中街山列島海域曼氏無針烏賊棲息地偏向性研究
2022-12-12 11:41:40周永東劉姝含胡揚杰徐開達
浙江海洋大學學報(自然科學版) 2022年5期
吳 天,梁 君,周永東,劉姝含,胡揚杰,趙 瑞,徐開達
(浙江海洋大學海洋與漁業研究所,浙江省海洋水產研究所,浙江省海洋漁業資源可持續利用技術研究重點實驗室,浙江舟山 316021)
曼氏無針烏賊Sepiella maindroni俗稱墨魚,是分布廣泛的淺海種類,隸屬軟體動物門Mollusca、頭足綱Cephalopoda、烏賊目Sepiida、烏賊科Sepiidae、無針烏賊屬Sepiella,主要分布在日本海、馬來群島海域和印度洋東海岸,在我國東海和黃海均有分布,其中心分布區在浙江近海和閩東海域。歷史上東海區曼氏無針烏賊最高年產超過7×104t,曾與大黃魚Larimichthys crocea、小黃魚Larimichthys polyactis、帶魚Trichiurus lepturus并稱為我國四大海產[1-3]。但是20 世紀80 年代以來,由于過度捕撈和對資源的不合理利用,加之全球變暖造成棲息地退化,使得曼氏無針烏賊資源出現嚴重衰退現象,資源變動較大[4]。中街山列島海域位于舟山漁場的中心區域,受沿岸水與外洋水的影響,水體交換快、餌料充足,是我國主要經濟魚類生長、索餌、繁殖的重要場所,歷史上素有“魚倉”之美譽,曾是曼氏無針烏賊的重要產卵場之一[5]。
棲息地是指生物個體和種群居住的場所,是由生物出現在環境中的空間范圍和環境條件綜合構成,棲息地的選擇是指生物為了達到生存目的而尋找相對適宜棲息地的過程[6]。棲息地的研究一直以來都是動物生態學的研究重點,當棲息地質量下降時,會影響物種的數量,甚至導致物種滅絕[7]。在漁業資源研究中,棲息地的研究內容主要是生物棲息環境的變化對生物活動的影響,生物對棲息地的選擇是受多種環境因子共同作用的結果,單因子分析并不能很好地解釋生物對棲息地的選擇機制,而廣義可加模型(GAM)能夠直接處理響應變量與解釋變量之間的非線性關系,通過非參數的方法檢驗數據結構[8-9]。本研究基于2018-2021 年中街山列島海域漁業調查數據和環境數據,通過GAM 構建非參數的擬合模型,探究中街山列島海域曼氏無針烏賊資源分布與環境因子之間的關系,了解曼氏無針烏賊的棲息地選擇機制,旨在為曼氏無針烏賊的種群保護及產卵場生物多樣性保護提供理論基礎,為浙江漁場修復振興計劃的實施提供科學依據。
1 材料與方法
1.1 研究區域和站位布設
于2018-2021 年春季(4 月)和秋季(10 月)在中街山列島海域進行了8 個航次的底拖網漁業資源調查,調查范圍如圖1 所示。本研究中站位設置采取區域隨機分布的原則,沿潮流主軸(西北-東南)共設置了16 個站位。

