氣候變化情景下中國(guó)外來(lái)入侵植物黃頂菊潛在分布區(qū)模擬與早期預(yù)警
2022-12-20 06:22:44柳曉燕趙彩云李俊生李飛飛朱金方
環(huán)境科學(xué)研究 2022年12期
柳曉燕,趙彩云*,李俊生,李飛飛,朱金方
1.中國(guó)環(huán)境科學(xué)研究院生態(tài)研究所,北京 100012
2.中國(guó)地質(zhì)調(diào)查局自然資源綜合調(diào)查指揮中心,北京 100055
生物入侵是導(dǎo)致生物多樣性喪失的重要因素之一,對(duì)生態(tài)系統(tǒng)和經(jīng)濟(jì)發(fā)展產(chǎn)生不可預(yù)估的影響.外來(lái)入侵物種一旦定殖成功將很難根除,提前預(yù)測(cè)其潛在分布是制定物種防控策略以阻其種群蔓延的重要基礎(chǔ)[1].近年來(lái),生態(tài)位模型被越來(lái)越多地運(yùn)用在外來(lái)入侵物種潛在分布分析中,其基本原理是利用外來(lái)入侵物種在原產(chǎn)地分布數(shù)據(jù)和環(huán)境條件構(gòu)建模型,推算外來(lái)入侵物種生長(zhǎng)所需的生態(tài)環(huán)境要求,進(jìn)而投影至不同空間或時(shí)間中預(yù)測(cè)物種的實(shí)際分布和潛在分布[2-3].與其他生態(tài)位模型相比,MaxEnt模型能夠較好地處理預(yù)測(cè)變量之間復(fù)雜的交互作用,在預(yù)測(cè)精準(zhǔn)度和穩(wěn)定性等方面更具有優(yōu)越性[4].
黃頂菊(Flaveria bidentis)隸屬于菊科(Asteraceae)堆心菊族(Helenieae),屬一年生草本植物,原產(chǎn)巴西、玻利維亞、厄瓜多爾、秘魯、阿根廷、智利、巴拉圭等南美洲國(guó)家和地區(qū),然后擴(kuò)散至中美洲、北美洲南部和西印度群島,后來(lái)因引種等原因傳播至非洲的埃及和南非,歐洲的英國(guó)和法國(guó)、澳大利亞以及亞洲的日本等地[5].1994年黃頂菊首次出現(xiàn)在河北省邯鄲市永年縣,隨后以河北省中南部為中心向周邊其他省市擴(kuò)散,目前已擴(kuò)散至河北省邯鄲市、邢臺(tái)市、衡水市、石家莊市、保定市、廊坊市、滄州市和唐山市8個(gè)地級(jí)市107個(gè)區(qū)縣,天津市靜海區(qū)、西青區(qū)、津南區(qū)、寧河區(qū)、濱海新區(qū)、寶坻區(qū)、薊州區(qū)、河西區(qū)、河北區(qū)和南開區(qū)10市區(qū),河南省安陽(yáng)市、鶴壁市、濮陽(yáng)市、新鄉(xiāng)市、鄭州市和開封市6個(gè)地級(jí)市18個(gè)區(qū)縣,山東省聊城市、德州市、濟(jì)南市、濱州市和東營(yíng)市5個(gè)地級(jí)市24個(gè)區(qū)縣,以及山西省長(zhǎng)治市黎城縣局部[6].黃頂菊喜光、喜濕、嗜鹽,植株生長(zhǎng)迅速、花期長(zhǎng)且種子結(jié)實(shí)量大,具有較強(qiáng)的擴(kuò)散能力,其植株產(chǎn)生的化感物質(zhì)能對(duì)20多種植物種子發(fā)芽產(chǎn)生抑制作用[7],一旦侵入農(nóng)田、草地等區(qū)域,會(huì)對(duì)已有的穩(wěn)定的生態(tài)環(huán)境造成嚴(yán)重的破壞[8].2010年原環(huán)境保護(hù)部將黃頂菊列入《中國(guó)第二批外來(lái)入侵物種名單》.
