酶基因發生突變且呈現出高表達趨勢,說明該基因在酒酒球菌脅迫耐受性中的重要作用[17]。與培養條件下不同的是,酸脅迫瞬時處理時,蘋果酸乳酸酶基因發生下調,而蘋果酸脫氫酶基因顯著上調,使蘋果酸更多的轉化為丙酮酸進入三羧酸循環(tricarboxylic acid cycle,TCA),產生能量,而不是乳酸[8]。這與模擬酒和葡萄酒環境下(酸和乙醇脅迫同時存在)酒酒球菌的表現類似,在模擬酒瞬時脅迫處理8 h過程中,蘋果酸滲透酶和蘋果酸脫氫酶基因均上調表達[12],葡萄酒環境中,酒酒球菌也上調了蘋果酸滲透酶和蘋果酸脫氫酶基因表達量[18]。
檸檬酸裂解酶是檸檬酸代謝的關鍵酶,在檸檬酸裂解酶的催化作用下,檸檬酸裂解形成草酰乙酸、乙酰輔酶A和磷酸鹽,并產生ADP。檸檬酸代謝會誘導香氣揮發物的產生,如雙乙酰和乙氨酸。酸脅迫處理幾乎不影響檸檬酸裂解酶基因的表達量,而在乙醇脅迫、模擬酒和葡萄酒環境下,檸檬酸裂解酶基因會大幅度上調表達[4, 12, 19],說明檸檬酸途徑基因的表達主要受乙醇的影響,pH的影響較小[20]。
氨基酸作為葡萄酒風味成分的重要前體,其合成代謝不僅對葡萄酒香氣改善,而且對細胞生長繁殖具有重要作用。在酸脅迫(pH 3.0)處理1 h和3 h時,與精氨酸、亮氨酸、異亮氨酸、蛋氨酸、半胱氨酸、絲氨酸、賴氨酸生物合成關鍵酶均保持較高表達水平,精氨酸代謝途徑的3個關鍵酶(精氨酸脫亞胺酶、鳥氨酸轉氨甲酰化酶和氨基甲酸激酶)保持正常表達水平,纈氨酸、亮氨酸和異亮氨酸的降解途徑的關鍵酶上調表達,這些過程上調表達可以代謝產生ATP,供細胞利用[8, 14, 17]。
1.4 修復DNA和蛋白質損傷
酸脅迫對細胞帶來的損傷很大程度上是通過破壞或影響DNA和蛋白質活性產生的,細菌常見應對DNA和蛋白質損傷的機制是錯配修復和伴侶蛋白的合成。在酒酒球菌中已經發現19個與基因錯配修復相關的基因(表1),這些基因在酸脅迫瞬時處理條件下,大部分未表現出差異表達,少數基因發生下調表達[8],這些基因的正常表達可以降低在基因復制過程中出現堿基突變的概率。