圖1 調查站位示意圖Fig.1 Sketch of survey station
1.2 采樣方法和樣品處理
調查船為58.8 kW 單拖漁船,拖速為2 kn,拖曳時間為30 min,調查網具網口周長25 m,囊網網目尺寸3.0 cm。在每一次調查時,使用溫鹽深測定儀(CTD)同步采集環境數據,并在各站位采集水樣。共測定水深、表層海水水溫、表層海水鹽度、底層海水水溫、底層海水鹽度、葉綠素a、溶解氧、懸浮物、無機氮和活性磷酸鹽10 類環境數據。樣品采集、處理和分析等均按照《海洋漁業資源調查規范》[10]的規定執行。
1.3 數據處理和方法分析
1.3.1 資源密度和生物量估算
曼氏無針烏賊的資源密度估算和生物量估算按照以下公式[11]計算:

式中:D 為現存資源密度,ind.·km-2;C 為每小時取樣面積內的漁獲量,kg;q 為網具漁獲率,由于曼氏無針烏賊屬于沿岸暖水底棲種,q 取0.5;A 為網具每小時掃海面積,km2;B 為生物量,kg·km-2;為個體平均質量,kg·ind-2。
1.3.2 環境因子的相關性分析
在建模中,需要對環境因子進行相關性分析,克服共線性較高的解釋變量對參數預估準確性的干擾,當2 個環境因子間存在共線關系時,保留1 個變量作為解釋變量[12]。本研究采用Pearson 相關性分析對于環境因子之間可能存在共線性關系的環境因子進行相關性分析,并通過方差膨脹因子(variance inflation factors,VIF)對所有候選變量進行檢驗。
1.3.3 廣義可加模型(GAM)分析
本研究以曼氏無針烏賊資源分布為響應變量,其中季節作為分類變量,各環境因子為解釋變量。由于曼氏無針烏賊的零值站位出現的次數較多,假設曼氏無針烏賊分布模式為二項分布,GAM 模型選擇logistic 為鏈接函數模擬中街山列島海域曼氏無針烏賊資源分布同環境因子的關系[12-13]。GAM 的表達式為:

式中:P 為曼氏無針烏賊出現的概率;α 表示截距項;Si 表示樣條平滑函數;xi表示第i 個解釋變量;ε 表示誤差項。
GAM 模型是在加性模型基礎上提出的非參數擬合模型,能夠直接處理響應變量與多個解釋變量之間的非線性關系。在模型的擬合過程中,首先確定曼氏無針烏賊資源分布與單一環境因子的關系,通過逐步回歸法將篩選后環境因子加入模型中,然后利用赤池信息準則(akaike information criterion,AIC)逐步檢驗加入不同環境因子后的模型擬合度,通過F 檢驗來評估各因子的顯著性,以檢驗模型中每個環境因子的重要性,最終遴選出最優模型[13-14]。