目前,國(guó)內(nèi)外學(xué)者陸續(xù)開展了黃頂菊調(diào)查與監(jiān)測(cè)、入侵機(jī)理、生態(tài)影響、防控策略等研究[5-8],揭示了黃頂菊入侵我國(guó)后的生物生態(tài)學(xué)適應(yīng)性,闡述了影響黃頂菊種子萌發(fā)、幼苗生長(zhǎng)的環(huán)境限制因素,對(duì)于潛在適生區(qū)變化趨勢(shì)及早期預(yù)警管理方面研究不多.白藝珍等[9-10]運(yùn)用GARP、MaxEnt、ENFA、Bioclim、Domain、CLIMEX等生態(tài)位模型預(yù)測(cè)了黃頂菊在中國(guó)的潛在適生區(qū),但未考慮未來(lái)氣候條件下黃頂菊適生分布的變化.李安等[11]基于MaxEnt模型預(yù)測(cè)了當(dāng)前及未來(lái)氣候條件下黃頂菊在中國(guó)的潛在適生分布區(qū),其在模型構(gòu)建中存在物種采樣偏差、空間維度過(guò)高、未降低環(huán)境變量空間相關(guān)性等問(wèn)題,且未指明潛在適生區(qū)在管理中的應(yīng)用.黃頂菊具有較強(qiáng)的繁殖能力和擴(kuò)散能力,一旦種群暴發(fā)會(huì)造成極大危害,結(jié)合種群動(dòng)態(tài)研究識(shí)別其潛在擴(kuò)散風(fēng)險(xiǎn)已成為入侵物種早期監(jiān)測(cè)預(yù)警的關(guān)鍵[12].基于此,研究利用黃頂菊在南美原產(chǎn)地分布數(shù)據(jù)和環(huán)境變量構(gòu)建本土生態(tài)位模型,投影至我國(guó)并預(yù)測(cè)當(dāng)前和未來(lái)氣候情景下黃頂菊的潛在分布格局,結(jié)合黃頂菊實(shí)際分布狀況及其在不同氣候情景下的潛在分布變化趨勢(shì),在黃頂菊擴(kuò)散前緣區(qū)建立監(jiān)測(cè)阻截帶,分級(jí)管控黃頂菊發(fā)生區(qū)及潛在適生區(qū),以期為其他外來(lái)入侵物種的風(fēng)險(xiǎn)評(píng)估與預(yù)警提供參考依據(jù).
1 材料與方法
1.1 物種分布數(shù)據(jù)
黃頂菊在全球的分布數(shù)據(jù)源于全球生物多樣性信息網(wǎng)絡(luò)(Global Biodiversity Information Facility, GBIF)、標(biāo)本數(shù)據(jù)、實(shí)地調(diào)查及相關(guān)文獻(xiàn)[6],其中原產(chǎn)地分布點(diǎn)集中于南美洲中部和西部,中國(guó)分布點(diǎn)集中于華北地區(qū)南部、華中地區(qū)北部、華東地區(qū)北部,這些分布數(shù)據(jù)基本包括了黃頂菊在原產(chǎn)地和我國(guó)的分布狀況.采樣偏差會(huì)造成生態(tài)位模型對(duì)物種需求的過(guò)度擬合,從而降低生態(tài)位模型的轉(zhuǎn)移能力[13-14],為降低采樣點(diǎn)的空間自相關(guān)以及與環(huán)境數(shù)據(jù)范圍一致,將研究區(qū)分成1 km×1 km的網(wǎng)格,隨機(jī)獲取網(wǎng)格中唯一物種分布點(diǎn),總計(jì)獲得178個(gè)原產(chǎn)地分布點(diǎn)和524個(gè)中國(guó)分布記錄.
1.2 環(huán)境變量
該研究所用的環(huán)境變量為22個(gè),分為氣候和地形兩類.氣候數(shù)據(jù)來(lái)自WorldClim 2.1(http://www.world clim.org)中的19個(gè)生物氣候因子(年均溫、晝夜溫差月均值、等溫性、溫度季節(jié)性變動(dòng)率、最熱月最高溫、最冷月最低溫、年溫度變化范圍、最濕季均溫、最干季均溫、最暖季均溫、最冷季均溫、年降水量、最濕月降水量、最干月降水量、降水量季節(jié)性變化、最濕季降水量、最干季降水量、最暖季降水量和最冷季降水量)[15],分辨率為30″(約1 km),數(shù)據(jù)年份為1970?2000年.地形數(shù)據(jù)包括海拔、坡度和坡向,海拔源于GTOPO(http://edcftp.cr.usgs.gov/pub/data/gtopo 30/global),坡度及坡向由海拔數(shù)據(jù)基于ArcGIS 10.6軟件處理得到.未來(lái)氣候模式數(shù)據(jù)采用2070年(2061?2080年)WorldClim 2.1中我國(guó)國(guó)家氣候中心開發(fā)的BCC_CSM2_MR模型對(duì)應(yīng)的數(shù)據(jù),該模型在全球溫度、降水分布等方面模擬能力較高[16].未來(lái)氣候模式包括4種共享的社會(huì)經(jīng)濟(jì)途徑(SSPs,shared socioeconomic pathways),其中SSPs 1-2.6代表低輻射強(qiáng)迫下可持續(xù)發(fā)展路徑,SSPs 2-4.5代表中輻射強(qiáng)迫下中間發(fā)展路徑,SSPs 3-7.0代表偏高輻射強(qiáng)迫下區(qū)域競(jìng)爭(zhēng)發(fā)展路徑,SSPs 5-8.5為高輻射強(qiáng)迫下的常規(guī)發(fā)展路徑[17-18].輻射強(qiáng)迫越高,增溫越快.
