常春藤皂苷元通過調控lncRNA PCAT19/miR-4319對TPC-1甲狀腺癌細胞生物學行為的影響
2022-12-02 14:39:02閆樂玉徐小波
中成藥 2022年7期
關鍵詞:檢測
鄭 奕, 閆樂玉, 陳 艷, 徐小波*
(1.黃淮學院醫學院,河南 駐馬店 463000;2.黃淮學院化學與制藥工程學院,河南 駐馬店 463000;3.黃淮學院建筑工程學院,河南 駐馬店 463000)
常春藤皂苷元是從藤梨根、威靈仙等植物中提取的一種齊墩果烷型單萜類化合物,具有抗腫瘤等作用,可抑制宮頸癌細胞增殖[1]。在喉癌細胞中,lncRNA PCAT19呈高表達,并可通過調節miR-182/PDK4促進細胞增殖[2]。靶基因預測軟件LncBase預測顯示,PCAT19與miR-4319有結合位點,并且后者在甲狀腺癌細胞中呈低表達,上調其表達可抑制甲狀腺癌細胞增殖及轉移[3],但PCAT19/miR-4319對甲狀腺癌細胞的調控機制尚不明確。因此,本研究探討常春藤皂苷元是否通過調控PCAT19/miR-4319表達影響甲狀腺癌細胞生物學行為,以期為甲狀腺癌治療藥物的研發提供新方向。
1 材料
1.1 細胞 甲狀腺癌TPC-1細胞,購自通派(上海)生物科技有限公司。
1.2 試劑與藥物 常春藤皂苷元(純度>98%)購自成都曼思特生物科技有限公司,溶于DMSO中備用。DMEM培養基、胎牛血清、MTT試劑、BeyoRTTMⅢ cDNA合成試劑盒、BeyoFastTMSYBR Green qPCR Mix均購自上海碧云天生物技術有限公司;Lipofectamine2000、Trizol試劑、pcDNA均購自美國Invitrogen公司;miR-4319模擬物(miR-4319 mimics)、mimic NC序列(miR-NC)、PCAT19干擾RNA(si-PCAT19)、陰性對照(si-NC)均購自廣州市銳博生物科技有限公司;PCAT19過表達(pcDNA-PCAT19)購自吉滿生物科技(上海)有限公司;Matrigel購自美國BD公司;Transwell購自美國Corning公司;兔抗人Twist、Snail、N-cadherin、E-cadherin抗體和二抗均購自美國Santa Cruz公司。
1.3 儀器 Nanodrop2000c超微量分光光度計、Multiskan Sky 酶標儀均購自美國Thermo Fisher公司;StepOnePlus實時熒光定量PCR儀購自美國ABI公司;CytoFLEX流式細胞儀購自美國貝克曼庫爾特公司;CO2培養箱購自濟南來寶醫療器械有限公司;電泳儀購自北京六一生物科技有限公司。
2 方法
2.1 細胞轉染、分組及給藥 取對數期TPC-1細胞接種于96孔板,用5、10、20 μg/mL常春藤皂苷元培養24 h[4],分別作為常春藤皂苷元低、中、高劑量組,將正常培養的細胞作為對照組。si-NC、si-PCAT19轉染TPC-1細胞,6 h后將培養基更換為含有10%胎牛血清的完全培養基培養24 h,分別作為si-NC組、si-PCAT19組。pcDNA、pcDNA-PCAT19分別轉染至TPC-1細胞,6 h后將培養基更換為含有20 μg/mL常春藤皂苷元的培養基培養24 h,分別作為pcDNA組、pcDNA-PCAT19組。
2.2 MTT檢測細胞增殖 各組TPC-1細胞接種于96孔板,每孔加20 μL MTT溶液培養4 h,棄上清,加150 μL DMSO,用酶標儀檢測490 nm波長處光密度(OD)值。
2.3 流式細胞術檢測細胞周期 各組TPC-1細胞制成單細胞懸液(1×104/mL),將細胞收集于流式管中,離心5 min后棄上清,加入300 μL含10%胎牛血清的培養基重懸細胞,再加入700 μL無水乙醇,置于-20 ℃冰箱中固定24 h,1 572×g離心30 s后棄上清,PBS洗滌后加入400 μL PI染色液染色10 min,采用流式細胞儀檢測細胞周期占比。