數據通過Excel 軟件計算和整理,曼氏無針烏賊資源分布圖采用ArcGIS 10.2 軟件繪制,Pearson 相關性分析和VIF 檢驗通過SPSS 23.0 實現,GAM 模型構建與檢驗及可視化分析通過R 4.2.0 實現。
2 結果
2.1 曼氏無針烏賊時空分布特征
曼氏無針烏賊資源空間分布季節變化明顯。春季,曼氏無針烏賊主要分布于島礁外側,東北部居多(圖2)。秋季,曼氏無針烏賊主要分布于島礁內的東南和西南方向(圖3)。時間因子月份對曼氏無針烏賊出現概率有明顯影響,2018-2021 年曼氏無針烏賊的出現概率秋季明顯高于春季。但春季曼氏無針烏賊的體質量高于秋季(圖4)。
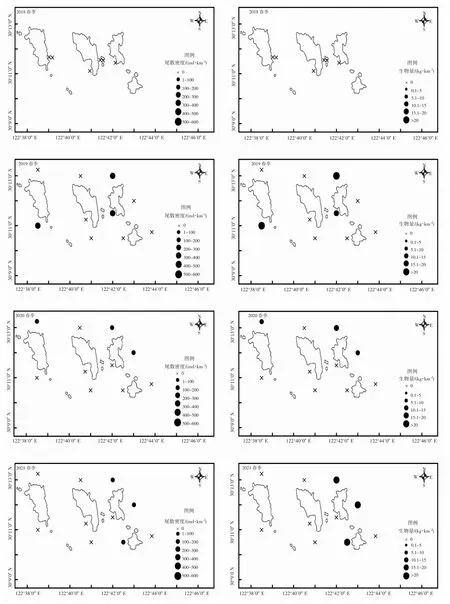
圖2 2018-2021 春季曼氏無針烏賊尾數密度和生物量空間分布圖Fig.2 2018-2021 spring distribution of S.maindroni tail density and biomass
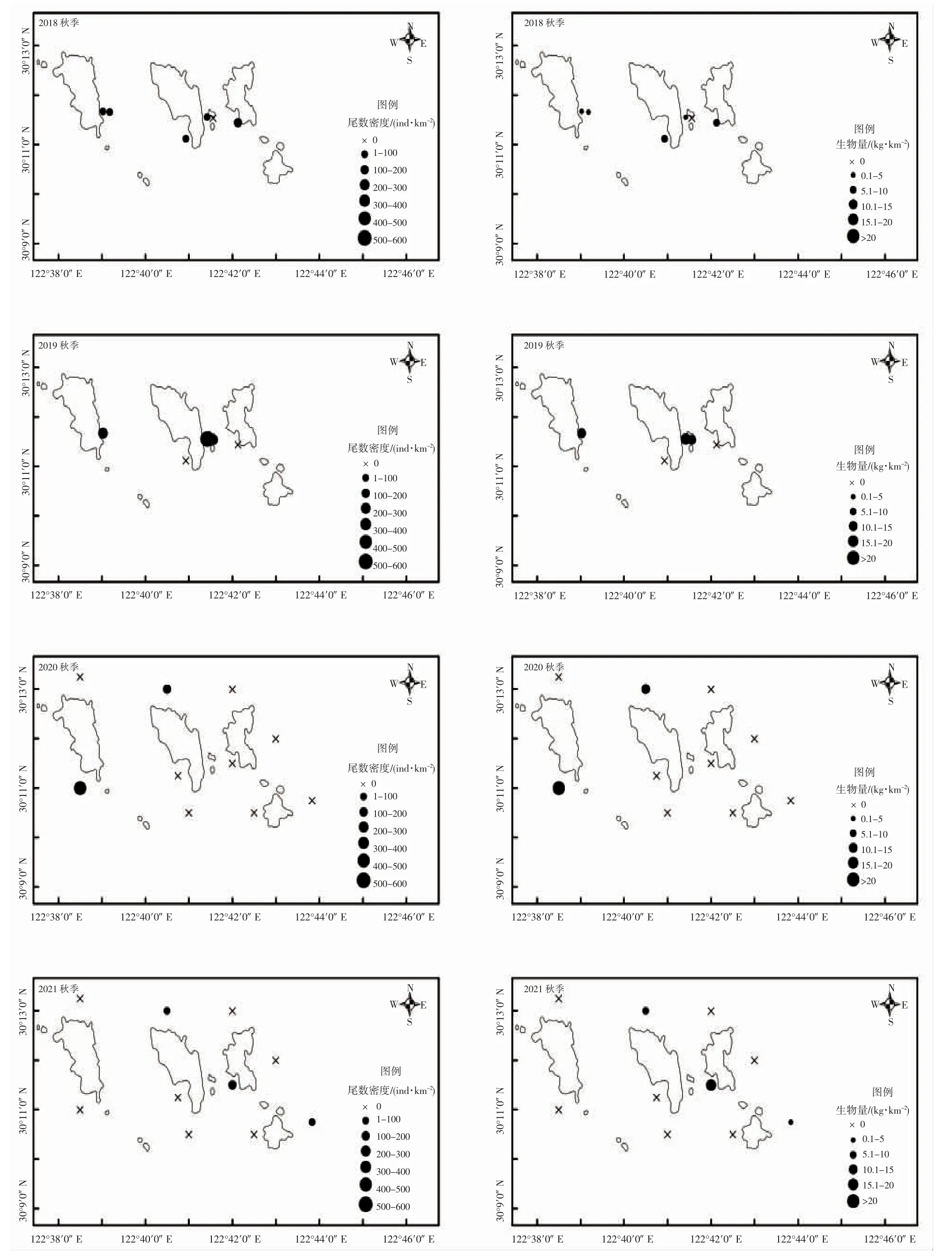
圖3 2018-2021 秋季曼氏無針烏賊尾數密度和生物量空間分布圖Fig.3 2018-2021 autumn distribution of S.maindroni tail density and biomass
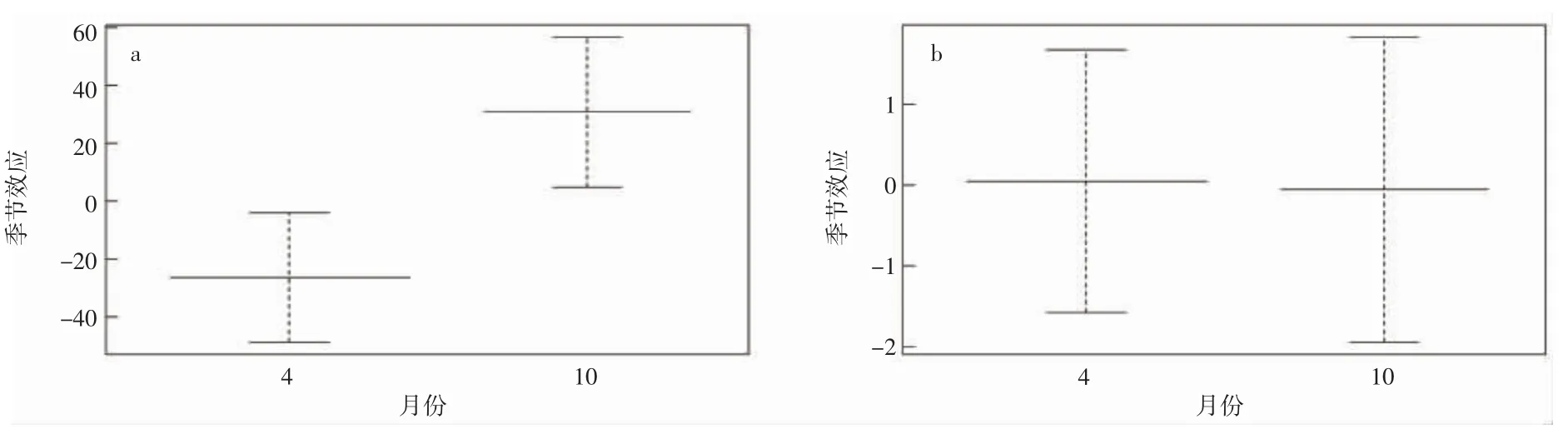
圖4 2018-2021 曼氏無針烏賊尾數密度和生物量對照圖(a.尾數密度;b.生物量)Fig.4 2018-2021 S.maindroni comparison of tail density and biomass(a.tail density;b.biomass)
2.2 關鍵環境因子識別
對于海洋生物,考慮的主要環境因子有溫度、鹽度、海表高度和葉綠素a等[5]。依據Pearson 相關性分析以及可視化結果(圖5)顯示,春季表層水溫和底層水溫、表層水溫和底層鹽度、底層水溫和底層鹽度、底層鹽度和活性磷酸鹽、葉綠素a和活性磷酸鹽存在顯著相關(P<0.05),秋季表層水溫、表層鹽度、底層水溫、底層鹽度之間均存在顯著相關,無機氮和活性磷酸鹽之間存在顯著相關,同時對各解釋變量進行多重線性檢驗,方差膨脹系數閾值設定為4,去掉相關性和方差膨脹系數均高于閾值的變量,因此本文選取水深、表層水溫、表層鹽度、懸浮物、葉綠素a、溶解氧和無機氮作為春季曼氏無針烏賊棲息地模型構建的解釋變量;水深、懸浮物、底層水溫、葉綠素a、溶解氧和活性磷酸鹽作為秋季曼氏無針烏賊棲息地模型構建的解釋變量。