1.3 模型構(gòu)建與評(píng)價(jià)
應(yīng)用MaxEnt模型對(duì)黃頂菊進(jìn)行適生區(qū)預(yù)測(cè)分析,隨機(jī)選取75%的物種分布點(diǎn)和10 000個(gè)擬不存在分布點(diǎn)作為訓(xùn)練數(shù)據(jù),剩余25%的數(shù)據(jù)用于模型的檢驗(yàn)和校正,模型訓(xùn)練方法選用線性(linear)、二次型(quadratic)和片段化(hinge)程序,其他參數(shù)為軟件默認(rèn)值,分析結(jié)果以Logistic格式、ASCII類型文件輸出.采用ROC曲線下面積(area under curve,AUC)值評(píng)價(jià)模型預(yù)測(cè)結(jié)果的可靠性,AUC值在0.5~0.6之間為失敗,在0.6~0.7之間為較差,在0.7~0.8之間為一般,在0.8~0.9之間為良好,在0.9~1.0之間為優(yōu)秀.AUC值越接近1,說(shuō)明與隨機(jī)分布相距越遠(yuǎn),環(huán)境變量與預(yù)測(cè)物種地理分布之間的相關(guān)性較大,模型預(yù)測(cè)結(jié)果比較準(zhǔn)確[19].
1.4 早期監(jiān)測(cè)預(yù)警區(qū)域劃定
基于上述構(gòu)建模型投影至中國(guó)以預(yù)測(cè)當(dāng)前及未來(lái)情景下黃頂菊潛在分布區(qū).模型預(yù)測(cè)結(jié)果為適生值位于0~1之間的連續(xù)的柵格數(shù)據(jù),適生值越接近1,表明物種存在的適生概率越高.研究選用靈敏度和特異性數(shù)值之和最大的閾值判定方法將預(yù)測(cè)結(jié)果劃分為4個(gè)等級(jí):適生值<0.35,為非適生區(qū);0.35≤適生值<0.7,為低適生區(qū);0.7≤適生值<0.8,為中適生區(qū);0.8≤適生值<1.0,為高適生區(qū).
聚焦黃頂菊中適生區(qū)和高適生區(qū),將黃頂菊適生值≥0.7的區(qū)域設(shè)為中-高潛在適生區(qū),賦值為1.將黃頂菊適生值<0.7的區(qū)域賦值為0.基于GIS柵格計(jì)算模塊,以當(dāng)前氣候情景下黃頂菊中-高潛在適生區(qū)面積為基礎(chǔ),計(jì)算未來(lái)氣候情景下黃頂菊中-高潛在適生區(qū)面積變化,并劃分為新增區(qū)域(當(dāng)前適生值為0,未來(lái)適生值為1)、保留區(qū)域(當(dāng)前適生值為1,未來(lái)適生值為1)、收縮區(qū)域(當(dāng)前適生值為1,未來(lái)適生值為0)和不適生區(qū)域(當(dāng)前適生值為0,未來(lái)適生值為0).基于GIS分區(qū)統(tǒng)計(jì)工具Zonal分別計(jì)算不同氣候情景下黃頂菊中-高潛在適生區(qū)幾何質(zhì)心(centroid),通過(guò)質(zhì)心位置的變化表征氣候變暖背景下黃頂菊潛在適生區(qū)地理空間重心的變遷.基于GIS技術(shù)提取不同氣候情景下黃頂菊中-高潛在適生區(qū)內(nèi)主要環(huán)境變量范圍和平均值.結(jié)合當(dāng)前氣候情景下黃頂菊中-高潛在適生區(qū)地理格局及其未來(lái)擴(kuò)散趨勢(shì),劃定我國(guó)黃頂菊重點(diǎn)監(jiān)測(cè)預(yù)警區(qū)域.
2 結(jié)果與分析
2.1 當(dāng)前氣候情景下黃頂菊潛在適生區(qū)的預(yù)測(cè)
為降低模型復(fù)雜度,增強(qiáng)模型轉(zhuǎn)移能力[20],基于刀切法對(duì)22個(gè)環(huán)境變量進(jìn)行重要性排序,利用多重共線性分析去除相關(guān)度絕對(duì)值高于0.8的環(huán)境變量[21],結(jié)合黃頂菊生活習(xí)性篩選出溫度季節(jié)性變動(dòng)率、年均溫、年降水量、最冷月最低溫、海拔、最干月降水量等6個(gè)影響黃頂菊地理分布的主要因素.基于原產(chǎn)地分布數(shù)據(jù)及篩選的環(huán)境變量構(gòu)建生態(tài)位模型并投影至全球范圍,發(fā)現(xiàn)模型的AUC值為0.882,遠(yuǎn)高于隨機(jī)預(yù)測(cè)值(0.5),模型模擬效果良好,說(shuō)明該模型能夠很好地?cái)M合已知分布點(diǎn)的環(huán)境需求.