2.4 Transwell實驗檢測細胞遷移與侵襲能力 在上室中加40 μL Matrigel稀釋液,加入含有TPC-1細胞(1×104個)、不含血清的培養基200 μL,下室加500 μL含有胎牛血清的培養基,培養24 h后多聚甲醛固定30 min,結晶紫染色30 min,在顯微鏡下拍照并計數細胞。在遷移實驗操作過程中,小室的上室不加入Matrigel基質膠。
2.5 RT-qPCR檢測PCAT19、miR-4319 mRNA表達 采用TRIzol試劑提取各組TPC-1細胞總RNA,按照BeyoRTTMⅢ cDNA合成試劑盒說明書操作反轉錄合成cDNA,進行RT-qPCR反應,按照BeyoFastTMSYBR Green qPCR Mix 說明書配制反應體系,采用2-ΔΔCT法檢測目的基因相對表達。
2.6 雙熒光素酶報告基因檢測PCAT19、miR-4319的靶向關系 將PCAT19野生型/突變型(WT/MUT)克隆至pmirGLO載體中,分別將miR-NC、miR-4319 mimics、WT-PCAT19、MUT-PCAT19轉染至TPC-1細胞,24 h后用雙熒光素酶活性檢測試劑盒檢測熒光素酶活性。脂質體轉染技術將pcDNA、pcDNA-PCAT19、si-NC、si-PCAT19轉染至TPC-1細胞,24 h后用RT-qPCR法檢測miR-4319的表達。
2.7 Western blot檢測Twist、Snail、N-cadherin、E-cadherin蛋白表達 收集各組TPC-1細胞,加入裂解液裂解細胞,100 ℃水浴10 min,13 700×g離心10 min,取上清,即得總蛋白,檢測其濃度。配制12%分離膠與濃縮膠,電泳分離蛋白,轉膜,封閉2 h,加入Twist(1∶1 000)、Snail(1∶1 000)、N-cadherin(1∶1 000)、E-cadherin(1∶1 000)一抗和內參GAPDH抗體(1∶2 000)稀釋液,在4 ℃下孵育過夜,加二抗稀釋液(1∶10 000)室溫孵育1 h,采用Image J軟件分析各條帶灰度值。
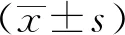
3 結果
3.1 常春藤皂苷元對TPC-1細胞增殖及周期的影響 與對照組比較,常春藤皂苷元各劑量組細胞存活率、S期細胞比例降低(P<0.05),G0/G1期細胞比例升高(P<0.05),并呈劑量依賴性,見表1。

表1 常春藤皂苷元對TPC-1細胞增殖及細胞周期的影響
3.2 常春藤皂苷元對TPC-1細胞遷移及侵襲能力的影響 與對照組比較,常春藤皂苷元各劑量組遷移及侵襲細胞數減少(P<0.05),Twist、Snail、N-cadherin蛋白表達降低(P<0.05),E-cadherin蛋白表達升高(P<0.05),并呈劑量依賴性,見圖1、表2。
3.3 常春藤皂苷元對TPC-1細胞中lncRNAPCAT19、miR-4319表達的影響 與對照組比較,常春藤皂苷元各劑量組PCAT19表達降低(P<0.05),miR-4319表達升高(P<0.05),并呈劑量依賴性,見表3。
3.4PCAT19與miR-4319的靶向關系 LncBase預測顯示,PCAT19與miR-4319存在結合位點,見圖2A。與miR-NC組比較,過表達miR-4319降低WT-PCAT19熒光素酶活性(P<0.05),見圖2B。PCAT19可負向調控miR-4319表達,見圖2C。
表2 常春藤皂苷元對TPC-1細胞遷移及侵襲的影響
表3 常春藤皂苷元對TPC-1細胞中lncRNA PCAT19、miR-4319表達的影響
3.5 抑制PCAT19表達對TPC-1細胞增殖、細胞周期、遷移及侵襲能力的影響 與si-NC組比較,si-PCAT19組細胞存活率、S期細胞比例、遷移及侵襲細胞數降低(P<0.05),G0/G1期細胞比例、E-cadherin蛋白表達升高(P<0.05),Twist、Snail、N-cadherin蛋白表達降低(P<0.05);2組間G2/M期細胞比例比較,差異無統計學意義(P>0.05),見圖3、表4。
