茶樹白化果皮超微結構觀察及分子機制研究
2023-01-11 07:42:54湯榕津劉浩然劉丁丁張晨禹龔洋葉圓圓陳杰丹陳亮馬春雷
茶葉科學 2022年6期
湯榕津,劉浩然,劉丁丁,張晨禹,龔洋,葉圓圓,陳杰丹,陳亮,馬春雷*
1. 中國農業科學院茶葉研究所/農業農村部特種經濟動植物生物學與遺傳育種重點實驗室,浙江 杭州 310008;2. 中國農業科學院研究生院,北京 100081
茶樹是我國傳統木本葉用經濟作物,在長期的自然選擇及人工繁育下發生了豐富的表型變異,現有的種質資源從外形到生化成分等方面各具特色。其中,茶樹葉色白化突變類型豐富,其研究范圍涵蓋了生理生化、細胞學、分子生物學等多方面。以前的研究表明,茶樹葉片的白化常常伴隨著葉綠體發育缺陷現象的發生,如葉綠體內部空泡化,缺乏類囊體片層等,但大多數茶樹資源的葉綠體在特定條件下又可恢復正常[1]。以茶樹白化葉片與返綠葉片為材料的研究中,轉錄水平的差異表達基因主要富集于光合作用、葉綠素、活性氧清除等代謝通路[2-4];而代謝組所獲得的差異代謝物則主要參與了碳水化合物代謝、氨基酸代謝、次級代謝物合成等通路[5-6]。但白化突變的形成機理實際上十分復雜,除了與葉綠體的異常發育或葉綠素合成、降解相關途徑的基因突變有關之外,核質信號轉導、轉錄因子的調控以及表觀遺傳的作用等均有可能引起植株的白化表型[7]。葉色白化突變體在自然界中是較為常見的變異類型,但其他組織或器官出現白化的情況仍較少見,僅少量研究提及玉米籽粒[8]、大麥穎殼[9]、甘藍形油菜角果[10]和甘藍種莢[11]的特異性白化。