基于上述構(gòu)建的模型投影至中國(guó)分析當(dāng)前氣候情景下黃頂菊適生分布格局,結(jié)果表明,黃頂菊潛在分布區(qū)面積為1.72×106km2,約占陸地面積的17.94%,適生范圍涵蓋華北平原、東北平原、黃土高原和云貴高原.中-高潛在適生區(qū)面積為4.87×105km2,適生范圍以華北平原為主,具體包括河北省中部和南部、北京市大部、天津市、遼寧省中部及沿海地區(qū)、山東省大部、河南省北部、山西省南部、陜西省中部及東部.高適生區(qū)面積為1.80×105km2,集中分布于河北省邢臺(tái)市、邯鄲市中部和西部、衡水市東部、石家莊市中部和西部、保定市大部、滄州市大部、廊坊市、唐山市、秦皇島市中部和南部,北京市中部和南部,天津市大部,遼寧省葫蘆島市南部沿海、丹東市零星地區(qū),山東省德州市、濱州市大部、東營(yíng)市南部、濟(jì)南市、泰安市大部、淄博市大部、濰坊市中部和西部、臨沂市北部、日照市西部,以及河南省安陽(yáng)市西部,山西省南部零星地區(qū),陜西省東部沿黃地帶等區(qū)域.中適生區(qū)面積為3.07×105km2,主要沿高適生區(qū)向周邊延伸分布,遼寧省中部和南部、河南省北部、山西省南部、陜西省中東部有大面積黃頂菊中適生區(qū).比較黃頂菊實(shí)際記錄點(diǎn)和適生分布格局發(fā)現(xiàn),99%的黃頂菊實(shí)際記錄點(diǎn)發(fā)生在中-高潛在適生區(qū)內(nèi),山東省中部、北京市中南部、遼寧省南部、山西省南部、陜西省中部具有較大面積的中-高潛在適生區(qū).
2.2 不同氣候情景下黃頂菊潛在分布格局的變化
當(dāng)采取低輻射強(qiáng)迫可持續(xù)發(fā)展路徑(SSPs 1-2.6)時(shí),2070年黃頂菊潛在適生區(qū)的面積為2.60×106km2,約占我國(guó)陸地面積的27.07%,其中中-高潛在適生區(qū)的面積為7.18×105km2.與當(dāng)前氣候情景相比,SSPs 1-2.6情景下黃頂菊中-高潛在適生區(qū)生境新增2.45×105km2(見表1),新增區(qū)域主要位于當(dāng)前適生區(qū)北緣,具體包括陜西省東北部、山西省中部零星地區(qū)、河北省北部、遼寧省北部、內(nèi)蒙古與遼寧交界處和新疆吐魯番零星地區(qū);適生生境收縮1.49×104km2,收縮區(qū)域大都位于當(dāng)前適生區(qū)南側(cè),具體包括山東省南部、河南省中部等.
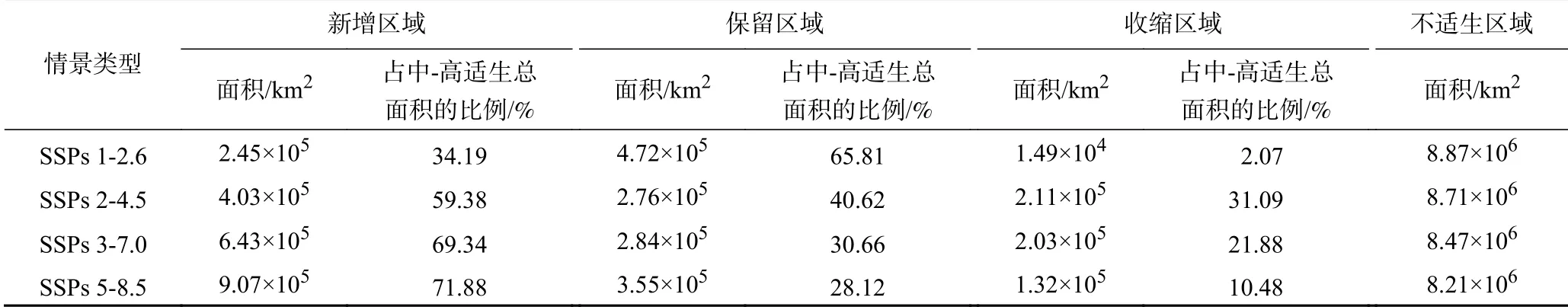
表1 與當(dāng)前氣候情景相比不同時(shí)期黃頂菊適生區(qū)的變化Table 1 The Change of Flaveria bidentis habitat in different periods compared with the current climate scenario
當(dāng)采取中等輻射強(qiáng)迫的中間發(fā)展路徑(SSPs 2-4.5)時(shí),2070年黃頂菊潛在適生區(qū)的面積為2.85×106km2,約占我國(guó)陸地面積的29.67%,其中中-高潛在適生區(qū)的面積為6.79×105km2. 與當(dāng)前氣候情景相比,SSPs 2-4.5情景下黃頂菊中-高潛在適生區(qū)生境新增4.03×105km2,新增區(qū)域仍以當(dāng)前適生區(qū)北緣為主,集中分布于陜西省東北部、山西省中部零星地區(qū)、河北省北部、遼寧省北部、內(nèi)蒙古西部地區(qū)、吉林省西部、黑龍江南部零星地區(qū)等;適生生境收縮2.11×105km2,收縮區(qū)域主要位于山東省西部和南部、河南省大部、山西省南部、陜西省中部、河北省南部零星地區(qū)等當(dāng)前適生區(qū)南部地區(qū).
