辣椒適應非生物脅迫的研究進展
2023-01-05 07:48:20胡華冉杜磊張芮豪鐘秋月劉發萬桂敏
生物技術通報 2022年12期
關鍵詞:研究
胡華冉 杜磊 張芮豪 鐘秋月 劉發萬 桂敏
(云南省農業科學院園藝作物研究所,昆明 650205)
辣椒(Capsicum annuumL.)因其豐富的營養價值和良好的風味深受消費者喜愛。除鮮食外,還可干制、醬制和泡制等,加工成辣椒粉、辣椒圈和辣椒酥等調味品,也可深加工提取辣椒素和辣椒紅色素等產品,用于食品、飲料、飼料、保健藥品、化妝品、醫學及軍事等領域[1-2]。根據國家特色蔬菜產業技術體系統計數據,我國辣椒播種面積在蔬菜作物中位居第一,成為我國蔬菜產業中第一大產業[3]。伴隨著辣椒用途的不斷開發和加工型產業的快速發展,辣椒成為我國鄉村振興和精準脫貧的重要抓手。
我國作為辣椒種植大國,對辣椒的研究及開發高度重視。目前,對辣椒的研究主要集中在辣椒種植與栽培、病蟲害防治、辣椒性狀基因定位、辣椒的藥理及風味物質等幾個方面。然而,辣椒在生長發育過程中不僅受到生物脅迫,還受到非生物因子的影響,導致辣椒的品質破壞、產量降低,所以對其抗性方面的研究很有必要。因此,本研究主要總結了近幾年內辣椒在萌發期和苗期響應非生物脅迫的研究進展,分別在形態、生理生化、分子層面以及緩解措施等方面對其研究近況進行綜述,有助于較全面了解辣椒在非生物脅迫條件下的最新研究進展,分析其存在的問題與未來的研究方向,以期為辣椒抗逆品種的培育及開發利用提供參考。
1 溫度脅迫
1.1 辣椒耐高低溫材料的篩選
溫度脅迫包括高溫脅迫和低溫脅迫,危害作物的生長發育,造成作物減產,篩選耐高低溫的品種具有重要的生產意義,相關研究主要集中于以萌發期指標、苗期形態指標和生理生化指標為依據進行篩選。王靜等[4]對20 份辣椒材料的生理指標進行分析,篩選高度耐熱、中度耐熱、不耐熱和極不耐熱4 個等級品種。張慧靜等[5]發現不同制干辣椒品種在脅迫條件下發芽勢、發芽率、發芽指數的差異性顯著,并利用這些指標鑒定品種間的耐冷性。胡能兵等[6]通過生理指標的隸屬度分析法和葉綠素熒光參數降幅的排名獲得可用于抗(耐)高溫的辣椒品種。而通過辣椒資源種子萌發期、幼苗及坐果期的相關形態指標和生理生化特性綜合分析,篩選耐高溫材料的研究也有報道[7]。此外,茍秉調等[8]在低溫弱光脅迫下獲得可用于鑒定和預測辣椒雜種優勢的光合指標,為預測低溫脅迫下的辣椒雜種優勢提供了理論依據。
1.2 生理指標響應