本課題組在國家種質杭州茶樹圃保存的眾多葉色變異資源中發現了一份葉色呈現出非純色系白化、果皮呈現白色表型的珍貴特異資源——云白1號,其種子播種后仍可正常發育,且一年生幼苗未出現任何白化或其他生長缺陷現象。在茶樹中,有關果皮白化的表型目前未能查閱到相關報道,為進一步探討白化果皮的形成原因,本研究采用轉錄組測序技術對云白1號、中茶129的果皮、種子進行分析,比較兩個茶樹品種的不同組織在轉錄層面的差異,以期為深入研究白化果皮的分子機理提供理論基礎。
1 材料與方法
1.1 試驗材料
于2020年8月上旬在國家種質杭州茶樹圃采集云白1號(YB1)和中茶129(ZC129)的一芽二葉新梢(圖1-A、E)和已成熟的果實(圖1-B、F),將采集到的果實剝開,留取新鮮果皮組織和種子分別保存,部分果皮切片后放于固定液中保存,用于后續超微結構的觀察;其余組織用液氮迅速固定并置于-80℃冰箱保存,用于轉錄組測序。
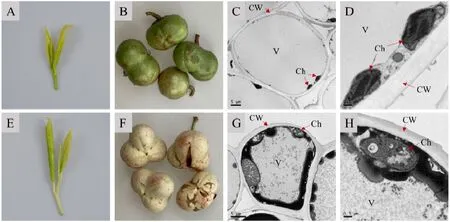
圖1 中茶129、云白1號果皮細胞在不同放大倍數下的超微結構圖Fig. 1 Ultrastructure of pericarp cells of ‘Zhongcha 129’ and ‘Yunbai 1’ at different magnification
1.2 透射電鏡掃描果皮結構
將果皮切成1 mm×1 mm左右的組織薄片后放入電鏡固定液進行前固定,之后組織薄片經漂洗、后固定、室溫脫水、滲透包埋、聚合等步驟制成樹脂塊,使用超薄切片機切片。切片經染色、清洗、干燥等步驟后,放置于透射電子顯微鏡下采集果皮超微結構圖像。
1.3 轉錄組測序及數據分析
1.3.1 文庫的構建、測序及比對
每個樣本設置 3個生物學重復,使用Trizol試劑盒提取總RNA,對純度、濃度等進行檢測后富集帶有 polyA結構的高質量mRNA,采用隨機引物對片段化后的 mRNA進行反轉錄,初步構建 cDNA文庫。此后經PCR富集、片段篩選、質量檢測、混合稀釋、堿變性等步驟建立單鏈文庫。在 Illumina HiSeq平臺對文庫進行雙末端測序,去除原始下機數據中的低質量或含有接頭的 reads,將獲得的高質量reads比對至舒茶早茶樹染色體級別參考基因組[12],得到BAM文件。