圖5 春季和秋季不同環境因子相關性分析熱圖(a 為春季;b 為秋季)Fig.5 Correlation analysis of different environmental factors in spring and autumn(a.spring;b.autumn)
2.3 GAM 模型構建
在GAM 模型中加入篩選后的環境因子,依據AIC 最小原則構建曼氏無針烏賊棲息地選擇的最優模型。結果如下:
春季曼氏無針烏賊棲息地選擇的最優模型:

秋季曼氏無針烏賊棲息地選擇的最優模型:

式中:P為曼氏無針烏賊出現的概率;Depth 為水深,m;Chl-a為葉綠素a,μg·L-1;DO 為溶解氧,mg·L-1;ε 為相應的GAM 模型截距(表1)。
春季(GAM1)模型最佳預測變量組合為Depth+Chl-a(表1),模型累計偏差解釋率為70.35%,水深和葉綠素a 對曼氏無針烏賊的出現概率具有顯著影響(P<0.05),其貢獻率分別為23.94%和46.41%(表2)。秋季(GAM2)模型最佳預測變量為DO(表1),模型累計偏差解釋率為24.93%(表2)。

表1 GAM 模型變量篩選及影響因子的參數分析Tab.1 Variable screening process for GAM and parameters analysis

表2 模型擬合結果及各因子參數Tab.2 Result from models fitting and parameters of each factor
模型結果顯示春季(GAM1)水深、葉綠素a 是影響曼氏無針烏賊棲息地選擇的主要因素。春季曼氏無針烏賊主要出現的水深范圍在25~35 m 之間,呈先上升后下降的趨勢,30 m 時曼氏無針烏賊出現的概率最高(圖6a)。葉綠素a 在0.5~1 μg·L-1時曼氏無針烏賊出現概率為上升趨勢,當葉綠素a 超過1 μg·L-1時曼氏無針烏賊出現的概率開始下降(圖6b)。秋季,影響曼氏無針烏賊出現概率的單一環境因子為溶解氧,范圍在5.6~6.2 mg·L-1之間,呈先上升后下降的趨勢(圖6c)。
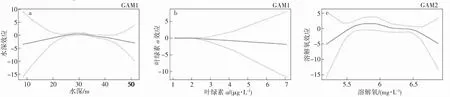
圖6 GAM1 和GAM2 環境因子對曼氏無針烏賊分布的影響Fig.6 Effects of GAM1 and GAM2 of environmental factors on the S.maindroni
3 討論
3.1 曼氏無針烏賊的時空分布變化