溫度脅迫對辣椒生理生化的影響主要表現在滲透調節物質、保護酶系統、膜脂過氧化和光合作用上。張子學等[9]研究發現,經高、低溫脅迫辣椒葉片中的可溶性糖(soluble sugars,SS)、脯氨酸(proline,Pro)和丙二醛(malondialdehyde,MDA)含量品種之間均存在極顯著差異,其中,SS、Pro 含量大幅度提高、MDA 含量明顯下降的品種,其耐熱或抗寒能力強。潘寶貴[10]研究發現隨著高溫脅迫溫度的升高和脅迫時間的延長,凈光合速率迅速下降,氣孔導度先降后升,超氧化物歧化酶(superoxide dismutase,SOD)活性急劇下降,過氧化氫酶(catalase,CAT)和抗壞血酸過氧化物酶(ascorbate peroxidase,APX)活性也隨之下降,而過氧化物酶(peroxidase,POD)活性持續上升且增加幅度較大,耐熱品種葉片中的酶活性(SOD、POD、APX、CAT)>耐熱性中等的品種>熱敏品種。該結果同何鐵光等[11]、馬寶鵬等[12]結果相似,說明高溫脅迫下,氧化酶活性以及MDA、Pro 含量的變化在不同辣椒品種中有較大的差異,可作為辣椒耐熱鑒定的輔助生理指標。
1.3 抗冷耐熱基因的研究
在形態生理研究的基礎上,學者開始在分子水平上對辣椒進行深入研究,目前已克隆得到多個與溫度脅迫相關的基因。例如,WRKY 家族基因CaWRKY13、CaWRKY27和CaWRKY41,HSP 家族基因CaHSP18、CaHSP24、CaHSP26和HSP90,以及CaERF18、CaERF109、CaSYT5、KCS、NAT、CaVIN02和CaCWIN03,通過生物信息學與表達量分析發現,這些基因對低溫和高溫具有較強的響應[13-21]。有學者對這些基因的機理進行了初步的探索,姬俏華[22]利用農桿菌介導的遺傳轉化方法獲得了CaWRKY20的過表達煙草植株,發現過表達CaWRKY20可顯著提高煙草對高溫的耐受性。顏坤[23]研究發現高溫脅迫下轉正義CaGPAT煙草的光合能力強于野生型。而郭尚敬[24]從甜椒葉片中分離到小分子量熱激蛋白基因CaHSP26和CaHSP18,低溫條件下過量表達對煙草、大腸桿菌均有保護作用。此外,也有學者探討DNA 甲基化在辣椒高溫多濕脅迫反應中的作用[25],以及在轉錄水平和轉錄后水平上闡明CaWRKY40介導的辣椒應答RSI 和HTHH 信號通路相互作用分子機制[26]。Li 等[27]通過比較轉錄組分析揭示了熱脅迫下敏感辣椒和耐熱辣椒的轉錄差異。上述基因在提高植物的溫度脅迫耐受性方面有一定的作用,可用于辣椒抗逆基因工程改良研究。辣椒溫度脅迫相關研究見表1。

表1 辣椒溫度脅迫研究進展Table 1 Recent studies of temperature stress in hot pepper
2 鹽堿脅迫
種子萌發期和苗期是植物對鹽分最敏感的時期,因而辣椒耐鹽性研究主要集中在這兩個時期。研究表明,鹽、堿脅迫使辣椒的發芽勢、發芽率、發芽指數、活力指數及相對鹽害率等指標降低,抑制胚芽及胚根的生長,主根長、側根長與側根分級數以及子葉展開百分比與鹽堿脅迫呈明顯負相關[28-29],幼苗株高、干重和根長顯著下降[30],導致鹽堿脅迫降低了辣椒的產量,影響辣椒果實品質,且堿性鹽的抑制效應大于中性鹽,強堿(Na2CO3)比弱堿(NaHCO3)的抑制作用更強[31-32]。此外,低濃度的中性鹽、堿性鹽對辣椒種子發芽勢均有促進作用。還有學者發現土壤鹽分影響辣椒果實的辣椒素含量[33]。
