茶樹(shù)生長(zhǎng)光調(diào)控研究進(jìn)展
2023-01-11 07:42:52蓋淑杰王奕雄李蘭劉碩謙李銀花程孝夏茂劉仲華周智
茶葉科學(xué) 2022年6期
關(guān)鍵詞:影響
蓋淑杰,王奕雄,李蘭,劉碩謙,李銀花,程孝,夏茂,,劉仲華*,周智,*
1. 湖南農(nóng)業(yè)大學(xué)湖南省光學(xué)農(nóng)業(yè)工程技術(shù)研究中心,湖南 長(zhǎng)沙 410128;2. 湖南農(nóng)業(yè)大學(xué)茶學(xué)教育部重點(diǎn)實(shí)驗(yàn)室,湖南 長(zhǎng)沙 410128;3. 湘豐茶業(yè)集團(tuán)有限公司,湖南 長(zhǎng)沙 410128
茶是世界公認(rèn)的健康飲料,具有增強(qiáng)神經(jīng)興奮、消食利尿等養(yǎng)生保健功能[1]。中國(guó)是世界最大的茶葉生產(chǎn)國(guó)和消費(fèi)國(guó),擁有豐富的茶樹(shù)資源和茶產(chǎn)品[2]。茶葉品質(zhì)受地理位置、栽培措施、氣候條件等因素的影響[3],環(huán)境的改變直接影響茶樹(shù)的生長(zhǎng)與葉片外形及內(nèi)含成分的變化[4-5]。不同環(huán)境因子對(duì)茶樹(shù)表現(xiàn)出明顯的調(diào)控差異,其中光調(diào)控效應(yīng)較為顯著[6]。光是影響植物光合作用的關(guān)鍵因子,也是維持植物生命的能量來(lái)源[7]。光對(duì)作物生長(zhǎng)發(fā)育、光合作用、物質(zhì)代謝以及基因表達(dá)均具有調(diào)控作用[8],光強(qiáng)、光質(zhì)和光周期不僅可以影響植物的生物學(xué)基礎(chǔ),也是植物形成優(yōu)良品質(zhì)的必要條件[9]。已有研究表明,茶樹(shù)的器官與形態(tài)、細(xì)胞結(jié)構(gòu)與代謝物組分等都受光照影響,且不同光強(qiáng)、光質(zhì)、光周期作用于茶樹(shù)上效果不同[10]。本文總結(jié)了茶樹(shù)生長(zhǎng)發(fā)育、光合作用、次生代謝產(chǎn)物等在不同光強(qiáng)、光質(zhì)、光周期下的變化情況,旨在為開(kāi)發(fā)可用于作物生產(chǎn)實(shí)踐的光調(diào)控技術(shù)提供理論支撐。
1 茶樹(shù)生長(zhǎng)發(fā)育光調(diào)控
1.1 茶樹(shù)形態(tài)建成
茶樹(shù)生長(zhǎng)受光照影響,光照強(qiáng)度會(huì)影響茶苗的生長(zhǎng)及形態(tài)。過(guò)度遮光(遮光率95%~98%)會(huì)推遲茶苗出土?xí)r間,降低茶苗出土成活率;遮光度高(遮光率 70%~90%)則會(huì)使茶苗莖桿變細(xì)、發(fā)育不良;而強(qiáng)烈自然光直接照射也會(huì)顯著抑制節(jié)間的生長(zhǎng),因此茶苗生長(zhǎng)需要適宜的光強(qiáng)[11]。在不同光質(zhì)下,茶苗的生長(zhǎng)也存在差異。Homma等[12]研究發(fā)現(xiàn),利用不同波長(zhǎng)(峰值波長(zhǎng),藍(lán)色465 nm、綠色502 nm、黃色592 nm、紅色660 nm)的LED光連續(xù)照射茶苗,發(fā)現(xiàn)紅色和綠色 LED照射的生根插穗的芽表現(xiàn)出良好的生長(zhǎng)狀態(tài),原因是紅色LED會(huì)激活光合作用,而綠光影響機(jī)制尚不清楚,需要進(jìn)一步試驗(yàn)驗(yàn)證,說(shuō)明光質(zhì)是影響茶樹(shù)生長(zhǎng)的重要條件之一。