除了錯配修復機制外,分子伴侶蛋白可以促進脅迫環境下微生物蛋白質的正確合成,降低發生錯誤的幾率,從而增強微生物脅迫耐受性[21]。在酸脅迫瞬時處理條件下,O.oeniSD-2a大部分分子伴侶蛋白基因都在1 h時呈現出上調表達,且在3 h時這些基因的表達量均顯著增加,說明分子伴侶蛋白在應對酸脅迫瞬時處理時持續發揮著關鍵作用,且hsp20和groES在O.oeniSD-2a酸脅迫應答反應中的響應時間要早于其他分子伴侶蛋白(表2)。
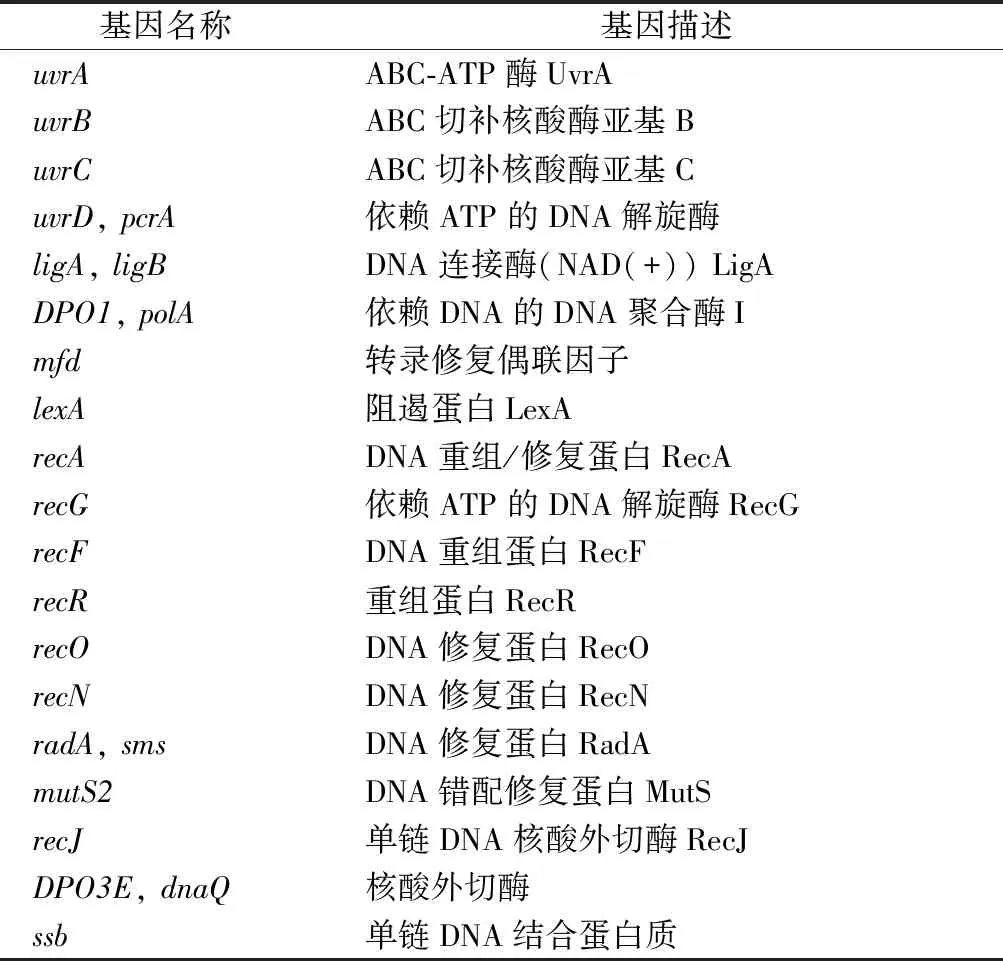
表1 與酒酒球菌DNA損傷修復相關的基因Table 1 Genes associated with DNA damage repair in O. oeni
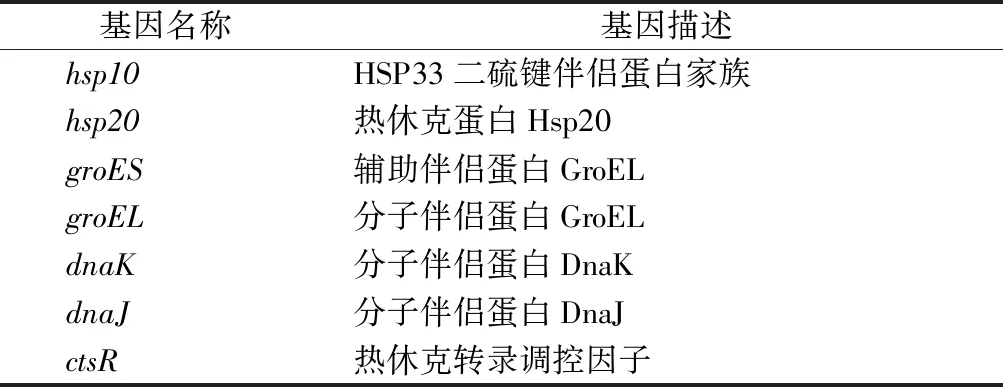
表2 酒酒球菌中分子伴侶蛋白基因Table 2 Genes encoded molecular chaperones in O. oeni
1.5 能量產生
在蘋果酸乳酸發酵過程中,酒酒球菌除了要抵御脅迫造成的損傷,還要從營養匱乏的葡萄酒生境中獲得能量。由于營養物質匱乏,酒酒球菌難以直接利用碳源產生能量,其獲得能量來源主要依靠細胞跨膜pH梯度(ΔpH)和電位差Δψ這兩種質子推動力,在質子推動力的作用下,細胞膜H+-ATP酶可以合成ATP[22]。細胞跨膜pH梯度在蘋果酸乳酸發酵早期增大,隨著蘋果酸的消耗,逐漸降低直至內外電位平衡,細胞內pH逐漸降低至等于培養基pH值。這說明ΔpH越大,O.oeni的蘋果酸乳酸發酵過程越短[22]。