2.1 辣椒耐鹽性評價研究
植物的耐鹽性有不同程度的表現,且不同辣椒品種在不同指標上表現各有優劣[34-35],目前,大多研究以形態生理指標鑒定品種間或不同地區辣椒材料間的耐鹽性為主[36]。逯明輝等[37]發現,相對發芽勢、相對發芽率和相對發芽指數等3 個指標均能區分不同辣椒品種的耐鹽性,耐鹽性由強到弱依次為尖椒類型>朝天椒類型和線椒類型>甜椒類型。郭春蕊等[38]研究發現,相對發芽指數和相對鹽害率也宜作辣椒發芽耐鹽篩選指標,而株高、地上部鮮重和干重,葉綠素、MDA、相對電導率、酶活性(CAT、SOD、POD)、滲透調節物質SS、Pro、可溶性蛋白(soluble protein,SP)等指標可用來篩選辣椒幼苗的耐鹽性[39-41]。
2.2 生理指標響應
在生理水平上,鹽堿脅迫使滲透調節物質(Pro、甜菜堿、有機酸)含量增加,抗氧化物質抗壞血酸(ascorbic acid,ASA)、還原型谷胱甘肽(reduced glutathione,GSH)、維他命E(vitamin E,VE)增加,保護酶活性(POD、SOD、CAT)增加,K+、Na+含量的變化與膜透性(MDA)的變化一致增加[42]。張弢[43]、吉雪花等[44]研究結果與以上研究基本一致。而鄭佳秋等[45]發現在鹽脅迫下所有供試辣椒品種在幼苗期的SS 和SP 含量總體呈現下降趨勢,不同品種SOD 表現也不同。光合性能指數(如實際光化學效率、表觀光合電子傳遞速率、最大光化學效率和光化學猝滅系數等)均能反應鹽脅迫對辣椒葉片光合性能的影響[46-47]。李漢釗等[48]、劉會芳等[49]發現較高的鹽濃度對辣椒的光合作用及熒光參數有明顯的抑制作用。
2.3 辣椒應對鹽堿脅迫的分子機制探討
目前,有學者對辣椒疑似耐鹽基因CaCBF1A、CabZIP1、CaNAC61、CaCBF1A進行克隆與分析,推測其在辣椒抗逆機制中起著重要的作用[53-55]。馮冬林[56]、尹延旭[57]通過轉基因分別獲得CaERF14、CaPIP1-1的轉基因株系,其對鹽脅迫的耐受力均顯著提高。也有學者通過轉錄組學探索了線辣椒的有關耐鹽機理[58]。由于土壤鹽化和堿化經常同時發生,因此,也有學者從生理和轉錄水平闡明辣椒幼苗對中性鹽、堿性鹽、混合鹽脅迫的差異響應機理。李慧姬[59]對鹽、堿脅迫不同時間點的辣椒幼苗葉片進行轉錄組測序發現,鹽堿差異基因主要富集在光合作用、滲透調節物質合成、氧化還原過程、氮代謝及有機酸代謝等過程。辣椒鹽脅迫相關研究如表2所示。
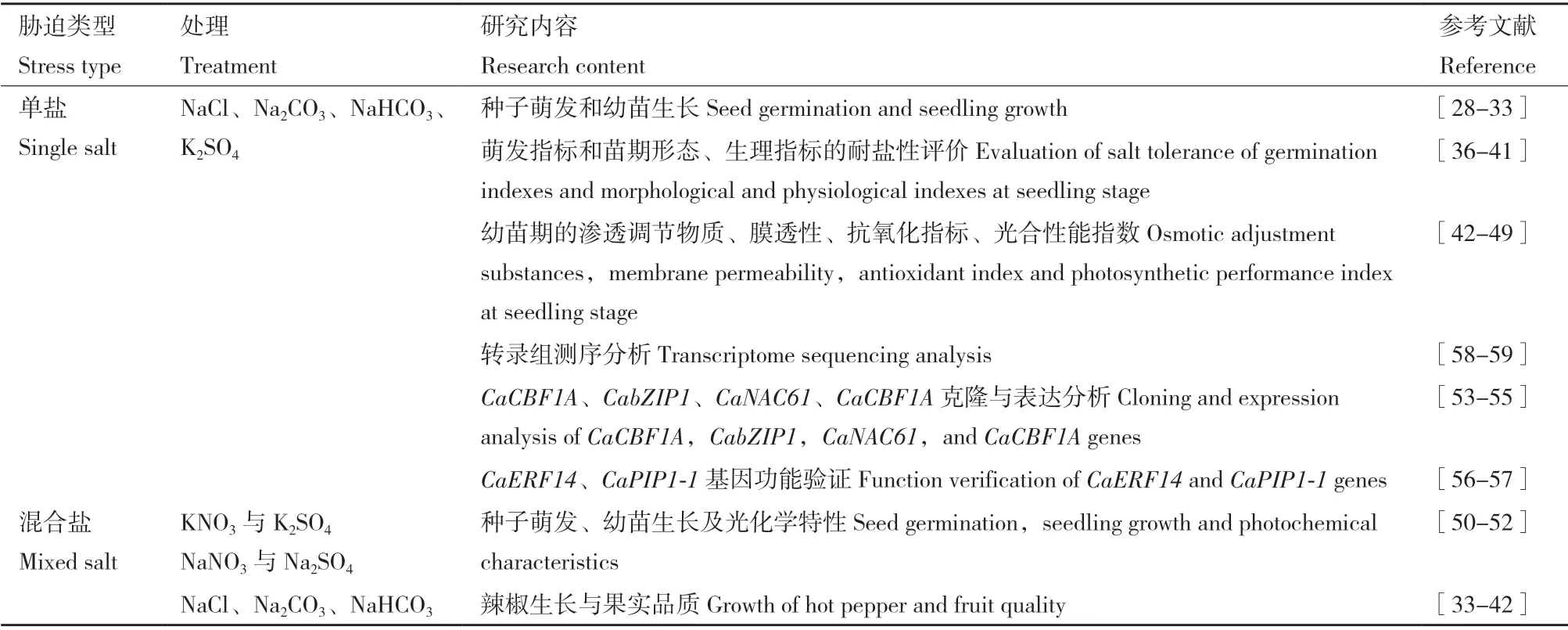
表2 辣椒鹽堿脅迫研究進展Table 2 Recent studies of saline-alkali stress to hot pepper