1.3.2 差異表達基因的篩選及富集分析
使用HTSeq軟件將BAM文件比對至參考基因組的結構注釋文件,統計比對到每一個基因上Read count值,作為基因的原始表達量,采用FPKM對表達量進行標準化。利用DESeq軟件分析基因的差異表達,以表達差異倍數|log2FoldChange|>1,P<0.05 對差異表達基因進行篩選。以整個基因組為背景,采用BinGO、R語言、TBtools分別繪制GO柱狀圖、KEGG氣泡圖、表達倍數熱圖。
2 結果與分析
2.1 果實的表型及果皮細胞超微結構
在中茶129和云白1號果皮的超微結構掃描電鏡圖中發現(圖1),兩者的葉綠體分布情況以及生長發育狀況不同,中茶129的綠色果皮細胞中僅具有少數葉綠體,其基粒和基粒片層均尚未形成,處于發育初期,分布于細胞壁附近(圖1-C,1-D)。相比而言,云白1號白色果皮細胞中的質體內部高度空泡化,缺乏類囊體和基粒等結構,不存在正在發育或發育成熟的葉綠體(圖1-G,1-H),這與擬南芥pds3白化突變體[13]、中茶108白化芽變枝條[14]葉肉細胞的超微結構類似,可能意味著云白1號果皮細胞中的前質體失去正常分化功能,無法進一步形成葉綠體。因此,云白1號果皮白化表型的產生與其細胞內質體階段的發育缺陷密切相關。
2.2 轉錄組測序質量分析
測序結果表明,18個文庫的 clean reads比率、Q30均大于 92%(表1),說明測序質量良好。果皮和新梢的clean reads比對至參考基因組的百分比較高,均達到87%以上,說明這兩個樣本的測序質量良好;而種子的比對率則處于67.51%~78.01%,這可能是種子組織作為茶樹轉錄組的研究材料具有一定的特殊性,數據庫缺乏充分的參考信息所致。