扦插繁殖可將茶樹(shù)優(yōu)良性狀穩(wěn)定遺傳到子代[13],而光照可以影響茶樹(shù)扦插繁殖效率。周漢忠[14]研究表明,中度遮光(遮光率70%~80%)條件下,插穗生根時(shí)間短,速度快,根群系發(fā)育良好。龔自明等[15]研究發(fā)現(xiàn),采用180 cm高平式黑色遮陽(yáng)網(wǎng)套40 cm高弧形無(wú)色透明薄膜進(jìn)行遮陰,可以提高扦插成活率、出圃率及平均苗高。劉任堅(jiān)等[16]也發(fā)現(xiàn),弱光和強(qiáng)烈直射光不利于茶樹(shù)插穗生根和存活,光照為自然光強(qiáng)的50%時(shí),可提高茶苗插穗存活率和生根率。同時(shí),也有研究表明,茶樹(shù)扦插過(guò)程中采用黃色塑料薄膜遮陰較紅色和黑色薄膜遮陰,對(duì)茶苗生根的促進(jìn)作用更明顯[17]。光照時(shí)間對(duì)插穗也有影響,延長(zhǎng)光照時(shí)間會(huì)促進(jìn)茶樹(shù)插穗的生長(zhǎng),其生根率、生根數(shù)均顯著增加;生根后,隨著光照時(shí)間的延長(zhǎng),新梢長(zhǎng)、新生葉片數(shù)、莖粗增加,光照時(shí)間在晝夜比為16 h/8 h時(shí)最佳[18]。
1.2 茶樹(shù)葉片結(jié)構(gòu)
茶樹(shù)生長(zhǎng)在不同的光照環(huán)境下,其葉片結(jié)構(gòu)存在差異[19]。左辭秋等[20]研究顯示,隨著光照強(qiáng)度的增加,茶樹(shù)葉片及表層細(xì)胞增厚、細(xì)胞間隙變小;光照強(qiáng)度降低時(shí)柵欄細(xì)胞末端變尖。張露荷等[21]研究發(fā)現(xiàn),遮陰后黃金芽葉片的柵欄組織層數(shù)增加且排列更整齊緊密。茶樹(shù)葉片組織木質(zhì)化也受到光照調(diào)控,Teng等[22]研究表明,進(jìn)行 80%的遮陰會(huì)降低茶樹(shù)葉片的木質(zhì)化程度,其原因可能是遮陰處理負(fù)調(diào)控木質(zhì)素生物合成相關(guān)基因(CsPAL和Cs4CL)的表達(dá),而CsPAL和Cs4CL與木質(zhì)素水平呈強(qiáng)正相關(guān)。
2 茶樹(shù)光合生理特性光調(diào)控
2.1 光合色素
高等植物中的光合色素主要有葉綠素 a(Chlorophyll a)、葉綠素 b(Chlorophyll b)和類胡蘿卜素(Carotenoids)。其中,葉綠素是植物葉綠體內(nèi)參與光合作用的重要色素,能夠捕捉光能并將其進(jìn)行轉(zhuǎn)化和重新導(dǎo)向。研究表明,光照強(qiáng)度的差異會(huì)影響茶樹(shù)光合色素的合成,進(jìn)而引起葉色的變化[23]。遮陰作為改變光強(qiáng)的重要手段,影響茶樹(shù)葉片中色素的合成。在遮陰條件下,葉綠素生物合成途徑中的結(jié)構(gòu)基因(HEMA1、PORA和CLH1等)和參與光信號(hào)傳遞的潛在成分基因(PHYA、HY5和CRY1等)的轉(zhuǎn)錄水平發(fā)生了顯著變化,細(xì)胞核和葉綠體之間的兩個(gè)中央信號(hào)整合子(GLK1和LHCB)對(duì)遮陰有明顯反應(yīng),促進(jìn)了葉綠素的合成[24]。也有研究表明,茶樹(shù)葉片中葉綠素的高積累可能是通過(guò)抑制CsHY5的表達(dá),阻止其抑制下游基因CsPORL-2的表達(dá)[25]。此外,適度遮陰可增加非甲戊酸途徑基因(CsDXS1和CsDXS3)以及類胡蘿卜素合成途徑中關(guān)鍵基因(CsPSY、CsLCYB和CsLCYE)的表達(dá),進(jìn)而提高葉片類胡蘿卜素總量[26]。
不同光質(zhì)下光合色素的合成也有差異。田月月[27]在光強(qiáng)為 150 μmol·m-2·s-1時(shí),使用不同光質(zhì)處理黃金芽,發(fā)現(xiàn)紅光可誘導(dǎo)葉片中與捕光色素蛋白相關(guān)基因的表達(dá),促進(jìn)葉綠素的合成,但藍(lán)光顯著下調(diào)該基因的表達(dá),并通過(guò)誘導(dǎo)葉片中參與脫落酸合成和葉綠素降解基因的表達(dá)降低葉綠素和類胡蘿卜素的合成。Tian等[28]推測(cè),紅光可以強(qiáng)烈誘導(dǎo)黃金芽中CsPHYA和CsPHYB的表達(dá),促進(jìn)葉綠素的合成。
2.2 光合作用