3 水分脅迫
3.1 干旱脅迫
干旱影響辣椒各階段的生長發育和生理代謝過程,進而影響辣椒的產量和質量[60]。近年來,有關辣椒抗旱脅迫的研究較少,主要集中在辣椒響應干旱脅迫的生理生化特性的研究。有學者研究發現當辣椒植株缺水時,細胞膨壓降低導致細胞伸長生長受到抑制,因而葉片較小,辣椒莖葉形態、根系、根冠比均會受到影響[61-63]。隨著脅迫程度的增加,水勢明顯降低,且細胞內脫落酸含量增高,使凈光合率隨之下降[64]。劉穎等[65]進一步研究發現辣椒地下部分對干旱脅迫的敏感度高于地上部分。朱冉冉[66]研究結果表明辣椒通過提升葉片抗氧化酶活性、Pro 含量、氧化還原酶活性、SP 含量使辣椒抗旱性得到進一步增強,此結果與婁喜艷等[67]、Okunlola 等[68]研究結果一致。總體上抗旱性強的品種組織相對含水量、葉綠素含量和Pro含量相對較高,而質膜透性和MDA 含量相對較低。此外,干旱影響辣椒的果實品質,不僅降低辣椒的果長、果粗、莖粗、株高、單果重以及干物質含量,而且其代謝物質如粗脂肪、粗纖維含量及辣椒素含量較CK 處理顯著下降[69],Ricardez-Miranda 等[70]在水分脅迫對辣椒生長、次生代謝產物和果實產量的研究中也得到類似的結果,但適度干旱可促進光合以及使α-胡蘿卜素、β-胡蘿卜素等上游物質含量顯著增加[66]。
3.2 耐澇脅迫
在淹水條件下,植株有氧呼吸受到抑制,水分難以吸收,蒸騰作用降低,細胞缺水失去膨壓,最終萎蔫。目前,集中于辣椒耐澇脅迫的研究甚少,有學者對耐澇脅迫下辣椒的生理響應進行了研究。Martínez-Acosta 等[71]研究發現淹水處理對植株的氣孔導度、光合和蒸騰速率均產生一定的抑制作用,導致植株的生物量顯著下降。付秋實等[72]研究了水分脅迫對辣椒光合作用及相關生理特性的影響,結果表明,在水分脅迫下辣椒植株總干物質含量、葉片水勢、葉片相對含水量、葉綠素以及類胡蘿卜素含量顯著下降。植物在淹水條件下,保護酶系統會被破壞,自由基就會大量產生,造成膜脂過氧化,MDA 含量大幅增加。劉佳等[73]研究結果表明,隨著處理時間的延長,辣椒葉片中MDA 的含量呈上升趨勢,SOD 和POD 均呈現先升高后降低的趨勢。
有關建立辣椒耐澇性評價體系的研究也有所報道,宋釗等[74]從辣椒受到澇漬脅迫后的植株死亡率、葉片顏色、莖基部淹水處的顏色與形態4 個方面建立辣椒耐澇性鑒定評價體系,并通過篩選高耐澇品種和澇漬敏感品種,證實了這個評價體系有效、可行。辣椒水分脅迫相關研究如表3所示。

表3 辣椒水分脅迫研究進展Table 3 Recent studies of water stress in hot pepper
4 重金屬脅迫
4.1 重金屬對辣椒種子萌發和幼苗生長的影響