當(dāng)采取區(qū)域競(jìng)爭(zhēng)發(fā)展路徑(SSPs 3-7.0)時(shí),2070年黃頂菊潛在適生區(qū)的面積為3.29×106km2,約占我國(guó)陸地面積的34.32%,其中中-高潛在適生區(qū)的面積為9.27×105km2.與當(dāng)前氣候情景相比,SSPs 3-7.0情景下黃頂菊中-高潛在適生區(qū)生境新增6.43×105km2,新增區(qū)域繼續(xù)向高緯度擴(kuò)散,具體包括陜西省北部、山西省中部、河北省北部、遼寧省北部、內(nèi)蒙古西部地區(qū)、吉林省西部、黑龍江南部、新疆阿克蘇和吐魯番零星地區(qū);生境收縮2.03×105km2,收縮區(qū)域包括山東省西部和南部、河南省大部、山西省南部、陜西省中部、河北省南部等.
當(dāng)采取高輻射強(qiáng)迫下的常規(guī)發(fā)展路徑(SSPs 5-8.5)時(shí),2070年黃頂菊潛在適生區(qū)的面積為3.45×106km2,約占我國(guó)陸地面積的35.98%,其中中-高潛在適生區(qū)的面積為1.26×106km2. 與當(dāng)前氣候情景相比,SSPs 5-8.5情景下黃頂菊中-高潛在適生生境新增9.07×105km2,新增區(qū)域仍以高緯度地區(qū)為主,具體包括陜西省北部、山西省中部及北部、河北省北部、遼寧省北部、內(nèi)蒙古西部地區(qū)、吉林省大部、黑龍江省南部和東部、新疆阿克蘇和吐魯番部分地區(qū);適生生境收縮1.32×105km2,收縮區(qū)域包括山東省南部、河南省大部、陜西省中部等中緯度地區(qū).
2.3 不同氣候情景下黃頂菊分布中心的變化
由圖1可見,黃頂菊在當(dāng)前情景下分布中心位于河北省衡水市東部(37.45°N、116.23°E). 2070年SSPs 1-2.6情景下黃頂菊分布中心向東北遷移至河北省滄州市中部(38.32°N、116.50°E),遷移距離約為101.94 km.SSPs 2-4.5情景下黃頂菊分布中心轉(zhuǎn)移至遼寧省葫蘆島市南部(40.29°N、119.90°E),與當(dāng)前情景相比分布中心大幅向東北方向偏移,遷移距離為295.73 km.SSPs 3-7.0情景下黃頂菊分布中心轉(zhuǎn)移至遼寧省朝陽(yáng)市中部(41.52°N、119.93°E),與當(dāng)前情景相比繼續(xù)向東北偏移567.69 km. SSPs 5-8.5情景下黃頂菊分布中心轉(zhuǎn)移至內(nèi)蒙古自治區(qū)赤峰市南部(41.60°N、119.08°E).整體來(lái)看,黃頂菊分布中心由華北中緯度地區(qū)向東北高緯度地區(qū)遷移趨勢(shì)明顯.
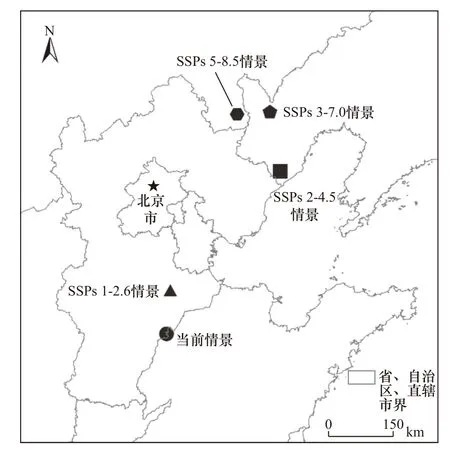
圖1 不同氣候情景下黃頂菊適生分布中心的遷移Fig.1 Migration of the center of suitable habitat for Flaveria bidentis under different climatescenario
2.4 不同氣候情景下環(huán)境變量的綜合分析
基于MaxEnt模型中刀切法獲得各環(huán)境變量對(duì)黃頂菊適生概率的相對(duì)重要性,發(fā)現(xiàn)溫度季節(jié)性變動(dòng)率、海拔、最冷月最低溫對(duì)黃頂菊適生分布預(yù)測(cè)有重要影響,貢獻(xiàn)率分別為27.05%、22.58%、17.73%,其次為年降水量、最干月降水量和年均溫,貢獻(xiàn)率分別為12.19%、11.43%、6.30%、9.01%.
基于GIS技術(shù)提取了不同氣候情景下黃頂菊中-高潛在適生區(qū)內(nèi)溫度季節(jié)性變動(dòng)率、海拔、最冷月最低溫范圍和平均值,發(fā)現(xiàn)當(dāng)前情景下黃頂菊中-高潛在適生區(qū)內(nèi)溫度季節(jié)性變動(dòng)率為1 068.71±86.44,海拔為(249.59±308.13) m,最冷月最低溫為(?9.32±3.48)℃,未來(lái)氣候變暖導(dǎo)致溫度季節(jié)性變動(dòng)率由1 108.34±112.68升至1 257.54±146.77,海拔由(378.71±398.43) m升至(448.94±402.83) m,最冷月最低溫由(?9.32±3.48)℃降至(?11.64±5.52)℃(見表2).全球變暖促使中國(guó)境內(nèi)黃頂菊向溫差變化大、高海拔、高緯度區(qū)域擴(kuò)散.