此外,蘋果酸和檸檬酸代謝也與質子的消耗、跨膜電位和跨膜pH梯度的形成有關,因此可以促進胞內和胞外ATP的合成[20]。在酸脅迫瞬時處理過程中,ATP酶的4個亞基(α、β、γ和ε)基因均呈現上調表達,而糖酵解和TCA循環的關鍵基因均未出現差異表達,說明在酸脅迫條件下,酒酒球菌可以快速啟動蘋果酸乳酸發酵,其主要能量來源于跨膜質子推動力[4]。在模擬酒環境中,ATP酶基因亞基均呈現出下調表達,直至蘋果酸乳酸發酵開始才有所回升,說明在酸和乙醇脅迫同時存在的情況下,蘋果酸乳酸發酵的啟動時間會延遲[12]。
2 提高酒酒球菌抗脅迫能力的方法
2.1 預適應和交叉保護
預適應是將菌株暴露到致命或亞致死劑量的脅迫條件下一定時間,它可以強化菌株再次暴露于脅迫條件下的適應與恢復能力。有研究表明,低強度的脅迫預適應可以提高酒酒球菌在復合脅迫條件下的耐受性,且酒酒球菌存在著脅迫交叉保護現象,經酸脅迫預培養的菌體細胞接種到葡萄酒后,可以增強對其他脅迫的耐受性[23]。酸性條件下預培養可以提高酒酒球菌在高濃度SO2存在條件下或冷凍干燥后細胞的恢復與存活能力,且對SO2的脅迫應答會啟動pH內外穩定及脅迫應答蛋白合成等機制,以此消除脅迫帶來的影響[24-25]。除了酸脅迫預培養外,培養時加入低濃度乙醇(體積分數為10%)可以顯著提高細胞冷凍干燥后的存活率[26]。此外,在脅迫預培養階段添加谷胱甘肽,可以顯著提高細胞在模擬酒環境(乙醇體積分數12%,pH 3.4)下的生存能力,并且MLF速率比未添加谷胱甘肽預培養的菌株高[27]。
2.2 外源添加物
已有報道表明,通過外源添加一些物質,提高乳酸菌的抗脅迫能力。其中還原型谷胱甘肽(Reduced glutathione, GSH)可以促進O.oeniSD-2a在酸脅迫(pH 3.4)和乙醇脅迫(乙醇體積分數12%)環境中的生長能力,且不同的添加濃度可以刺激不同脅迫應答相關基因的表達, 顯著提高蘋果酸乳酸發酵速率[27]。MARGALEF-CATAL等[28]也有相似的發現,在培養時加入谷胱甘肽可以增加細胞膜環丙烷脂肪酸含量,從而提高酒酒球菌在高乙醇(體積分數為14%)和其他條件下(pH 4、pH 3.4和體積分數為6%的乙醇)的生存能力和生長速度。此外,在研究乳酸菌脅迫應答過程中,發現添加一些外源氨基酸可以提高乳酸菌酸脅迫耐受性,例如添加亮氨酸可以顯著提高菌體酸脅迫后的存活率[29],顯著提高了其酸脅迫耐受性。此外,也有添加谷氨酸、精氨酸、賴氨酸和瓜氨酸提高細菌酸脅迫耐受性的報道[30-32]。