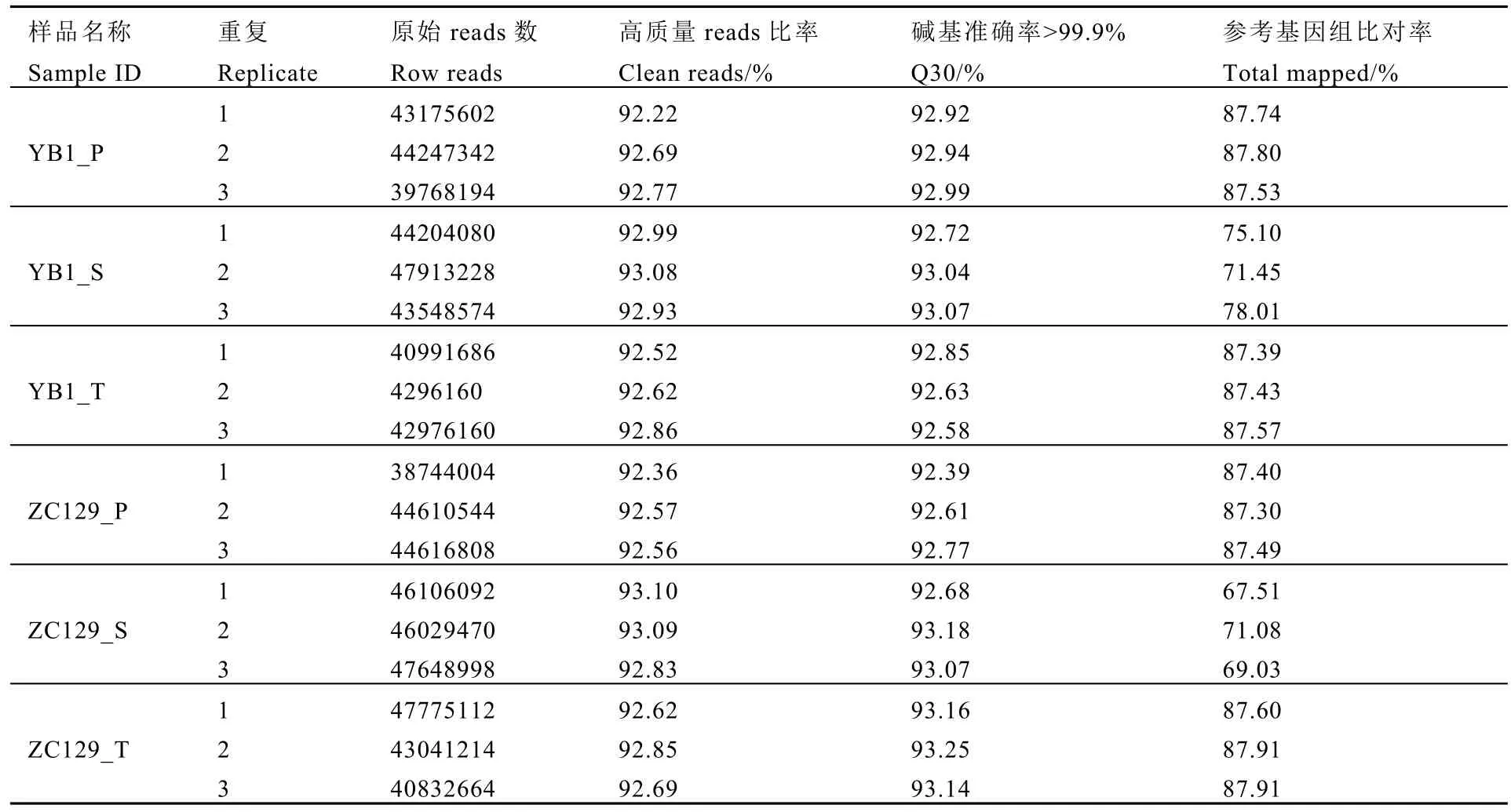
表1 測序質量及參考基因組比對結果統計表Table 1 The statistics of sequencing data and mapping data
2.3 轉錄組樣本的主成分分析
對18個樣本的轉錄本表達量信息進行主成分分析(Principal component analysis,PCA)與聚類分析(圖2),結果顯示,第一主成分(PC1)的貢獻率為43.583%,第二主成分(PC2)的貢獻率為28.212%,各樣品的重復性較好,且按照樣本組織類別兩兩相互聚集,而非按照品種類別區分開來。這表明兩個主成分所代表的各組織之間的轉錄本信息差距大于兩個品種相同組織之間的轉錄本信息差距,反映了茶樹的組織特異性,推測與相同組織中的基因執行相似的生物學功能有關。同時,此聚類結果也意味著不同品種的相同組織之間的轉錄組比較能夠在一定程度上減少與目標表型無關的信息噪音,提高樣本比較的可信度。因此,本研究以不同果皮、不同種子作為差異組合,分析組合間的基因表達差異。

圖2 18個樣本的轉錄組PCA得分Fig. 2 Expression data of 18 transcriptome sample analyzed by PCA
2.4 差異表達基因分析
對果皮和種子的轉錄組數據分別進行差異表達分析并繪制火山圖(圖3),結果顯示,云白1號和中茶129果皮之間的差異基因數目較多,一共篩選出7 537個,其中上調的基因為3 616個,包含云白1號果皮中特異表達的基因508個;下調的基因為3 921個,包含中茶 129果皮中特異表達的基因 639個(圖3-A)。