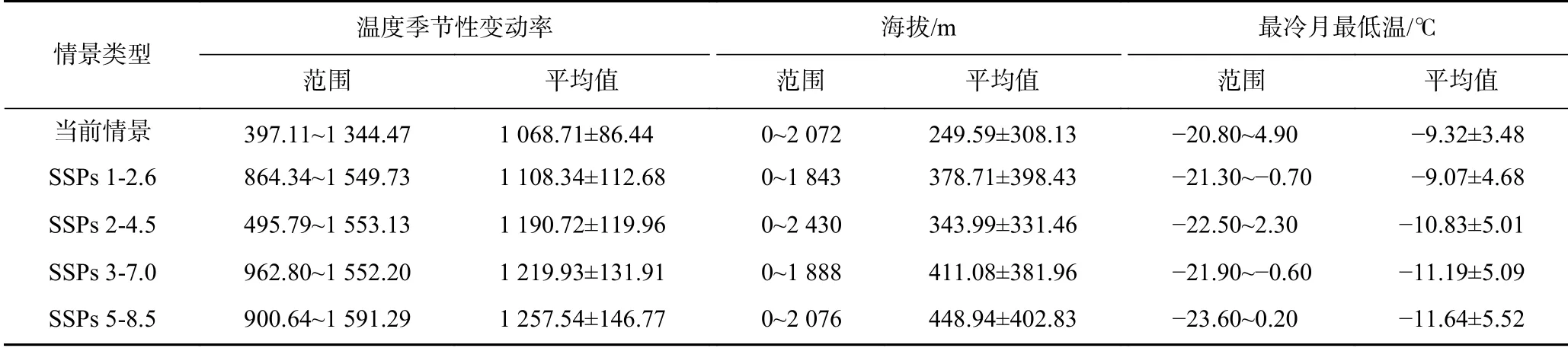
表2 不同氣候情景下黃頂菊適生分布區(qū)內(nèi)重要環(huán)境變量的變化Table 2 Changes in important environmental variables under different climate scenarios of Flaveria bidentis
2.5 中國(guó)黃頂菊早期預(yù)警
綜合比對(duì)黃頂菊在我國(guó)實(shí)際分布區(qū)、當(dāng)前情景下潛在適生區(qū)與未來(lái)擴(kuò)張區(qū),發(fā)現(xiàn)其在我國(guó)具有較大潛在分布空間,為阻斷其向鄰近地區(qū)進(jìn)一步擴(kuò)散蔓延,建議在天津市北部、山西省東部、河南省北部、山東省北部等黃頂菊擴(kuò)散前緣區(qū)建立擴(kuò)散前沿監(jiān)測(cè)阻截帶,通過(guò)生態(tài)修復(fù)與物種調(diào)控等措施增強(qiáng)本地植物群落對(duì)黃頂菊的抵御能力.
為加強(qiáng)黃頂菊屬地管理,建議以地級(jí)及以上城市行政區(qū)為單位劃分重點(diǎn)管控區(qū)、重點(diǎn)監(jiān)測(cè)區(qū)和重點(diǎn)預(yù)防區(qū)(見表3),并制定預(yù)警策略.其中天津市、山西省長(zhǎng)治市、山東省西部和北部、河南省北部、河北省中部和南部等黃頂菊發(fā)生區(qū)列為重點(diǎn)管控區(qū),著重開展黃頂菊防控和生態(tài)修復(fù)工作.北京市、陜西省北部、山西省南部、山東省中部和南部、遼寧省中部和南部、河南省西北部、河北省北部等黃頂菊潛在適生區(qū)列為重點(diǎn)監(jiān)測(cè)區(qū),加強(qiáng)黃頂菊監(jiān)測(cè)和早期預(yù)警.山西省北部、內(nèi)蒙古西部地區(qū)、遼寧省東北部和西北部、吉林省、黑龍江省南部和東部等黃頂菊未來(lái)擴(kuò)張區(qū)列為重點(diǎn)預(yù)防區(qū),防范黃頂菊入侵(見表3).

表3 黃頂菊預(yù)警區(qū)域的地級(jí)及以上城市行政區(qū)名單Table 3 The list of cities at prefecturelevel or above with in the early warning area of Flaveria bidentis
3 討論
生態(tài)位模型是利用外來(lái)入侵物種已知分布數(shù)據(jù)和環(huán)境數(shù)據(jù)構(gòu)建模型,判斷外來(lái)入侵物種生長(zhǎng)所需的環(huán)境需求,然后投影至選定時(shí)空范圍內(nèi),以概率形式反映物種對(duì)生境的偏好程度[2,22].一般來(lái)言,外來(lái)入侵物種潛在分布區(qū)預(yù)測(cè)有本土模型構(gòu)建方案和全球模型構(gòu)建方案兩種方法,在生態(tài)位保守的假設(shè)前提下,前者模擬結(jié)果比后者更具有風(fēng)險(xiǎn)分析的價(jià)值[23-24],但在轉(zhuǎn)移至入侵地時(shí)常出現(xiàn)模型轉(zhuǎn)移能力低等問(wèn)題[25].物種在研究區(qū)內(nèi)平衡狀態(tài)、采樣點(diǎn)偏差、環(huán)境變量是影響生態(tài)位模型轉(zhuǎn)移能力的重要影響要素[3],其中環(huán)境變量篩選時(shí)既要考慮其對(duì)物種分布的限制性作用,也要權(quán)衡模型構(gòu)建區(qū)域、環(huán)境變量空間維度和變量間相關(guān)性對(duì)物種分布的影響[3,24].研究基于環(huán)境變量重要性排序,選擇了溫度季節(jié)性變動(dòng)率、年均溫、年降水量、最冷月最低溫、海拔、最干月降水量等6個(gè)相關(guān)性較小且與黃頂菊生物學(xué)特征密切相關(guān)的環(huán)境變量進(jìn)行潛在適生區(qū)預(yù)測(cè),其中最冷月最低溫、最干月降水量有助于模擬黃頂菊在低溫脅迫和干旱脅迫等極端氣候下的潛在分布[26],溫度季節(jié)性變動(dòng)率、年均溫、年降水量會(huì)影響黃頂菊種子萌發(fā)和植株生長(zhǎng)[26-28],地形有助于細(xì)化海拔對(duì)潛在適生格局影響.基于篩選后環(huán)境變量構(gòu)建的本土模型能夠較好地預(yù)測(cè)黃頂菊在中國(guó)的實(shí)際分布.