2.3 菌種改良
菌種改良的方法主要分為基因重組法和非基因重組法2種,各有優缺點。基因重組技術通常具有較高的精度,通常側重于特定基因的添加或刪除,它的應用需要對菌株的遺傳背景與基因功能有清楚的認識。相比之下,非基因重組方法通常不需要事先了解某一性狀的遺傳基礎,但它的突變一般是隨機發生的,需要花費大量時間進行特定性狀突變株的篩選[33]。目前,已有成功在酒酒球菌中進行轉化實驗的報道[34-35],但由于轉化率較低,這些實驗尚未在其他實驗室完成重現。加之,政府及消費者對轉基因微生物直接應用于食品生產的限制,因此,酒酒球菌生產菌株改良主要采用非基因重組方法。
非基因重組方法,包括誘變、傳統菌株篩選法和定向進化等方法,這些方法不需要特定的遺傳知識,但需要經過繁瑣的篩選過程來選擇最佳突變菌株。紫外誘變技術已成功應用于葡萄酒釀造菌株篩選中,且從經過紫外照射處理的菌株中篩選出了蘋果酸乳酸發酵優良菌株[36]。盡管誘變操作容易實現,但其缺點是突變位點的隨機性,目標表型出現突變的概率較低。傳統菌株篩選法一般是從與葡萄酒生產相關環境中(葡萄表皮、葡萄汁、自然發酵葡萄酒等)分離和篩選酒酒球菌。許多具有應用前景的酒酒球菌菌株都是來自于傳統菌株篩選法[37-38]。定向進化可以直接利用脅迫條件下大量傳代培養,利用微生物自發有益突變在脅迫條件下的逐漸積累,獲得所需表型的優良突變菌株,但是,這一過程一般耗時較長。BETTERIDGE等[39]對酒酒球菌進行了乙醇脅迫下的定向進化研究,試驗獲得的突變菌株A90在20%(體積分數)的酒精存在情況,其存活數量比對照菌株提高了3個數量級。JIANG等[40]以A90為出發菌株,進行了酒精、二氧化硫和酸多重脅迫因素的定向進化實驗,經過350代的篩選,獲得了3株在極端模擬酒條件下發酵性能顯著提升的突變菌株。