茶樹(shù)在其生命周期中產(chǎn)生的全部有機(jī)物質(zhì)的碳骨架都由光合作用提供,光照則能為光合作用提供輻射能[29]。Hajiboland等[30]研究表明,不同年齡的茶樹(shù)葉片利用光合作用吸收的碳會(huì)合成不同類型的化合物,影響茶葉的品質(zhì);而且隨著茶樹(shù)嫩芽逐漸成熟,凈光合速率(Pn)逐漸增加,在芽準(zhǔn)備采收時(shí)達(dá)到最大值,茶樹(shù)成熟葉的凈光合速率也隨光強(qiáng)的增加呈漸近響應(yīng),溫室試驗(yàn)發(fā)現(xiàn),幼葉吸收CO2的最佳光強(qiáng)(250 μmol·m-2·s-1)低于老葉(500 μmol·m-2·s-1),說(shuō)明光照不僅影響茶樹(shù)光合作用,也會(huì)間接影響茶葉品質(zhì)。Xia等[31]研究證實(shí),在較高光強(qiáng)下(411 μmol·m-2·s-1),茶樹(shù)葉片蒸騰速率和氣孔導(dǎo)度值通常較大。但Mohotti等[32]證明,光強(qiáng)超過(guò) 1 400~1 500 μmol·m-2·s-1時(shí),Pn 會(huì)降低,原因是強(qiáng)光抑制了光合作用中補(bǔ)光色素天線蛋白等與光反應(yīng)相關(guān)蛋白的下調(diào),進(jìn)而抑制了葉片的光合電子傳遞效率,而在適度遮陰下生長(zhǎng)的茶樹(shù)光合速率高于正常光照,且比正常光照更容易達(dá)到光飽和點(diǎn)[33]。
不同光質(zhì)對(duì)茶樹(shù)光合作用也有影響。雜交品種福云6號(hào)在冬季光照為120 lx的條件下,較藍(lán)光(430 nm)和紫光(400 nm)而言,補(bǔ)充紅光(660 nm)可加快光合速率,促進(jìn)光合產(chǎn)物的積累,同時(shí)可使環(huán)境溫度提升[34]。Suzuki等[35]試驗(yàn)表明,在光強(qiáng)控制為 100 μmol·m-2·s-1時(shí),水培茶樹(shù)在藍(lán)光(450 nm)照射下促進(jìn)了氣孔開(kāi)口,并且伴隨蒸騰速率的加快和光合速率的上升,促進(jìn)了氮的吸收同化。Zheng等[36]研究表明,夜間補(bǔ)充藍(lán)光、綠光可有效改善低光照下茶樹(shù)的生長(zhǎng)和光合能力,引發(fā)活性氧和激素信號(hào)對(duì)光合作用的調(diào)控。說(shuō)明茶樹(shù)光合作用受光質(zhì)與其他環(huán)境因子協(xié)同調(diào)控,不同品種之間存在差異性。
2.3 葉綠素?zé)晒鈪?shù)
通過(guò)葉綠素?zé)晒夥治黾夹g(shù),可以更直觀了解光調(diào)控對(duì)植物光合生理狀況的影響[37]。其中表觀量子效率(AQY)和光系統(tǒng)Ⅱ(Photosystem Ⅱ complex,PSⅡ)光化學(xué)效率(Fv/Fm)的降低是植物發(fā)生光抑制的明顯特征,在其他環(huán)境因子一定時(shí),光照使茶樹(shù)葉片的AQY和Fv/Fm下降,表明抑制了光合作用。研究也表明,隨著光照強(qiáng)度的增加,PSⅡ的Fv/Fm顯著降低,高光強(qiáng)會(huì)造成茶樹(shù)光抑制以及對(duì)PSⅡ的嚴(yán)重?fù)p害[38]。張露荷等[39]通過(guò)遮陰降低光強(qiáng),發(fā)現(xiàn)遮陰可以顯著提高黃金芽的Fv/Fm,同時(shí)降低葉片的光化學(xué)淬滅系數(shù)(qP),驗(yàn)證了過(guò)量的光強(qiáng)會(huì)造成光抑制。韋朝領(lǐng)等[40]試驗(yàn)發(fā)現(xiàn),生長(zhǎng)在飽和光強(qiáng)以下的茶樹(shù),突然遇到強(qiáng)光后Fv/Fm顯著降低,但在恢復(fù)一段時(shí)間后回到正常值,這說(shuō)明強(qiáng)光產(chǎn)生的光抑制雖然較嚴(yán)重,但并沒(méi)有對(duì)光合機(jī)構(gòu)造成不可逆轉(zhuǎn)的破壞。Sano等[41]發(fā)現(xiàn),茶樹(shù)遮陰后突然暴露于強(qiáng)光下會(huì)導(dǎo)致Fv/Fm降低,羰基化蛋白水平升高,說(shuō)明Fv/Fm的降低可能反映了茶樹(shù) PSⅡ作為強(qiáng)光防御機(jī)制會(huì)下調(diào)。因此葉綠素?zé)晒庾兓梢宰鳛楣庹{(diào)控茶樹(shù)生長(zhǎng)效果的評(píng)判標(biāo)準(zhǔn)之一。
綜上所述,光調(diào)控會(huì)引起茶樹(shù)葉片光合生理特性的變化(表1),茶樹(shù)葉片會(huì)自主對(duì)環(huán)境進(jìn)行適應(yīng),做出一系列應(yīng)激反應(yīng)以保護(hù)自身不受強(qiáng)光損害,適應(yīng)生長(zhǎng)。
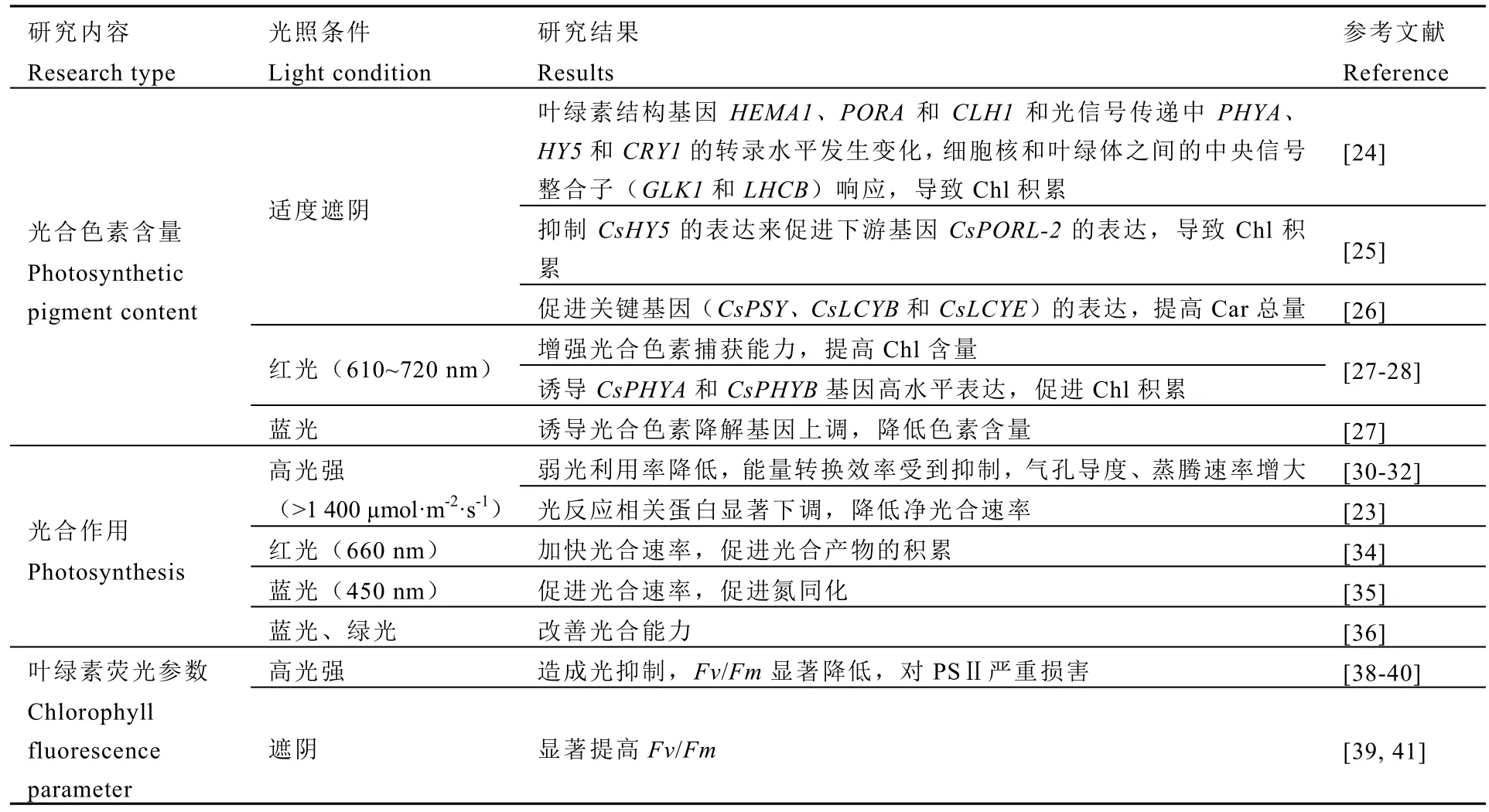