目前,由于環境污染,植物受到重金屬毒害的現象極為普遍,國內外學者就植物耐受重金屬脅迫方面進行了大量研究,但對辣椒重金屬耐受性的研究較少,目前的研究主要集中在重金屬鎘(Cd)、鉛(Pb)、鉻(Cr)離子對辣椒種子萌發和幼苗生長的影響。高濃度Cd2+(≥20 mg/L)不僅會降低辣椒種子發芽率,抑制辣椒胚根、胚軸生長,還會造成辣椒幼苗植株矮小,抑制植株生長和根系長度,苗鮮重減少[75];同時Cd2+毒害使辣椒幼苗葉片MDA 含量增加,SOD、CAT 活性下降[76]。有學者還發現辣椒的其他指標,如株高、莖粗、展葉數、葉面積、辣椒葉片的光合指標也會在重金屬處理下降低[77]。此外,低濃度Cd2+、Pb 可促進辣椒植株的生長[78-79]。
4.2 辣椒對重金屬的富集作用
辣椒具有較強的吸收和富集重金屬的能力,因此,有學者研究辣椒不同品種、不同部位、以及不同部位中亞細胞對重金屬的吸收富集作用,多數學者發現辣椒對重金屬(Cd、Pb、As)的富集表現為根部最強[79-81]。楊曉磊等[82]通過植株中Cd 的富集系數研究發現,重金屬的遷移是通過根部吸收后由莖葉轉運,分布濃度依次為根系>莖葉>果實,蘇園[83]研究結果與此一致。此外,品種和土壤不同,辣椒重金屬轉移、富集能力均有所改變。有學者發現辣椒各部位鎘的分布在黃壤上表現為根>莖、葉>果實,石灰(巖)土中為根、葉>果實>莖;不同辣椒類型的鎘含量也存在差異,根部為線椒>朝天椒、雜交椒,葉和果實為朝天椒>線椒、雜交椒[84]。談敏等[85]通過對鎘在辣椒品種間以及辣椒不同器官的富集差異、鎘對辣椒理化性狀的影響、外源物質對辣椒吸收富集鎘的調控4 個方面進行了探討和概述。
4.3 重金屬脅迫相關基因的研究
目前,有關辣椒重金屬脅迫在分子方面的報道較少。劉海波等[86]從辣椒中分離到一個能夠響應熱脅迫的金屬伴侶蛋白基因CaHPP7,其表達受Cu2+、Cd2+等重金屬的誘導;利用基因沉默技術發現辣椒對銅脅迫的抗性都降低;而基因過表達后,銅脅迫下表現為種子發芽率高于對照,幼苗生長的受抑制程度和葉圓片葉綠素含量下降程度均低于對照。故推測CaHPP7在植物應對銅脅迫的過程中起正調控作用。此外,李桃[87]不僅探究了3 個品種辣椒營養器官和果實的生長及生理效應、Cd 吸收、遷移富集及積累情況,也進一步研究了耐性相關基因表達量的變化,初步探明了辣椒低積累Cd 的分子機理,并獲得Cd 積累/耐性過程的關鍵基因。辣椒重金屬脅迫研究進展如表4所示。

表4 辣椒重金屬脅迫研究進展Table 4 Recent studies of heavy-metal stress in hot pepper
5 改良措施
多年來,我國學者不僅在生理和分子水平上對辣椒各種脅迫(溫度、鹽堿、水分、干旱、重金屬)指標進行了系統研究,同時也對如何緩解這些逆境進行了探索。其中,浸種、葉面噴施是常用的方式。