中街山列島海域海洋性氣候明顯,春季在季風的影響下,東海表層水進入浙江近岸,與陸地徑流(長江、錢塘江)交匯,形成鋒區,漁場近岸水溫較高,向東部水溫逐步遞減,近岸水與外側水鹽度差距較大。秋季季風轉換,偏北風頻率的增加,漁場內受冷暖水團交匯影響較強,由近岸往東水溫逐步升高,鹽度自西向東逐步增大[15]。曼氏無針烏賊屬沿岸暖水底棲種,生命周期僅有1a,春季由外海低溫高鹽的越冬場游向近岸島礁附近高溫低鹽的產卵場繁殖產卵,秋季隨季風轉換,開始向外海高鹽海域移動,進行索餌、越冬洄游[16-19]。研究結果顯示,春季(4 月)曼氏無針烏賊主要分布于島礁外側的東北部,秋季(10 月)曼氏無針烏賊主要分布于島礁內部偏南方向,這與倪正雅等[17]和李成林等[20]的研究結果相一致。故不同季節曼氏無針烏賊的時空分布與海水溫度、海水鹽度和洄游路徑有關。
本研究中,秋季曼氏無針烏賊的尾數密度明顯大于春季,但春季的生物量卻遠高于秋季。春季為曼氏無針烏賊的繁殖期,主要為個體較大的曼氏無針烏賊產卵親體,而秋季為曼氏無針烏賊的補充群體,故春季烏賊的體質量要高于秋季。而造成尾數密度秋季大于春季的原因可能與增殖放流和海洋保護區的資源養護作用有關。從2013 年開始,在中街山列島海洋保護區內實施了大規模曼氏無針烏賊增殖放流,累計放流受精卵達7 835×104ind.,一定程度上增加了曼氏無針烏賊的補充群體[21]。中街山列島國家級海洋特別保護區的設立有利于海洋生物的資源保護和修復,烏賊增殖礁的投放,也有利于自然海域曼氏無針烏賊受精卵的附著,提高了曼氏無針烏賊的附卵量,增加補充群體[22]。
3.2 棲息地選擇的關鍵影響因子
不同生活史階段對于生境的需求也有所不同,模型擬合結果顯示春季曼氏無針烏賊的分布受水深和葉綠素a2 個關鍵因子的影響,秋季曼氏無針烏賊的分布受單一因子溶解氧的影響。
曼氏無針烏賊主要分布于近海淺水海域,其產卵場位于島礁周圍50 m 范圍內,在繁殖期時從60~80 m的越冬場游向10~40 m 的產卵場作生殖洄游[3]。李星頡等[23]的研究表明,中街山列島海域曼氏無針烏賊的附卵水深范圍在10~20 m。本研究通過GAM 模型顯示春季曼氏無針烏賊會選擇水深范圍在25~35 m 的海域作為產卵場和育幼場,在水深為30 m 時,曼氏無針烏賊的資源密度達到最高值,原因可能是本研究基于二項分布的GAM 模型來表達曼氏無針烏賊的出現概率,從已知數據預測整體分布特點,推斷曼氏無針烏賊出現理論分布區,而前人研究以產卵附著基(柳珊瑚)上受精卵的附著率為調查內容,故本研究與前人研究結果略有差異。