表1 光照對(duì)茶樹(shù)光合特性的影響Table 1 Effects of light on photosynthetic characteristics of tea plants
3 茶樹(shù)主要代謝產(chǎn)物光調(diào)控
茶葉品質(zhì)及茶葉風(fēng)味會(huì)受到光調(diào)控的影響,因?yàn)椴煌庹諚l件下茶樹(shù)的代謝途徑和代謝產(chǎn)物組分發(fā)生了變化[42]。
3.1 茶多酚
茶多酚(Tea polyphenols)是茶葉中多酚類物質(zhì)的總稱,主要分為兒茶素類、花色素類、黃酮及黃酮醇類、酚酸及縮酚酸類四大類[43]。光照可調(diào)控茶樹(shù)中多酚類物質(zhì)組分,其中紫外線對(duì)其作用效果顯著(圖1)。Lin等[44]試驗(yàn)得出,在低強(qiáng)度UV-B(20 μW·cm-2)下照射2 h可刺激茶樹(shù)中部分兒茶素的積累,使總兒茶素水平增加;而在高強(qiáng)度UV-A(300 μW·cm-2)和 UV-B(600 μW·cm-2)下照射 8 h則會(huì)抑制茶樹(shù)中兒茶素的積累;在適當(dāng)?shù)淖贤饩€照射下,EGCG比其他兒茶素增加得更快,這可能與茶樹(shù)中的CsHY5與CsMYB12基因相互作用,介導(dǎo)UV-B信號(hào),通過(guò)直接結(jié)合CsFLS、CsDFRa和CsLARa啟動(dòng)子中的Gbox結(jié)構(gòu)域促進(jìn)兒茶素等黃酮類化合物的合成有關(guān)(圖1-A)。Liu等[45]試驗(yàn)證明,遮陰可使茶樹(shù)中黃酮醇和黃烷醇減少,因?yàn)檎陉帡l件下 UVR8同源二聚體的解離受到限制,導(dǎo)致HY5的穩(wěn)定性降低,抑制了黃酮類生物合成途徑中下游響應(yīng)基因(FLS、CHSs和F3'H等)的激活,因此減少了黃酮醇等的合成;同時(shí),UVR8的減少可能通過(guò)MYB4的調(diào)節(jié)(激活)導(dǎo)致黃酮醇(C4H、4CL和CHS等)和兒茶素(LAR和ANR等)相關(guān)基因的表達(dá)量下調(diào),最終影響黃酮醇和兒茶素的產(chǎn)生(圖1-B)。遮陰也是影響多酚類物質(zhì)組分的重要手段之一,Zhao等[46]剖析了遮陰條件下黃酮醇生物合成的綜合調(diào)控網(wǎng)絡(luò),發(fā)現(xiàn)遮光使CsbZIP1失活以及CsPIF3激活,進(jìn)一步激活了MYB抑制基因CsMYB4和CsMYB7,最終導(dǎo)致黃酮醇的合成受抑制。同時(shí)Xu等[47]研究表明,遮陰還可以改變茶樹(shù)全基因組DNA甲基化模式,進(jìn)而促進(jìn)酯型兒茶素向非酯型兒茶素的轉(zhuǎn)化。而且遮陰會(huì)使部分兒茶素類化合物(CG、EC)、總黃酮的含量降低,且在 24 h全黑暗處理下,參與類黃酮化合物代謝過(guò)程的必需基因下調(diào)也會(huì)導(dǎo)致類黃酮化合物的含量降低[48]。Wang等[49]通過(guò)對(duì)類黃酮生物合成途徑關(guān)鍵基因的表達(dá)進(jìn)行分析,發(fā)現(xiàn)參與兒茶素生物合成的花青素還原酶的兩個(gè)同源基因ANR1和ANR2在遮陰條件下都有不同程度的下調(diào),遮陰處理可以抑制茶樹(shù)類黃酮和木質(zhì)素的生物合成。Zhang等[50]研究發(fā)現(xiàn),遮陰抑制茶樹(shù)中黃酮類化合物的積累,認(rèn)為可能與苯丙烷/類黃酮途徑的上游和下游分支存在底物競(jìng)爭(zhēng)有關(guān),并且受光強(qiáng)和溫度的影響。在黑網(wǎng)和不同顏色棚膜遮陰下,黃酮醇苷(TFG)前體產(chǎn)物的減少和兒茶素生物合成分支的增強(qiáng)導(dǎo)致 TFG含量顯著下降,表現(xiàn)出了對(duì)光照強(qiáng)度的高敏感性,其苷元部分(M-苷和 Q-苷)的響應(yīng)普遍較大,研究也表明了紫外線強(qiáng)度與黃酮苷生物合成基因的相關(guān)性更強(qiáng),紫外線可以通過(guò)上調(diào)參與花青素生物合成的結(jié)構(gòu)和調(diào)節(jié)基因來(lái)誘導(dǎo)花青素的積累,輻射抑制了部分類黃酮合成酶的表達(dá)水平,導(dǎo)致花青素還原酶(ANR)活性降低,向花青素生物合成的代謝通量轉(zhuǎn)移[51]。