3.6 上調PCAT19表達可逆轉常春藤皂苷元對TPC-1細胞增殖、細胞周期、遷移及侵襲的作用 與pcDNA組比較,pcDNA-PCAT19組miR-4319表達降低(P<0.05),細胞存活率升高(P<0.05),G0/G1期細胞比例降低(P<0.05),S期細胞比例升高(P<0.05),遷移及侵襲細胞數增多(P<0.05),Twist、Snail、N-cadherin蛋白表達升高(P<0.05),E-cadherin蛋白表達降低(P<0.05),見圖4、表5。
表4 抑制PCAT19表達對TPC-1細胞增殖、細胞周期、遷移及侵襲的影響
表5 上調PCAT19表達逆轉常春藤皂苷元對TPC-1細胞增殖、細胞周期、遷移及侵襲的作用
4 討論
甲狀腺癌細胞的高度轉移能力導致患者預后較差,且患者生存期縮短,越來越多的研究表明部分中藥可減緩甲狀腺癌的發展進程,其可通過多途徑或調控多個靶點而達到治療目的[5-6]。lncRNA在甲狀腺癌中表達上調或下調,并可能發揮癌基因或抑癌基因作用,當癌基因活化或抑癌基因失活時均可能促進甲狀腺癌的發生發展,通過調控lncRNA表達可逆轉甲狀腺癌細胞生物學行為進而抑制其發生發展[7-8]。
常春藤皂苷元可抑制乳腺癌細胞增殖、侵襲及誘導細胞凋亡[9]。常春藤皂苷元可能通過抑制PI3K/Akt信號通路,從而抑制前列腺癌細胞增殖、遷移及侵襲,并可促進細胞凋亡[10]。常春藤皂苷元還可通過抑制順鉑耐藥性頭頸癌細胞凋亡,從而增強其對順鉑的敏感性[11]。但常春藤皂苷元對甲狀腺癌的研究尚未見報道。本研究發現,常春藤皂苷元以劑量依賴性地降低甲狀腺癌細胞存活率,并可誘導G0/G1期細胞比例升高,而降低S期細胞比例,提示常春藤皂苷元可通過誘導細胞周期阻滯于G0/G1期而抑制甲狀腺癌細胞增殖。轉錄因子Twist、Snail與細胞遷移調控相關,是調控上皮-間質轉化(EMT)的重要轉錄因子,EMT主要是指在某些條件下,具有極性的上皮細胞失去極性轉化成間充質細胞,從而促使遷移及侵襲發生,Twist、Snail、N-cadherin表達增加,E-cadherin表達降低可促進EMT發生及腫瘤轉移[12]。本研究結果顯示,常春藤皂苷元處理后甲狀腺癌遷移及侵襲細胞數減少,Twist、Snail、N-cadherin蛋白表達降低,E-cadherin蛋白表達升高,提示常春藤皂苷元可抑制甲狀腺癌細胞遷移及侵襲,但常春藤皂苷元調控甲狀腺癌細胞生物學行為的分子機制尚需進一步探究。
本研究發現,不同劑量常春藤皂苷元處理后甲狀腺癌細胞中PCAT19表達降低,miR-4319表達升高。研究表明抑制PCAT19表達可抑制神經膠質瘤細胞增殖及侵襲[13],沉默PCAT19可抑制非小細胞肺癌細胞增殖[14]。本研究結果顯示,抑制PCAT19表達可降低甲狀腺癌細胞存活率,誘導細胞周期阻滯于G0/G1期,還可抑制細胞遷移及侵襲,提示PCAT19高表達可通過調控細胞生物學行為而促進甲狀腺癌的發生發展。本研究進一步證實PCAT19可靶向結合miR-4319。研究表明miR-4319可通過靶向ABTB1而抑制結直腸癌細胞增殖及誘導細胞周期阻滯[15]。在食管鱗狀細胞癌細胞[16]、乳腺癌[17]中miR-4319表達下調,上調miR-4319表達能抑制細胞增殖、轉移。本研究顯示,上調PCAT19表達可通過抑制miR-4319表達而逆轉常春藤皂苷元對甲狀腺癌細胞生物學行為的作用,提示PCAT19/miR-4319可能介導常春藤皂苷元調控甲狀腺癌細胞生物學行為過程。
綜上所述,常春藤皂苷元可誘導細胞周期阻滯于G0/G1期,抑制甲狀腺癌細胞增殖、遷移、侵襲,其作用機制與抑制PCAT19表達及促進miR-4319表達有關。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