結合自己的“下水”心得和做題情境,我傾向于第三類同學的做法,因為有的放矢確實可以讓閱讀的目光變得更敏銳,提筆標畫更有方向性和側重點(其實關于不動筆墨不讀書的道理不難懂,問題是——非知之艱,惟行之艱),不會出現要么沒重點,要么處處是重點而變成沒重點,耗時耗力,答題效率低下。
5.1 降低溫度脅迫的改良方法
通過葉面噴施不同濃度的5-氨基乙酰丙酸(ALA)、亞精胺(Spd)、氯化鈣(CaCl2)、殼聚糖(CS)、聚乙二醇(PEG)、蔗糖及2,4-表油菜素內酯(2,4-epibrassinolide,EBR),均可以降低辣椒低溫脅迫[88-91]。葉面噴施沼液肥20 倍液可以顯著降低熱害指數[92]。還有學者發現,外源施鈣以及噴施適宜濃度的1-甲基環丙烯或者經過6-BA 處理均可以減少高溫脅迫對辣椒作物帶來的傷害[93-95]。此外,通過嫁接的方式也可以使嫁接苗分別對高溫和低溫脅迫具有較好的適應性[96]。
5.2 降低鹽堿脅迫的改良方法
為緩解鹽脅迫對辣椒的損害,許多學者通過施加外源物質如水楊酸、硝普鈉、油菜素內酯以及通過赤霉素浸種來降低鹽害。有研究發現水楊酸不僅能夠緩解鹽脅迫對辣椒幼苗的生長抑制,促進葉片葉綠素的合成,保護其光合作用,維持植物的正常生長[97-98],還可以增加辣椒幼苗滲透物質的含量和抗氧化酶活性,從而提高其抗鹽能力[99-100]。但超過一定濃度則會抑制辣椒種子的萌發,使種子浸出液電導率和MDA 含量升高[101]。一定濃度赤霉素浸種和硝普鈉處理可以緩解辣椒種子鹽脅迫,提高種子的發芽率、發芽勢和發芽指數,低濃度時有促進作用,高濃度時抑制[102-103]。此外,采用腐植酸和硝酸鈣、生物炭基Rs-198 菌劑也可以減輕鹽脅迫對辣椒造成的危害[104]。
5.3 降低旱澇脅迫的改良方法
已有研究發現通過添加化學試劑水楊酸、一氧化氮、5-氨基乙酰丙酸、茉莉酸甲酯等,均可以提高辣椒抗旱能力[105-107]。而Matmarurat 等[108]通過對辣椒種子進行200 Gy 的γ 射線輻照,以提高辣椒的耐旱性。Neto 等[109]通過使用牛生物肥料、溝槽側覆聚乙烯薄膜、直接土壤覆蓋的方式,來緩解干旱地區的水分蒸發問題。結果表明,在甜椒栽培中,使用溝槽側覆可以提高產量,提高生產率;但牛生物肥料×側覆×土壤覆蓋的相互作用會抑制植物中葉綠素的產生。另外,通過外源Ca2+浸種處理,調節辣椒幼苗根系內呼吸代謝可以緩解淹水脅迫對植株的傷害,改善根系(根系總長度、總表面積、體積、根尖數)的生長狀況,提高根系活力[110-111]。
5.4 降低重金屬脅迫的改良方法
目前,土壤重金屬污染的治理包括工程法、植物修復法和農藝調控法等。部分學者提出施加組配改良劑來改善辣椒對重金屬的吸收。例如,施加不同添加量的組配改良劑海泡石+石灰石后,與對照相比,辣椒可食部位和根部重金屬Pb、Cd、Cu、Zn 含量明顯降低[112]。葉面噴施不同濃度的Si、P、Zn,會緩解鎘對辣椒的毒害,促進其生長[113]。此外,在鎘或鋁環境下,施用一定濃度的有機肥、雙氰胺和石灰、外源鈣,可以緩解辣椒植株的金屬脅迫,并提高辣椒的果實品質[114-115]。