說(shuō)明光照調(diào)控類黃酮化合物代謝是多途徑的。與此同時(shí),Wang等[5]試驗(yàn)證明,適當(dāng)提高藍(lán)光照射的強(qiáng)度,可以改善茶樹(shù)的轉(zhuǎn)錄重編程和代謝通量重定向,從而調(diào)節(jié)茶樹(shù)的多酚物質(zhì)含量。王加真等[52]通過(guò)不同光強(qiáng)對(duì)茶樹(shù)進(jìn)行調(diào)控得出,光強(qiáng)為 200 μmol·m-2·s-1時(shí)對(duì)茶樹(shù)葉片中功能成分的積累影響較大,有利于提高茶多酚含量和酚氨比。由此可見(jiàn),通過(guò)遮陰來(lái)調(diào)節(jié)光照或直接調(diào)節(jié)光強(qiáng),都會(huì)對(duì)茶樹(shù)中多酚類物質(zhì)的組分有影響。
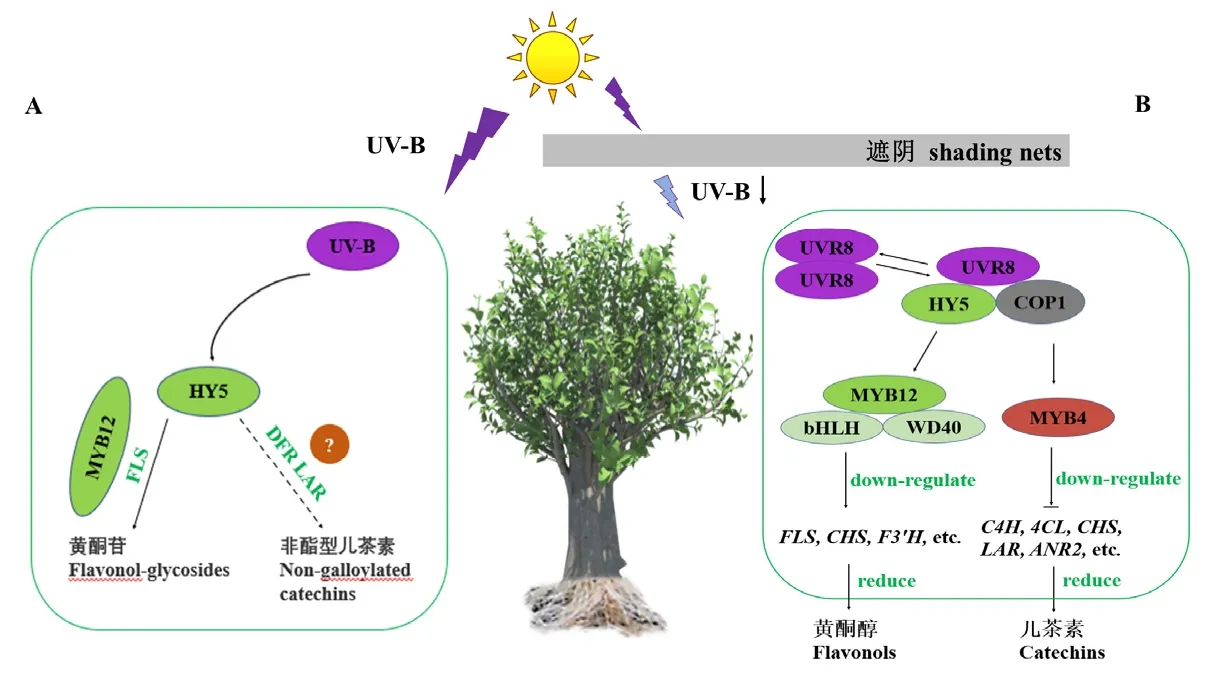
圖1 紫外線條件對(duì)茶樹(shù)中多酚類物質(zhì)含量的影響[44-45]Fig. 1 Effect of Ultraviolet light on polyphenol contents in tea plants[44-45]
光質(zhì)的改變也會(huì)對(duì)類黃酮化合物的組分產(chǎn)生影響。李麗田等[53]研究發(fā)現(xiàn),在藍(lán)色薄膜遮光處理下中茶 108新梢中茶多酚及兒茶素的合成受到抑制,而在紅色、黃色、紫色薄膜遮光處理下中茶 108新梢中茶多酚及兒茶素含量變化不明顯。但Zheng等[36]研究認(rèn)為,藍(lán)光(460 nm)能誘導(dǎo)中黃 3號(hào)中CRY2/3、SPAs、HY5、R2R3-MYBs基因的表達(dá),促進(jìn)茶樹(shù)花青素和兒茶素的積累。藍(lán)光環(huán)境下的結(jié)果差異性可能是藍(lán)光波段不同以及茶樹(shù)品種的差異所導(dǎo)致。綠光(520 nm)則能抑制CRY2/3和PHOT2基因介導(dǎo)的藍(lán)光反應(yīng),影響花青素和兒茶素的生物合成,并且可以刺激功能性物質(zhì)的積累(如B2原花青素/B3和L-抗壞血酸)[54]。吳慶東[55]研究表明,500~780 nm波長(zhǎng)的光可以促進(jìn)茶樹(shù)新梢中茶多酚的積累,黃酮醇苷生物合成的關(guān)鍵酶(F3050H)在紅光下顯著上調(diào),EGCG含量在紅光下可以高積累[56]。以上研究表明,多酚類物質(zhì)對(duì)不同光質(zhì)的響應(yīng)存在差異。
光周期對(duì)茶樹(shù)中的酚類物質(zhì)也有影響。張澤岑等[57]研究表明,延長(zhǎng)白光照射,可以提高茶樹(shù)愈傷組織培養(yǎng)中酚類化合物的合成。Shamala等[58]發(fā)現(xiàn),茶樹(shù)暴露于UV-B下30 min,葉片中大多數(shù)兒茶素類物質(zhì)持續(xù)會(huì)增加,但長(zhǎng)期暴露于紫外光下(2~8 h),其含量則受到顯著抑制。Wang[59]等通過(guò)分析不同時(shí)段茶鮮葉內(nèi)成分變化得出,白天茶樹(shù)總兒茶素和主要兒茶素的積累量均高于傍晚,是因?yàn)榘滋斓母吖庹蘸筒铇?shù)活躍的光合作用利于其積累,并且發(fā)現(xiàn)茶樹(shù)中兒茶素隨晝夜節(jié)律的變化是光照、溫度和濕度等多種環(huán)境因子共同作用的結(jié)果。
3.2 氨基酸
茶葉中的氨基酸是影響其品質(zhì)的重要因子,其中茶氨酸(L-Theanine)是茶葉中含量最高的游離氨基酸[60]。茶樹(shù)中氨基酸的組分也會(huì)受到光照等環(huán)境因子的影響。