葉綠素a的濃度值反映了海域初級生產者的現存量,能直接反映海區的肥癟程度,是養育生物資源的直接指標,也能間接反映海區初級生產力的高低[24-26]。葉綠素a作為浮游植物豐富度的重要指標,能為浮游生物提供有利的生長條件[27]。葉綠素a濃度與頭足類的漁場分布有一定關聯性,如劍尖槍烏賊Uroteuthis edulis、神戶烏賊Sepia kobiensis、金烏賊Sepia esculenta、長蛸Octopus variabilis等漁場分布均受到葉綠素a濃度值的影響[28]。根據GAM 模型分析的結果,葉綠素a對曼氏無針烏賊的分布影響較大,隨著葉綠素a濃度的增加曼氏無針烏賊的資源密度也呈上升趨勢,浮游生物的增多,可以為烏賊提供充足的餌料[29]。成體曼氏無針烏賊主要以小型魚類、甲殼動物和少量軟體動物為主,幼烏賊主要以浮游動物,如箭蟲、橈足類等,特別是夜光藻可作為烏賊漁汛的直觀指標[23,30]。
海水中充裕的溶解氧是維持海洋異養生物生命活動、避免水體有機物發生厭氧分解而散發臭味的必要條件,故海水中溶解氧被作為保障海洋生物生存的水化學指標和水體的感官指標[31]。李星頡等[23]對中街山列島海域曼氏無針烏賊產卵區的生態環境進行調查,結果顯示4-6 月自然海域曼氏無針烏賊產卵場的溶解氧為7.0~8.8 mg·L-1。蔣霞敏等[32]和劉振勇等[33]的研究表明曼氏無針烏賊室內孵化期的溶解氧為4.3~4.8 mg·L-1,在養殖過程中溶解氧應維持在5.0 mg·L-1以上。頭足類的所有活動均需要消耗氧氣,且曼氏無針烏賊的耗氧率與水溫成正比例關系,個體體重成反比例關系,在低溶解氧的狀態下,會造成曼氏無針烏賊生長緩慢,使其新陳代謝率降低,影響個體活動[34-35]。本研究結果顯示,春季中街山列島海域平均溶解氧為7.0 mg·L-1,這與李星頡等[23]的研究結果一致。模型擬合結果顯示秋季曼氏無針烏賊偏向棲息于溶解氧濃度在5.5~6.5 mg·L-1之間的海域,同春季相比產卵區溶氧含量有所降低,蔣國昌等[36]的研究結果表明,東海區海水氧含量季節變化明顯,春季含量最高,秋季含量最低。故季節變化同產卵區溶解氧濃度變化有明顯關系。此外,近年來曼氏無針烏賊小型化明顯,人工養殖的曼氏無針烏賊個體普遍偏小,生長速度快,壽命周期為4 個月左右,且人工養殖環境中溶氧可調控,故自然海域和養殖環境下曼氏無針烏賊所需溶氧有所不同[37]。
在構建曼氏無針烏賊棲息偏向性模型時,除水深、水溫、鹽度、葉綠素a、溶解氧等環境因子外,餌料生物與捕食者、天然附著基柳珊瑚與海藻、底質類型、流速等均可對曼氏無針烏賊棲息地的選擇產生影響。因此,今后還需考慮生物與非生物因素共同影響下曼氏無針烏賊棲息地的選擇,以期進一步提高模型的預測能力,全面客觀地分析曼氏無針烏賊棲息地選擇偏好與分布特征。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
光學精密工程(2016年6期)2016-11-07 09:07:19
