半滑舌鰨腸道中抗氧化活性乳酸桿菌的篩選與鑒定
2022-12-26 11:12:24包瑞璇呂明生王淑軍
中國飼料 2022年23期
郭 東, 包瑞璇, 王 逸, 盧 靜, 呂明生*, 王淑軍
(1.江蘇海洋大學海洋生物資源與環境重點實驗室/海洋生物技術重點實驗室,江蘇海洋大學,江蘇連云港 225005;2.江蘇省海洋生物產業技術協同創新中心,江蘇連云港 222000)
半滑舌鰨魚(Cynoglossus semilaevis)屬于舌鰨屬,經中國水產研究院黃海研究所成功訓化后,是我國重要水產養殖品種之一(張馨馨,2021)。魚體內脂肪中含有豐富的不飽和脂肪酸,具有防止動脈粥樣硬化、防治心血管疾病、增強記憶力和防止視力衰退等功效(張永真等,2016;王娜,2010)。該種魚多產于我國渤海灣、膠州灣和海州灣等海域(孫中之等,2005),近年來隨著養殖數量的增加,腹水病、爛鰭病、腸炎病等疾病經常發生(蘇兆軍,2015)。水產養殖中急需有效的病害防控措施。
乳酸菌是水產養殖中重要的微生態菌種之一,通過產生有機酸、乳酸菌素等提高養殖動物抵抗力(鞏華等,2018)。研究發現,乳酸菌和發酵產物還具有抗氧化作用。通過在飼料中添加具有抗菌和抗氧化作用的乳酸菌,可以實現健康養殖(曹衛華,2017)。本研究從半滑舌鰨魚腸道中篩選乳酸菌,并對其生長特性、抑菌作用、抗氧化活性以及其對飼料脂質抗氧化作用進行了研究,擬為獲得的乳酸菌的應用奠定基礎。
1 材料和方法
1.1 試驗材料
1.1.1 原材料 健康的半滑舌鰨魚:水產品養殖場;高脂半滑舌鰨魚飼料:通威股份;創傷弧菌、大腸桿菌、副溶血弧菌、維氏氣單胞菌、霍亂弧菌、金黃色葡萄球菌、枯草芽孢桿菌、庫氏鏈霉菌、擬桿菌、嗜水氣單胞菌等指示菌株:江蘇海洋大學海洋資源與環境重點實驗室。
1.1.2 試劑與培養基 胰蛋白酶、胃蛋白酶、木瓜蛋白酶、蛋白酶K:購于南京都萊生物技術有限公司;過氧化氫酶:購于上海阿拉丁試劑;革蘭氏染色試劑盒:購于北京索萊寶科技有限公司;DNA marker2000、DNA marker5000:購自Takara-寶日醫生物技術(北京)有限公司;電泳用瓊脂糖:購于BIOWEST公司;細菌DNA基因組提取試劑盒:購于生工生物工程(上海)股份有限公司;1,1-二苯基-2-三硝基苯肼(DPPH):Sigma公司;MRS肉湯:北京陸橋技術股份有限公司。
1.1.3 儀器設備 1510酶標儀:美國Thermo公司;SPX-150B-Z生化培養箱:上海博訊實業有限公司;XN-26高速冷凍離心機:美國Beckman Coulter公司。
1.2 試驗方法
1.2.1 乳酸菌鑒定
1.2.1.1 革蘭氏染色 將培養10 h的菌株進行革蘭氏染色鑒定。
1.2.1.2 16SrDNA鑒定 將革蘭氏染色陽性的菌株提取基因組,選用細菌鑒定通用引物,經提取菌株基因組,PCR擴增,瓊脂糖凝膠電泳檢測后寄上海生工生物公司檢測,測序結果與GenBank數據庫比對后繪制系統進化樹(高鵬,2016;呂愛軍等,2010)。