大量研究表明[61-63],遮陰處理可以增加茶樹(shù)葉片中茶氨酸的含量(圖2)。有關(guān)遮陰程度的影響,王玉花等[64]在春季、夏季、秋季,通過(guò)設(shè)置不同程度遮光,發(fā)現(xiàn)56%遮光率的黑色遮陽(yáng)網(wǎng)遮光可使氨基酸含量大幅增加,酚氨比最低;胡永光等[65]在春茶采摘末期進(jìn)行了60%和40%遮光處理,發(fā)現(xiàn)遮光率60%的遮陽(yáng)網(wǎng)處理下茶葉的酚氨比降低,鮮爽味提高。陸安霞等[66]對(duì)茶樹(shù)進(jìn)行53%和70%的遮光處理,發(fā)現(xiàn)在前期可起到提高茶葉中氨基酸含量的作用,但是隨著遮光時(shí)間的延長(zhǎng),作用逐漸減弱,說(shuō)明遮光時(shí)間對(duì)于氨基酸含量也存在影響。陳建姣等[42]研究發(fā)現(xiàn),湘妃翠和黃金茶1號(hào)分別在 95%和 80%的遮陰度處理下,茶葉茶氨酸含量高且品質(zhì)較好,但在短期復(fù)光后其含量降低。傅海平等[67]研究發(fā)現(xiàn),遮陰處理6 d明顯促進(jìn)了保靖黃金茶 1號(hào)中游離氨基酸的積累,而且雙層黑色遮陰網(wǎng)處理的效果優(yōu)于單層黑色遮陰網(wǎng),但隨遮陰時(shí)間延長(zhǎng),游離氨基酸總量變化不明顯。以上結(jié)果表明,遮陰處理可以提高茶樹(shù)氨基酸含量并降低酚氨比,但是研究并未對(duì)不同遮陰程度和遮陰時(shí)間進(jìn)行系統(tǒng)性比較,未得出普遍性規(guī)律,還有待下一步研究。
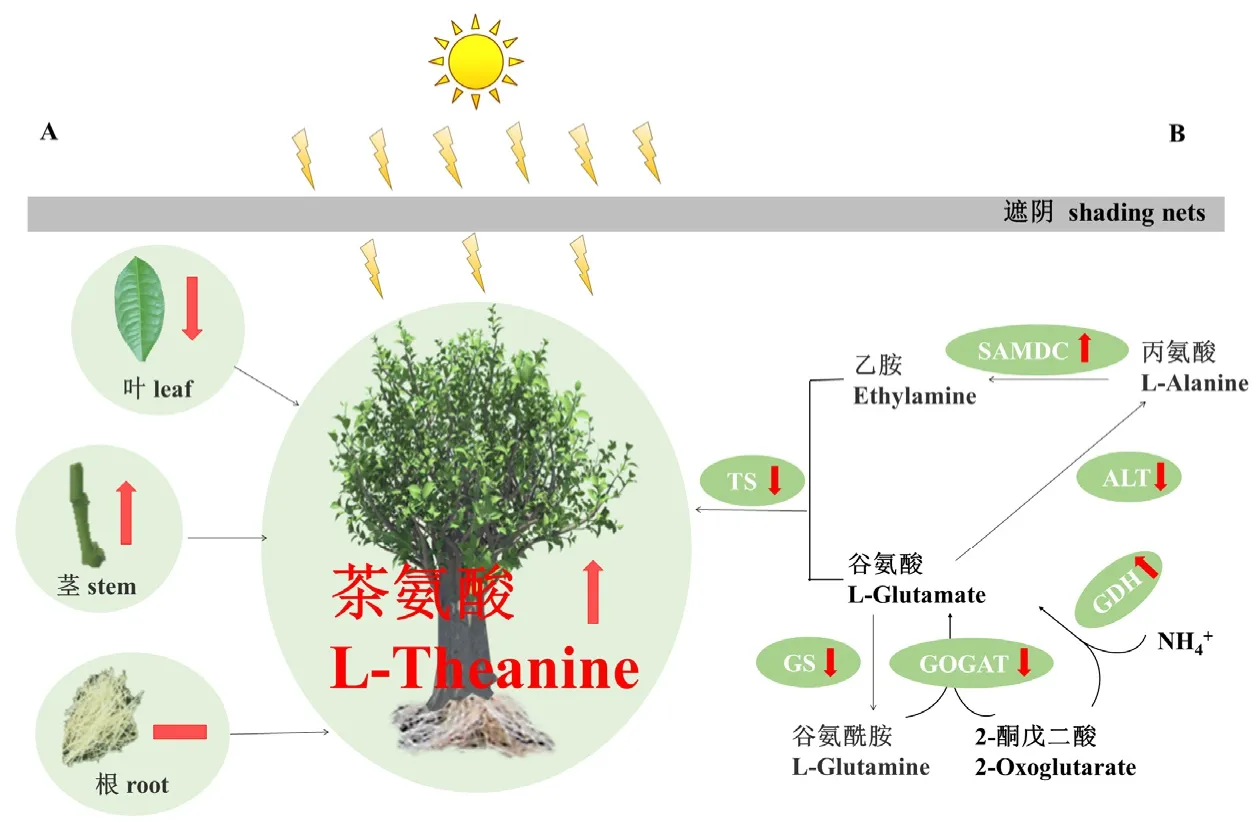
圖2 遮陰對(duì)茶氨酸含量的影響機(jī)制[47,68]Fig. 2 Mechanism of shading on theanine content change[47,68]
在研究遮陰效果的同時(shí),也對(duì)遮陰影響的原理進(jìn)行了探究。對(duì)于茶氨酸的作用效果,Yang等[68]發(fā)現(xiàn),在遮光條件下,茶氨酸含量在莖中增加,在葉中減少,在根中保持穩(wěn)定(圖2-A),原因是遮陰顯著增加了莖中茶氨酸轉(zhuǎn)運(yùn)蛋白基因CsAAP2/4/5/8的表達(dá),同時(shí)降低了葉中CsAAP1/2/4/5/6的表達(dá)。陳琪等[69]針對(duì)葉片部分進(jìn)行了研究,發(fā)現(xiàn)遮陰處理上調(diào)了茶樹(shù)嫩葉中茶氨酸合成酶基因(TS)的表達(dá),與葉片氨同化作用相關(guān)基因均在老葉中顯著上調(diào),而與氮素吸收轉(zhuǎn)化相關(guān)基因在老嫩葉中表達(dá)量均下調(diào)。此外,在茶樹(shù)根中,遮光可顯著增加乙胺并激活茶氨酸生物合成途徑和茶氨酸轉(zhuǎn)運(yùn)蛋白基因,同時(shí)影響茶樹(shù)根系對(duì)外源氮的吸收與利用,進(jìn)而調(diào)節(jié)根中編碼氨基酸生物合成相關(guān)基因(CsGDH、CsAlaDC、CsAspAT)的表達(dá)[70]。Xu等[47]試驗(yàn)發(fā)現(xiàn),參與茶氨酸生物合成的基因(Gs、GOGAT、TS和ALT)在遮光處理下受到抑制,但谷氨酸脫氫酶(GDH)和精氨酸脫羧酶(ADC)的表達(dá)量在遮光處理后增加,負(fù)責(zé)茶氨酸合成前體乙胺生物合成的S-SAMDC在遮光條件下過(guò)度表達(dá),這可能是遮陰導(dǎo)致茶氨酸含量增加的主要原因之一(圖2-B)。此外,Shao等[71]研究發(fā)現(xiàn),遮陰 12 d后的湘妃翠和金萱茶樹(shù)葉片和根系中的氨基酸含量顯著升高,葉片和根系中特定氨基酸合成基因(CsGS1.1和CsTSI)的表達(dá)量增加,這兩個(gè)茶樹(shù)品種氨基酸含量的調(diào)控存在相似性。由此可見(jiàn),遮陰可以促進(jìn)茶樹(shù)茶氨酸的生物合成和分配,影響其含量。
不同光質(zhì)處理也會(huì)對(duì)茶樹(shù)中氨基酸含量產(chǎn)生影響。王加真等[72]綜合了紅藍(lán)兩種單色光的優(yōu)點(diǎn),提出紅光和藍(lán)光為1∶3的復(fù)配光質(zhì)的照射效果較佳,有利于茶樹(shù)葉片中功能成分的積累,降低酚氨比。復(fù)合光中藍(lán)光比例越高,越有利茶氨酸、谷氨酸、天冬氨酸的積累,游離氨基酸含量的增加、酚氨比下降,可能是藍(lán)光能阻止氨基酸向多酚轉(zhuǎn)化的結(jié)果[73]。Zhang等[74]也證實(shí),光量子通量密度為 100 μmol·m-2·s-1,紅光(660 nm)和藍(lán)光(450 nm)比為1∶3時(shí),茶樹(shù)葉片中氨基酸含量高。而陳思肜[75]采用不同強(qiáng)度的紅藍(lán)光對(duì)茶樹(shù)進(jìn)行補(bǔ)光處理,在新鮮嫩葉中共檢測(cè)到28種游離氨基酸,其中茶氨酸、谷氨酸、谷氨酰胺和天冬酰胺含量受紅藍(lán)光補(bǔ)光光強(qiáng)、光照時(shí)間的影響較大,紅光補(bǔ)光的光照強(qiáng)度為 480 μmol·m-2·s-1、光照時(shí)間為 14 d 時(shí),茶氨酸、谷氨酸、谷氨酰胺和天冬酰胺含量均較高,而精氨酸含量與紅藍(lán)光無(wú)明顯相關(guān)性。但是已有研究中涉及光質(zhì)較少,光質(zhì)對(duì)氨基酸含量調(diào)控的分子機(jī)制還有待進(jìn)一步研究。