通過以上文獻分析發現,緩解辣椒非生物脅迫的措施存在一個共同點,在非生物脅迫下,外源物質處理的辣椒幼苗抗逆性與葉綠素含量、保護酶活性和滲透調節物質的含量均有密切關系,可以通過測定生長指標(苗高、莖粗、根鮮重、根干重、地上部鮮重、地上部干重等)和生理指標(SS、SP、Pro、MDA、SOD 和POD 等)來鑒定外源物質對提高辣椒幼苗抗逆的作用效果,獲得其生理改良機制。辣椒非生物脅迫的改良方法如表5所示。

表5 辣椒非生物脅迫的改良方法Table 5 Improved methods of abiotic stress in pepper
6 展望
我國是世界第一大辣椒(含甜椒)生產國與消費國,近年來全球氣候變化對辣椒生產帶來的影響不容忽視,因此,加強辣椒抗逆性方面的研究具有重要的意義。截至目前,關于辣椒非生物脅迫的研究雖然已經取得了一定的進展,但與其他重要的農作物相比,仍然差距較大,今后可從以下幾個方面開展研究工作。
在提高辣椒對逆境的耐受能力方面,我國研究相對薄弱,尤其是通過探索植物的抗逆機理并利用現代生物技術,改變其遺傳基礎來提高耐受性方面仍處于初級階段。目前,已經利用現代分子生物技術等手段,從辣椒中發掘到一些抗逆相關基因,下一步則需著重闡明其在植物體內行使的功能并對此基因進行克隆并導入其他高產辣椒,從而使辣椒間的優良基因結合,最終培育出品質好、產量高、抗性強的辣椒新品種。但受品種遺傳性、環境因素、激素配比、操作方法等因素的影響,辣椒組培體系并未十分成熟,難以建立完善的轉化體系,因此,抗逆基因的導入任務有一定困難,需要加強辣椒抗性基因的挖掘與利用。
辣椒的生長受多因素的影響,其響應變化是相互協同或者相對獨立仍需進一步思考。有報道稱,辣椒屬于干旱敏感、適度耐鹽的蔬菜作物,其生長發育容易受到干旱以及由過度施肥造成的鹽分脅迫影響。有關鹽堿、溫度及水分脅迫兩兩互作或三者互作對辣椒生長發育在生理水平的影響已有相關研究,但在分子層面上對其進行深入研究的報道并不多,對辣椒抗逆性機理的研究可以作為后續的一個研究方向。同時在機理研究的基礎上,篩選與辣椒抗逆性有關的分子標記或基因,從而加快辣椒抗逆性品種的選育。
不同生育期的辣椒對非生物脅迫的響應程度不同,目前大部分的研究都集中在對苗期的影響上,而對成株期影響的研究較少,特別是對采收期辣椒果實產量、品質如何響應非生物脅迫缺少全面的認識,所以還需要進一步的研究。
通過前面綜述可知,關于辣椒逆境脅迫條件下激素水平變化規律及其對耐澇、重金屬脅迫的響應的研究不僅數量少,而且多數停留在生理水平。因此,需要進行調控機理的深入研究,進一步為辣椒抗性育種的親本選擇提供理論依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