1.2.2 乳酸菌對病原菌的抑制 采用牛津杯法。將分離菌株接種MRS液體培養基,37℃,48 h,取部分發酵液7300×g,4℃離心30 min(郭鳳茹,2019;Komora等,2017)。選取創傷弧菌、大腸桿菌、副溶血性弧菌、哈維氏菌、霍亂弧菌、金黃色葡萄球菌、枯草芽孢桿菌、庫氏鏈霉菌、擬桿菌和嗜水氣單胞菌活化后,稀釋涂布于LB培養基,滅菌牛津杯放置涂菌培養基表面加入200μL發酵上清液,37℃,24 h,測定抑菌圈直徑。
1.2.3 乳酸菌生長特性
1.2.3.1 耐酸性 將菌株接種于用1 mol NaOH和1 mol的HCl調節pH至2.0和3.0的MRS液體培養基中,37℃培養,分別于0、2、4、6 h取樣測定發酵液OD600(杜金城等,2016;Pakarian等,2016),不調節pH的MRS培養基為對照。
1.2.3.2 耐膽鹽 在MRS液體培養基添加0、0.1%、0.3%、0.5%的膽鹽(杜金城等,2016;郝志明等,2006)。接種菌株,37℃培養,分別于0、2、4、6 h取樣測定發酵液OD600。
1.2.4 抗生素對乳酸菌生長的影響 采用藥敏紙片擴散法。菌株100μL菌液涂布MRS培養基,放置數分鐘,選用硫酸卡那霉素、氨芐青霉素、硫酸鏈霉素和氯霉素四種抗生素,藥敏試紙含抗生素量為10μg/mL。將藥敏試紙分別放置于培養基,37℃培養24 h(任士菊等,2014;Newaj-Fyzul等,2014),測定抑菌圈直徑。
1.2.5 抑菌物質的鑒定 分別檢測過氧化氫酶、蛋白酶、pH、加熱處理和超濾對抑菌物質活性的影響,采用牛津杯法,測定上清液抑菌圈的大小(馬迎濤,2016;Ngo,2015)。
1.2.5.1 過氧化氫酶 調節菌株發酵上清液至過氧化氫酶的最適pH 7.0,加入濃度為5 mg/mL的過氧化氫酶,37℃處理2 h,處理后調節pH與對照上清液相同。
1.2.5.2 蛋白酶 菌株發酵上清液分別加入胰蛋白酶、木瓜蛋白酶、蛋白酶K、胃蛋白酶,使其終濃度為5 mg/mL,分別在酶的最適溫度和pH條件下2 h,將上清液pH調至與對照組相同(Papagianni,2015)。
1.2.5.3 pH 菌株發酵上清液的pH調節至3.0、5.0、7.0、9.0,對照為未調節pH。
1.2.5.4 加熱處理 菌株發酵上清液置于60、80、100℃條件下處理15 min,未處理的上清液為對照(遲海等,2020)。
1.2.5.5 超濾 檢測活性物質的分子量。選用3、10、30、50 K和100 K超濾管,取5 mL發酵上清液超濾,4℃,2400×g離心后,取200μL,采用牛津杯法檢測。指示菌為副溶血弧菌、嗜水氣單胞菌。
1.2.6 乳酸菌發酵液的抗氧化活性 菌株發酵上清液、菌懸液及無細胞提取物(CFE)的制備(Li等,2013;Chen等2008)。菌株以1%接種MRS液體培養基,于37℃培養16 h。(1)發酵液4℃條件下,7300×g離心30 min,收集上清液和菌體;(2)菌體經PBS緩沖液洗3次,調整濃度為1.0×1010cfu/mL菌懸液;(3)取部分菌體超聲(1500 W,10 min)破碎,4℃,7300×g離心30 min,上清液為CFE。