3.3 芳香類物質(zhì)
茶葉的香氣是由茶樹(shù)中芳香類物質(zhì)組成所決定[76]。Yang等[77]發(fā)現(xiàn),茶樹(shù)遮光處理 3周后,其揮發(fā)性物質(zhì)苯丙素/苯二酚(VPBs)顯著增加。二甲基硫醚(DMS)是綠茶“新茶香”形成的關(guān)鍵香氣物質(zhì),甲基蛋氨酸硫銨(MMS)是 DMS的前體,Sakaida等[78]試驗(yàn)得出,適度遮光可降低MMS含量,進(jìn)而導(dǎo)致DMS的合成受到抑制。
不同光質(zhì)對(duì)茶樹(shù)芳香類物質(zhì)的影響也不同。Hao等[79]利用藍(lán)色、紫色和黃色塑料薄膜處理茶樹(shù),發(fā)現(xiàn)藍(lán)光可下調(diào)苯丙烷類生物合成途徑基因的表達(dá),紫光和黃光可下調(diào)苯丙烷類生物合成和苯丙氨酸代謝途徑基因的表達(dá),影響茶葉香氣。谷夢(mèng)雅等[80]發(fā)現(xiàn),補(bǔ)充 120 μmol·m-2·s-1和240 μmol·m-2·s-1的紅光,茶樹(shù)的差異基因顯著富集在苯丙烷類生物合成、氨基糖和核苷酸糖代謝等代謝通路中。此外,藍(lán)光(470 nm)和紅光(660 nm)可顯著增加采前茶葉中的大多數(shù)內(nèi)源揮發(fā)物,包括揮發(fā)性脂肪酸衍生物(VFADs)、揮發(fā)性苯丙烷/苯類化合物(VPBs)和揮發(fā)性萜烯(VTs),主要因?yàn)樗{(lán)光和紅光顯著上調(diào)了參與VFADs形成的9/13脂氧合酶(LOX)、參與 VPBs形成的苯丙氨酸氨化酶(PAL)和參與VTs形成的萜烯合成酶(TPS)的活性[81]。Xiao等[82]研究發(fā)現(xiàn),受茶小綠葉蟬侵染的茶葉在藍(lán)光(450 nm)和紅光(660 nm)下處理 96 h可誘導(dǎo)芳樟醇氧化物及其糖苷的產(chǎn)生,這說(shuō)明光引起的非生物脅迫與茶小綠葉蟬侵染協(xié)同促進(jìn)了芳樟醇向芳樟醇氧化物及其糖苷的轉(zhuǎn)化。對(duì)于UV-B處理,發(fā)現(xiàn)短期照射能夠促進(jìn)茶葉揮發(fā)性物質(zhì)的釋放,苯甲醇、苯乙醇等芳香醇及其氧化物等含量顯著提高,并且茶樹(shù)葉片中催化糖苷酶轉(zhuǎn)化為香氣成分的關(guān)鍵酶β-櫻草糖苷酶與β-葡萄糖苷酶基因高表達(dá)[83]。Shamala等[58]試驗(yàn)得出,茶樹(shù)在UV-B下照射30 min可誘導(dǎo)完整茶葉部分單萜類氣味物質(zhì)(如芳樟醇和香葉醇)的生成,隨著光照時(shí)間的延長(zhǎng),萜類香氣物質(zhì)的積累受到抑制。適當(dāng)?shù)腢V-B可以刺激β-半乳糖苷酶和β葡萄糖苷酶基因(TBP)的表達(dá),從而導(dǎo)致茶葉醇類芳香揮發(fā)物的釋放[84]。
綜上所述,茶樹(shù)體內(nèi)多酚類、氨基酸以及芳香類物質(zhì)的光調(diào)控具有復(fù)雜性,各項(xiàng)指標(biāo)對(duì)光的響應(yīng)不同(表2),表現(xiàn)出復(fù)雜的光生物學(xué)效應(yīng)[85]。光可直接影響代謝產(chǎn)物合成,也可通過(guò)調(diào)控基因表達(dá)來(lái)影響茶葉中物質(zhì)變化。因此,光可以作為調(diào)控茶葉中主要代謝產(chǎn)物組分以及提高茶葉品質(zhì)的重要手段。

表2 光照對(duì)茶樹(shù)主要代謝產(chǎn)物的影響Table 2 Effects of light on main metabolites of tea plants
4 茶葉產(chǎn)量光調(diào)控