3 展望
葡萄酒是由多種物理、化學脅迫因子組成的液體環境,這些脅迫因素的存在,常常造成蘋果酸乳酸發酵啟動的延遲甚至是失敗,因此越來越多的研究者將目光轉向酒酒球菌脅迫應答機制研究中,以期尋找高效方案解決蘋果酸乳酸發酵啟動穩定性的問題。隨著測序技術及質譜技術的發展,研究者利用基因組學、轉錄組學、蛋白質組學及代謝組學等技術,初步解析了酒酒球菌脅迫應答機制,但目前研究主要集中在酒酒球菌乙醇脅迫應答機制中,少有對酸、冷凍干燥、二氧化硫等脅迫因素的應答機制系統性的研究,需要進一步拓展。
本文通過總結已報道酒酒球菌酸脅迫應答研究,繪制酒酒球菌酸脅迫應答機制示意圖(圖1),酒酒球菌細胞暴露在酸脅迫環境中時,細胞通過改變細胞膜脂肪酸組分、降低細胞膜流動性和細胞膜損傷修復等阻隔氫離子向細胞內部流動,H+-ATP酶等氫離子轉運蛋白將流向胞內的氫離子轉運出細胞,蘋果酸、檸檬酸、氨基酸代謝等過程消耗胞內氫離子,產生ATP,ATP酶借助細胞跨膜pH梯度(ΔpH)和電位差Δψ產生能量,受損傷的DNA和蛋白質在錯配修復基因和伴侶蛋白作用下保持正常的復制過程與催化活性。
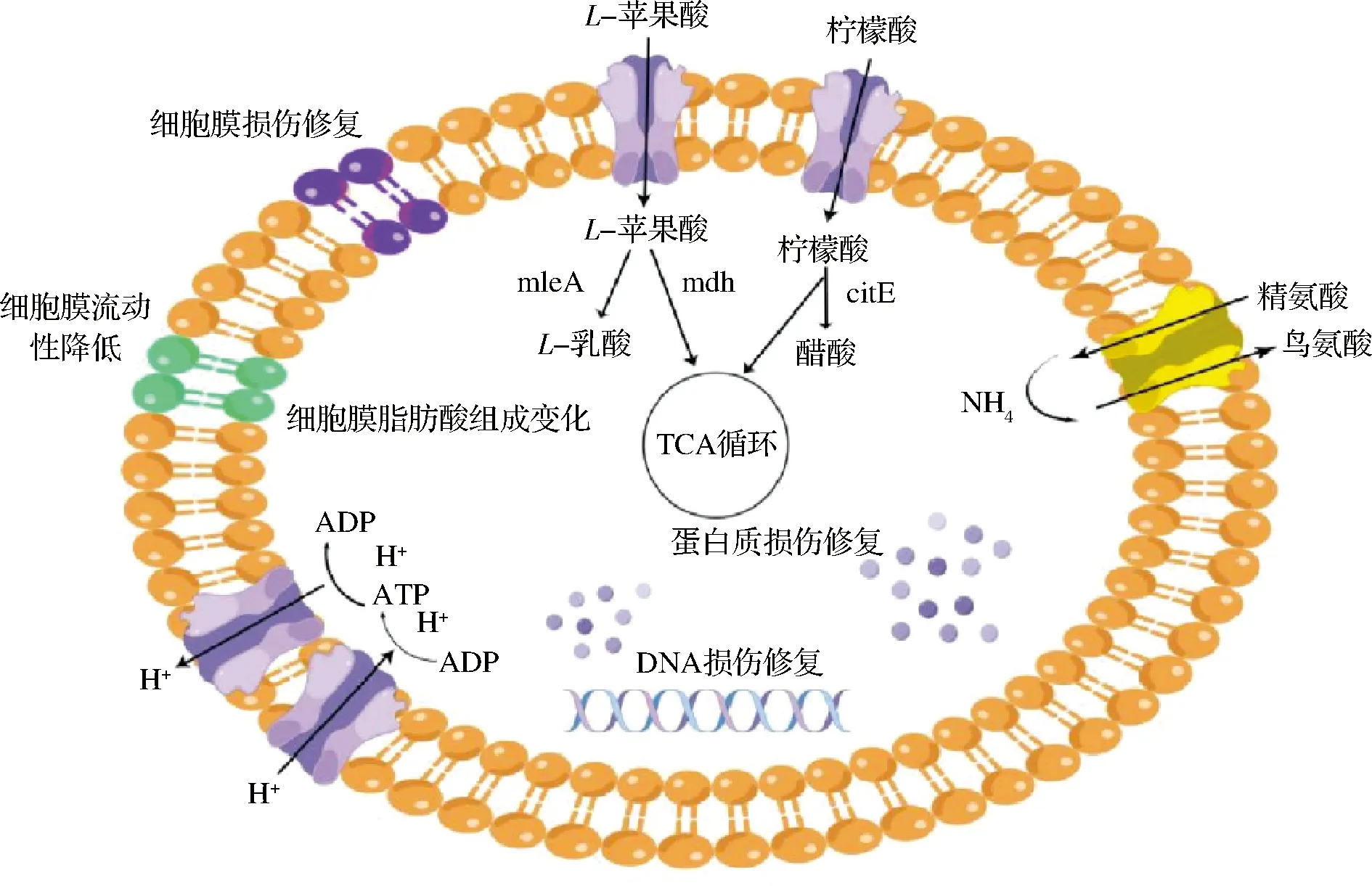
圖1 酒酒球菌酸脅迫應答機制示意圖Fig.1 The schematic diagram of acid stress response mechanism in Oenococcus oeni
雖然酒酒球菌酸脅迫應答機制已經取得一定的研究進展,但仍然缺乏系統性,多數研究者側重于利用組學技術發掘差異表達基因,但這些基因在其酸脅迫應答機制中的具體作用,酒酒球菌在酸脅迫條件下連續的生理、生化變化,酒酒球菌酸脅迫反應的啟動順序等方面,仍需要研究者進行系統性的研究。通過解析酒酒球菌酸脅迫應答機制,可以促進蘋果酸乳酸發酵啟動的穩定性,促進蘋果酸乳酸發酵菌劑的國產化與規模化,有利于我國葡萄酒產業的健康與可持續發展。