1.2.6.1 超氧陰離子自由基的清除能力 在含有0.05 mol的Tris-HCl緩沖液(pH 8.0,4.5 mL)和2.5mol的鄰苯三酚0.4 mL溶液中加入樣品100μL,置于25℃恒溫水浴中放置8 min,立即滴加一滴8 mol HCl終 止反應 (余芳等,2017;Box等,2014)。在325 nm波長測定吸光度。樣品的吸光度:A1,生理鹽水代替樣品:空白對照A0。
超氧陰離子自由基清除率/%=(A0-A1)/A0×100。
1.2.6.2 羥自由基(·OH)的清除能力 在試管中加入的FeSO4溶液(5 mol)2 mL,C7H6O3-C2H5OH(5 mol)2 mL,H2O2溶液(3 mol)2 mL,混合均勻,加入3 mL樣品,37℃保溫20 min,在4℃,7300×g離心10 min,上清液測定OD510數值為Aβ。以無菌生理鹽水代替H2O2為空白,測定的吸光度數值為Aα;生理鹽水代替樣品為對照,測定的吸光度數值為Aε(Das等,2015)。
·OH自由基的清除率/%=[1-(Aβ-Aα)]/Aε×100。
1.2.6.3 DPPH自由基的清除能力 試管中加入樣品和含有0.2 mol DPPH的無水乙醇溶液各2 mL,混均后避光靜置30 min,4℃,7300×g離心10 min,取上清液測定OD517數值為Aγ。空白不含DPPH,記為Aθ,對照為無菌水,記為AΩ(黃玉軍,2013)。
DPPH自 由 基 清 除 率/%=[1-(Aγ-Aθ)]/AΩ×100。
1.2.6.4 還原能力 玻璃試管加入樣品、K3[Fe(CN)6](1%)、pH 6.6的PBS緩沖液各1 mL,混合均勻后(姚杰玢等,2015),50℃保溫15 min,降至室溫后加入1 mL的10%體積的C2HCl3O2,混勻,于4℃,7300×g離心10 min。取上清液、純水和三氯乙酸試劑(0.1%)各2 mL,充分混勻,靜置5min后 測 定OD700(Mikulski等,2020;Wang,2020)。樣品的吸光度值為Aa,以PBS緩沖液代替樣品的吸光度值為Ab。
還原能力/%=(Aa-Ab)/Ab×100。
1.2.7 抗脂質過氧化 取250 g高脂質半滑舌鰨魚飼料樣品碾碎成粉末,加入50 mL培養16 h的菌懸液或發酵上清液,拌勻,空白對照不加。于60℃條件下放置24 h后取出放入500 mL錐形瓶中,加入適量石油醚浸沒樣品,靜置24 h后過濾,置于45℃烘箱內干燥2 h,參考伍立鋒等(2011)的方法測OD572并繪制標準曲線計算過氧化值。
1.3 數據分析 采用三組平行試驗設計,利用Origin 2021進行作圖,應用IBM SPSS Statistics 26進行數據處理與統計,MEGA 7.0繪制系統發育樹。
2 結果與分析
2.1 乳酸菌鑒定
2.1.1 革蘭氏染色結果 經過多輪篩選,共獲得有透明圈的菌株32株。經革蘭氏染色、芽孢染色和鞭毛染色觀察。其中,菌株CS1、CS2、CS4為革蘭氏陽性,桿菌、無芽孢、無鞭毛、菌端鈍圓。結果見圖1。
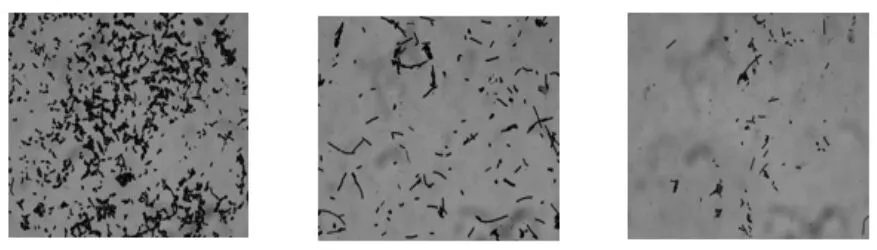
圖1 菌株CS1、CS2、CS4革蘭氏染色結果
2.1.2 菌株16SrDNA的序列分析 三株菌16S rDNA的PCR產物的瓊脂糖電泳見圖2,產物大小約為1500 bp。經上海生工進行測序、比對,構建的系統進化樹見圖3、圖4、圖5。