光照調(diào)控不僅影響茶樹(shù)的生長(zhǎng)發(fā)育,同時(shí)也影響茶葉產(chǎn)量。殷劍美等[86]通過(guò)分析大棚茶園小氣候特征,對(duì)比大棚茶園和露天茶園茶葉產(chǎn)量,發(fā)現(xiàn)茶樹(shù)冠面的光照強(qiáng)度與茶青產(chǎn)量呈正相關(guān),適度遮光的明前茶、春茶和夏秋茶產(chǎn)量均大幅度增產(chǎn)。劉青如等[87]對(duì)夏茶進(jìn)行遮陰處理,設(shè)置遮光度50%、70%和90%,得出遮光程度為70%時(shí)夏茶收益最大。張文錦等[88]研究發(fā)現(xiàn),黃旦、本山和鐵觀音3個(gè)茶樹(shù)品種在 30%和 45%遮陰處理下,其產(chǎn)量均高于未遮陰處理。這主要因?yàn)橄那锛局形鐪囟容^高,遮陰可有效減少高溫對(duì)茶芽的灼傷,提高茶葉產(chǎn)量[89]。冬春季覆蓋薄膜可以在改善光強(qiáng)的同時(shí)使大棚茶園的總積溫增加,起到保溫、防霜凍的作用,溫度和光照的互作調(diào)控使得茶產(chǎn)量增加。但單武雄等[90]試驗(yàn)發(fā)現(xiàn),不同遮光處理對(duì)秋季白露毛尖茶產(chǎn)量無(wú)明顯差異,這可能是由于特殊光周期下導(dǎo)致茶樹(shù)葉片受到的光照不足,一定程度影響了茶樹(shù)的光合作用,使茶芽生長(zhǎng)速度受限。
密度和百芽質(zhì)量是衡量茶葉產(chǎn)量的基礎(chǔ),也受到光照的影響。肖文敏等[91]研究發(fā)現(xiàn),覆蓋灰、紅、藍(lán)色遮陽(yáng)網(wǎng)的茶樹(shù)芽頭密度效果較好,且有利于茶葉滋味物質(zhì)的合成。王加真等[92]在貴州茶區(qū)通過(guò)紅藍(lán)LED光調(diào)控試驗(yàn)發(fā)現(xiàn),藍(lán)光占比高的LED燈(紅藍(lán)光質(zhì)比0.81)能夠增加茶樹(shù)芽頭數(shù)量和重量,可滿足對(duì)茶樹(shù)高質(zhì)量和高產(chǎn)量的需求。上述研究表明,以是遮陰手段調(diào)控光照可以影響茶葉產(chǎn)量,但是調(diào)控光質(zhì)對(duì)茶葉產(chǎn)量影響的研究還較少,不同波段、光強(qiáng)參數(shù)下的調(diào)控機(jī)制也不清晰,有待下一步研究。
5 展望
近年來(lái),國(guó)內(nèi)外學(xué)者在光對(duì)茶樹(shù)生長(zhǎng)影響的方面進(jìn)行了廣泛的研究,證明了光調(diào)控是促進(jìn)茶樹(shù)生長(zhǎng),改善茶葉品質(zhì)的有效手段,但有些研究仍有待進(jìn)一步深入探索。
(1)精準(zhǔn)補(bǔ)光。由于不同光對(duì)茶樹(shù)的影響有差異,明確光譜波段、光量子通量密度、光周期的比例和范圍,有助于高效準(zhǔn)確地調(diào)控茶樹(shù)生長(zhǎng)發(fā)育。現(xiàn)有研究成果范圍太過(guò)寬泛,同類型補(bǔ)光不夠全面,遮陰或者使用濾光膜無(wú)法準(zhǔn)確判斷茶樹(shù)對(duì)光的最佳需求,缺少適合的補(bǔ)光燈具,因此,可針對(duì)性開(kāi)發(fā)專用LED燈具。而且茶樹(shù)種類繁多、不同品種茶樹(shù)對(duì)光的感受不同,某一種光對(duì)茶樹(shù)的調(diào)控是否具有普遍性,光調(diào)控不同品種茶樹(shù)是否具有規(guī)律性,均值得深入研究。
(2)分子機(jī)制研究。目前光調(diào)控茶葉品質(zhì)的研究,對(duì)多酚類物質(zhì)的變化分子機(jī)制解釋較多,研究比較深入,但是在氨基酸、芳香類化合物方面研究還不夠深入,機(jī)制解析不清晰,可能會(huì)成為未來(lái)的研究熱點(diǎn)。
綜上所述,光調(diào)控在茶樹(shù)生長(zhǎng)方面還有很多的未知領(lǐng)域,需要加快光對(duì)茶樹(shù)生長(zhǎng)精準(zhǔn)調(diào)控的探索研究,解析變化的分子生物學(xué)機(jī)制,助力茶產(chǎn)業(yè)的發(fā)展。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(xiāo)(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