外來(lái)物種入侵是一個(gè)長(zhǎng)期的歷史過(guò)程,從最初引入至大面積擴(kuò)散通常需要持續(xù)幾十年甚至幾個(gè)世紀(jì)[29].探討特定氣候情景與入侵植物生態(tài)格局之間的相互作用是評(píng)估物種未來(lái)地理分布格局、管控入侵植物的重要研究方法[30].研究基于黃頂菊原產(chǎn)地分布數(shù)據(jù)和環(huán)境變量構(gòu)建本土MaxEnt生態(tài)位模型,投影至我國(guó)并預(yù)測(cè)了當(dāng)前氣候條件下黃頂菊潛在適生格局,發(fā)現(xiàn)黃頂菊潛在適生區(qū)包括華北平原、東北平原、黃土高原和云貴高原等,中-高潛在適生區(qū)集中分布在華北平原,研究結(jié)果能較好地?cái)M合黃頂菊在我國(guó)的實(shí)際分布狀況,AUC值也顯示良好,結(jié)果可靠.與黃頂菊實(shí)際分布相比,山東省中部、北京市中部和南部、遼寧省南部、山西省南部、陜西省中部具有較大面積的中-高潛在適生區(qū),黃頂菊在我國(guó)沒(méi)有達(dá)到其最大潛在分布范圍,仍有可能繼續(xù)擴(kuò)散.這與鄭志鑫等[6]基于文獻(xiàn)、標(biāo)本記錄及實(shí)地調(diào)查得出的黃頂菊在我國(guó)仍處于快速擴(kuò)散階段的結(jié)論基本一致.此外,研究將本土生態(tài)位模型投影至我國(guó)并預(yù)測(cè)了2070年未來(lái)氣候條件下黃頂菊的潛在適生格局,發(fā)現(xiàn)氣候變暖導(dǎo)致黃頂菊潛在適生區(qū)的面積增加,并呈向北擴(kuò)張、向南收縮的趨勢(shì),分布中心由華北中緯度地區(qū)向東北高緯度地區(qū)遷移,氣溫增加越明顯,黃頂菊潛在適生區(qū)向北擴(kuò)張的面積越大.這與鐘永德等[31-32]得出的氣候變暖會(huì)促進(jìn)生物入侵的結(jié)論相符.在全球變暖的大背景下,氣候變化的幅度可能會(huì)改變區(qū)域降水量及分配,引發(fā)極端氣候和自然災(zāi)害,弱化本地植物群落對(duì)外來(lái)物種的抵御能力,進(jìn)而間接促進(jìn)外來(lái)入侵物種的快速擴(kuò)散[31].氣候變化雖可改變?nèi)肭治锓N空間分布格局和適宜生境,但其變化趨勢(shì)在連續(xù)時(shí)間上或連續(xù)氣候幅度上不具有恒定性[33].如未來(lái)氣候情景下克氏原螯蝦適生區(qū)在空間分布上不僅有緯度方向上的擴(kuò)散,也有向海拔較高地區(qū)遷移的趨勢(shì)[34];豚草在未來(lái)氣候變化背景下向更冷和溫度季節(jié)性更強(qiáng)的地區(qū)入侵[35].惡性外來(lái)入侵植物歐洲菊苣和天竺葵在相同未來(lái)氣候情景下全球生境適宜性變化規(guī)律迥異,前者在南美和歐洲西海岸等高緯度地區(qū)的生境適宜程度顯著增加,后者生境適宜程度顯著增加區(qū)域?yàn)闊釒У貐^(qū)[30].氣候變化對(duì)外來(lái)入侵物種潛在適生分布影響可能是氣候情景與外來(lái)入侵植物內(nèi)部特征交互作用的結(jié)果[36],在預(yù)測(cè)外來(lái)入侵植物動(dòng)態(tài)分布時(shí)應(yīng)針對(duì)具體情況和物種特異性慎重對(duì)待.