圖2 16S rDNA PCR產物的瓊脂糖凝膠電泳結果

圖3 菌株CS1系統進化樹

圖4 菌株CS2系統進化樹

圖5 菌株CS4系統進化樹
菌株CS1和CS2與發酵乳桿菌(Lactobacillus fermentum)的同源性為99%;CS4與植物乳桿菌(Lactobacillus plantarum)的同源性為98%,確定三菌株分另為Lactobacillus sp.CS1,Lactobacillus sp.CS2,Lactobacillus sp.CS4。
2.2 菌株對病原菌生長的影響 菌株發酵上清對病原菌生長的影響見表1。菌株CS1對大腸桿菌、維氏氣單胞菌、副溶血弧菌、枯草芽孢桿菌抑菌能力最強,對庫氏鏈霉菌、擬桿菌、嗜水氣單胞菌和金黃色葡萄球菌抑制能力次之,對霍亂弧菌以及創傷弧菌則無明顯的抑制作用。菌株CS2和CS4對維氏氣單胞菌的抑菌能力最強,對創傷弧菌、霍亂弧菌以及幽門螺旋桿菌則無抑制作用,對其余菌株均有不同程度的抑制作用。結果表明,菌株的上清液可抑制病原菌生長,具有廣譜抑菌效果。

表1 菌株上清液的抑菌圈直徑結果
2.3 菌株的生長特性
2.3.1 耐酸性研究結果 菌株CS1、CS2、CS4在pH 2~3條件下均可生長,生長速率略有降低(圖6)。該特性有利于菌株在腸道中的定植。

圖6 菌株CS1、CS2、CS4在不同pH條件下的生長曲線
2.3.2 耐膽鹽研究結果 從圖7可以看出,在0.1%膽鹽濃度條件下,菌株CS1、CS2、CS4均可正常生長。隨著膽鹽濃度的增加,對菌株CS1、CS2、CS4的生長有一定影響。在0.5%膽鹽條件下,菌株的生長速率較正常條件降低40%左右。腸道中膽鹽可以抑制微生物的生長,菌株在不同膽鹽條件下的生長特性有利于其成為腸道優勢菌。

圖7 菌株CS1、CS2、CS4在不同濃度膽鹽下的生長曲線
2.4 藥敏試驗 由表2可知,氨芐青霉素鈉、氯霉素對乳酸菌的生長有顯著的抑制性,顯示出清晰的抑菌圈,而菌株CS1、CS2、CS4對硫酸鏈霉素和硫酸鹽卡那霉素則不敏感。

表2 乳酸菌對抗生素的耐藥性
2.5 抑菌物質活性研究
2.5.1 排除過氧化氫試驗結果 由表3可以看出,經過氧化氫酶處理后的上清液,菌株CS1、CS2、CS4的抑菌圈略有下降,但差異不顯著。結果表明,乳酸菌不是通過產生H2O2抗菌的。

表3 乳酸菌上清液排除過氧化氫試驗結果
2.5.2 蛋白酶對抑菌物質活性影響的試驗結果由表4可知,經蛋白酶K處理后,沒有抑菌活性,經胰蛋白酶處理后,抑菌活性顯著降低。木瓜蛋白酶和胃蛋白酶對上清液的抑菌作用沒有影響。推測上清液中抑菌成分為蛋白質,這與王騰騰(2017)從許氏平鲉腸道中分離的乳酸菌的結果相似。

表4 乳酸菌上清液驗證蛋白性試驗結果
2.5.3 pH對抑菌物質活性的影響 由表5可見,在酸性條件下,菌株CS1、CS2、CS4的發酵上清液對指示菌株的抑制能力沒有顯著差異;在中性條件下,抑菌能力下降;堿性條件時,抑菌能力喪失。