通常情況下,入侵植物在種群規(guī)模較小時(shí)很難被監(jiān)測(cè)到,一旦發(fā)現(xiàn)往往已處于種群爆發(fā)階段,難以根除,對(duì)其開展風(fēng)險(xiǎn)評(píng)估、早期預(yù)警和快速阻截是有效防控生物入侵的重要手段[37],區(qū)分不同區(qū)域可入侵性水平并確定物種優(yōu)先管控區(qū)是管理外來(lái)入侵物種的重要環(huán)節(jié)[38-40].考慮到國(guó)家“遏增量、去存量”的生物入侵防控需求,以及地方生物安全屬地管理要求,研究基于黃頂菊在我國(guó)實(shí)際分布狀況及其在不同氣候情景下潛在分布的變化趨勢(shì),以我國(guó)地級(jí)及以上城市行政區(qū)為管控單元明確了黃頂菊實(shí)際分布、潛在適生分布和未來(lái)擴(kuò)張范圍,并制定了針對(duì)性的防控措施,具體包括:在黃頂菊擴(kuò)散前緣劃定監(jiān)測(cè)阻截區(qū),構(gòu)建和完善人工群落阻截帶,增強(qiáng)本地植物群落對(duì)黃頂菊的抵御能力;將黃頂菊發(fā)生區(qū)列為重點(diǎn)管控區(qū),著重開展黃頂菊的普查和滅除工作,重視替代控制與生態(tài)修復(fù),逐步壓縮黃頂菊分布范圍;把黃頂菊潛在適生區(qū)列為重點(diǎn)監(jiān)測(cè)區(qū),開展動(dòng)態(tài)監(jiān)測(cè)、風(fēng)險(xiǎn)評(píng)估和早期預(yù)警,增強(qiáng)河流、港口、交通干線等區(qū)域黃頂菊繁殖體檢疫,一經(jīng)發(fā)現(xiàn)應(yīng)及時(shí)清除.把黃頂菊未來(lái)擴(kuò)張區(qū)列為重點(diǎn)預(yù)防區(qū),堅(jiān)持應(yīng)急和防御并存,避免氣候變化導(dǎo)致黃頂菊進(jìn)一步入侵.
外來(lái)物種入侵與擴(kuò)散與眾多因素相關(guān),如氣候因素、地形因素等非生物因素,以及人類活動(dòng)、物種相互作用等生物因素.基于生態(tài)模型預(yù)測(cè)物種分布常被模型所用的環(huán)境變量限制,該研究強(qiáng)調(diào)了氣候、地形等非生物因素對(duì)黃頂菊分布的限制作用,忽略了人類活動(dòng)、物種間相互作用和物種遷移能力等生物因素對(duì)外來(lái)入侵物種擴(kuò)散的影響,這可能對(duì)氣候變化情景下黃頂菊潛在分布區(qū)及預(yù)警管理帶來(lái)一定的偏差,需要在以后的研究中進(jìn)一步完善.
4 結(jié)論
a)研究充分考慮了物種采樣點(diǎn)偏差、環(huán)境變量空間維度、環(huán)境變量間空間自相關(guān)性等對(duì)生態(tài)模型模擬結(jié)果的影響,基于黃頂菊在南美原產(chǎn)地分布數(shù)據(jù)和環(huán)境變量構(gòu)建本土MaxEnt模型,投影至我國(guó)并預(yù)測(cè)了當(dāng)前和未來(lái)氣候條件下黃頂菊的潛在分布格局.MaxEnt模型的AUC值為0.882,模擬效果良好.
b)與實(shí)際分布相比,當(dāng)前氣候條件下黃頂菊在山東省中部、北京市中部和南部、遼寧省南部、山西省南部、陜西省中部具有較大面積的中-高潛在適生區(qū),黃頂菊在我國(guó)沒(méi)有達(dá)到其最大潛在分布范圍,仍有可能繼續(xù)擴(kuò)散.
c)未來(lái)氣候條件下黃頂菊潛在適生分布預(yù)測(cè)結(jié)果表明,氣候變暖導(dǎo)致黃頂菊潛在適生區(qū)明顯向北擴(kuò)張、向南縮小,分布中心由華北中緯度地區(qū)向東北高緯度地區(qū)遷移,氣溫增加越明顯,黃頂菊潛在適生區(qū)向北擴(kuò)張面積越大.
d)綜合比對(duì)黃頂菊實(shí)際分布狀況及其在不同氣候情景下潛在分布的變化趨勢(shì),結(jié)合國(guó)家和地方管控需求,提出在黃頂菊分布區(qū)邊緣建立擴(kuò)散前沿監(jiān)測(cè)阻截帶,以地級(jí)及以上城市行政區(qū)為管控單元?jiǎng)澏ㄖ攸c(diǎn)管控區(qū)、重點(diǎn)監(jiān)測(cè)區(qū)和重點(diǎn)預(yù)防區(qū)等針對(duì)性防控措施.研究形成的黃頂菊分區(qū)監(jiān)管策略可為其他外來(lái)入侵物種風(fēng)險(xiǎn)評(píng)估與預(yù)警提供參考依據(jù).
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
童話王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
核科學(xué)與工程(2015年4期)2015-09-26 11:59:03