表5 pH對乳酸菌上清液抑菌的影響
2.5.4 抑菌物質熱穩定性研究試驗結果 菌株CS1、CS2、CS4發酵上清液經不同溫度處理后,其抑菌能力均未有顯著變化(表6),表明抑菌物質具有較強的耐高溫能力,短時間在高溫條件下處理,其抑菌能力不受影響。
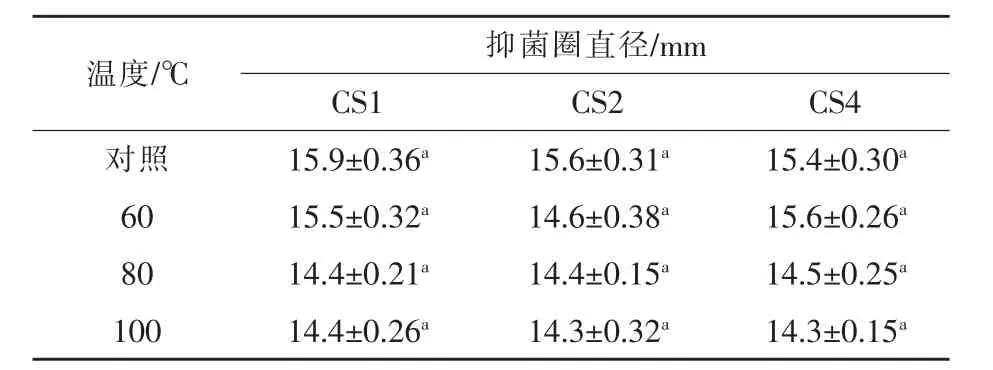
表6 乳酸菌上清液熱穩定性影響的試驗結果
2.6 抑菌物質的抗氧化活性
2.6.1 對超氧陰離子自由基(O2-)的清除能力 由圖8可見,三株試驗菌株的上清液對超氧陰離子的清除能力最強,其中,菌株CS1最強達到(29.8±2.41)%,高于菌株CS2與CS4。三株菌株的無細胞提取物的清除能力弱,表明抗氧化物主要為菌株發酵的胞外產物。與劉洋等(2012)的研究結果相同。
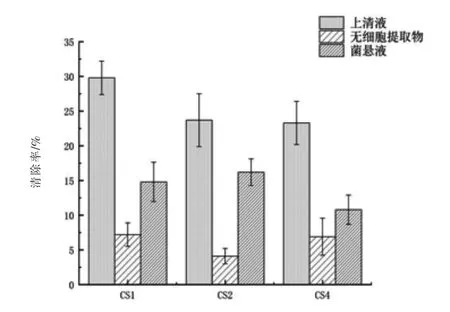
圖8 對超氧陰離子的清除結果
2.6.2 對羥自由基(·OH)的清除能力 由圖9可知,菌株CS1、CS2、CS4的發酵上清液對羥基自由基的清除能力最強,菌懸液次之,無細胞提取物對羥基自由基的清除能力最弱;不同菌株間,CS2的上清液較其他菌株清除能力較強,達 (35.0±1.54)%;對羥基自由的的清除能力強弱則體現了該物質抗氧化能力的大小。

圖9 對羥基自由基的清除結果
2.6.3 對DPPH自由基的清除能力 菌株的發酵上清液對DPPH清除能力最強,其中,菌株CS1達到(52.6±4.10)%,最弱的是菌株CS2,清除能力為(41.7±1.55)%。菌懸液的DPPH清除能力次之,無細胞提取物清除能力最弱。

圖10 對DPPH自由基的清除結果
2.6.4 菌株發酵液的還原能力 從圖11可以看出,菌株發酵上清液的還原能力最強,其中,菌株CS2為(27.5±3.12)%,明顯高于其他兩株菌株。從菌懸液的角度,菌株CS4的還原能力最強,為(20.3±2.35)%,甚至高于其自身上清液。

圖11 菌株發酵液還原性試驗結果
2.7 活性物質分子量估算 選取嗜水氣單胞菌和副溶血弧菌作為指示菌,利用超濾管能將不同分子量物質分離開來的特性,表7和表8顯示,菌株CS1、CS2、CS4的上清液中的抑菌物質主要存在于3~10 kDa,且對于不同病原菌,抑菌物質存在一定差異。結果表明,不同乳酸菌的產物和作用機制有較大的差異,與王偉(2018)試驗結果相同。
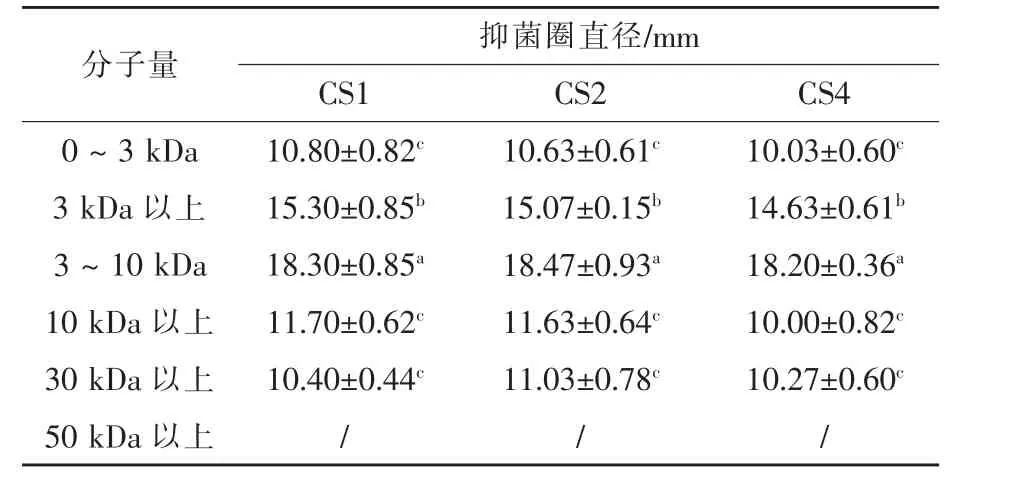
表7 不同分子量范圍的上清液對嗜水氣單胞菌的抑制能力

表8 不同分子量范圍的上清液對副溶血弧菌的抑制能力
2.8 抗脂質過氧化 由圖12可見,與空白相比,加入乳酸菌或其發酵上清的飼料,其抗脂質氧化作用顯著提高,單純加入發酵上清的效果最好。其中,菌株CS4的作用最強。

圖12 乳酸菌菌株的抗脂質過氧化作用
從半滑舌鰨腸道中篩選獲得的乳酸桿菌菌株CS1、CS2和CS4,具有一定的抵抗病害菌的能力,還可以提高飼料的抗脂質過氧化能力,在水產飼料加工中具有一定的應用潛力。
3 結論
從半滑舌鰨腸道中篩選并鑒定了三株乳酸桿菌菌株CS1、CS2和CS4,對病害菌具有一定的抑制能力,其中,對維氏氣單胞菌的作用最強。氨芐青霉素鈉和氯霉素對乳酸菌的生長有顯著的抑制性;相反,菌株對硫酸鏈霉素和硫酸鹽卡那霉素不敏感。通過排除試驗,推測抑菌物質為一種耐熱的蛋白質,分子量在3~10 kDa。菌株在低pH和膽鹽存在條件下可以生長。菌株發酵液具有抗氧化活性,其在飼料中添加可以顯著提高飼料的抗脂質過氧化能力。乳酸桿菌CS1、CS2和CS4在水產養殖以及水產飼料加工中具有一定的應用潛力。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
小讀者(2021年2期)2021-03-29 05:03:48
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