種子之間的差異基因數目較少,僅篩選出2 612個,其中上調的基因為1 233個,包括在云白1號種子中特異表達的基因137個;下調的基因為1 379個,包括在中茶129種子中特異表達的基因 179個(圖3-B)。與中茶129相比,云白1號的果皮以及種子的下調基因數目較多,特異性表達的基因數目較少,這部分差異基因的表達模式可能與白化果皮的形成有所關聯。
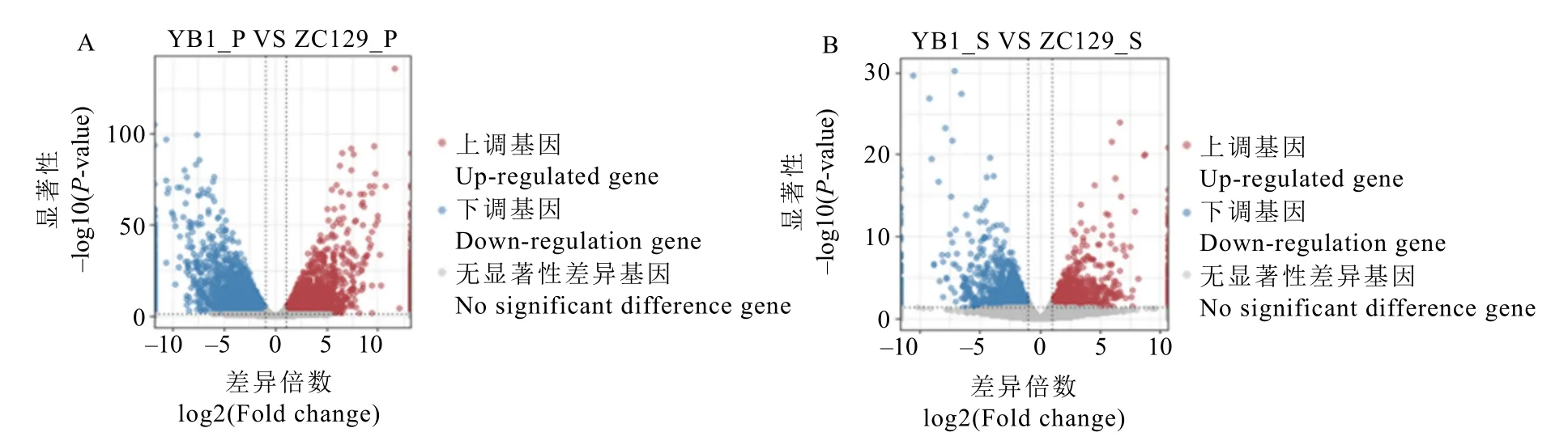
圖3 差異基因火山圖Fig. 3 The volcano plots of DEGs
2.5 GO富集分析
為進一步研究差異基因涉及的生物學功能,將中茶129與云白1號的果皮差異基因與種子差異基因分別進行了GO功能富集分析。結果顯示,果皮差異基因在細胞組分類別中主要富集在與葉綠體、質體相關的功能條目中;在生物學過程類別中差異基因主要參與有機酸代謝過程與小分子代謝過程;在分子功能類別中,差異基因的功能主要與催化活性相關(圖4-A)。其中,質體相關功能條目中共富集了269個上調基因及327個下調基因,這些基因主要參與了光合作用、有機酸代謝、胺生物合成代謝、金屬離子轉運等過程。說明相比于綠色果皮,白化果皮中的光合作用、碳代謝及氮代謝通路相關基因的轉錄水平發生了較大的變化,這與果皮超微結構觀察的結果相符,推測這一結果可能是由于白化果皮中呈現異常狀態的質體未能進一步形成具有完整結構的葉綠體,使得多數在葉綠體被膜或類囊體膜上裝載或在葉綠體內部行使功能的基因無法正常表達而導致的。
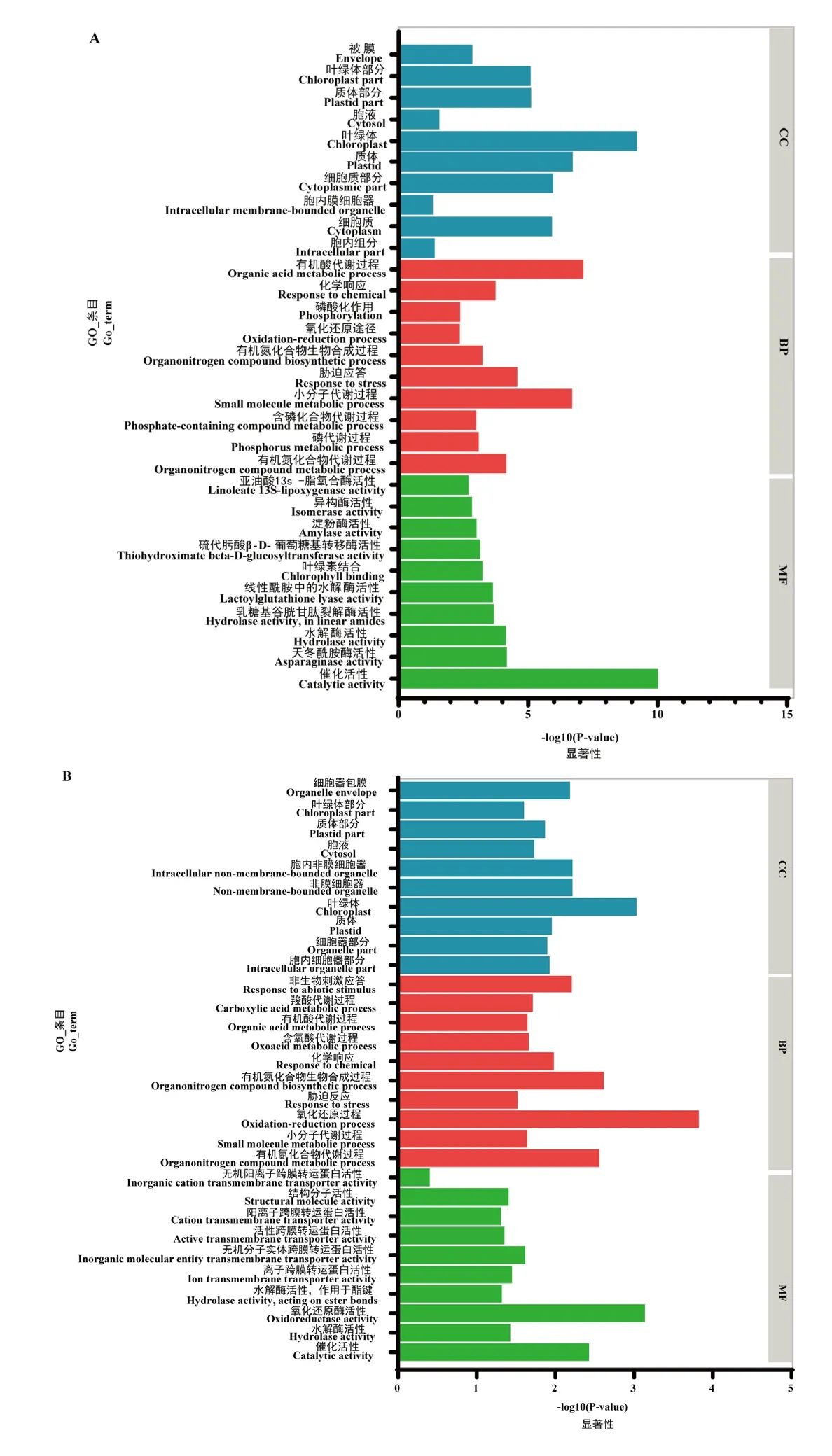
圖4 差異表達基因的GO分析Fig. 4 GO analysis of differentially expressed genes
中茶129與云白1號種子轉錄組GO分析表明(圖4-B),差異基因主要顯著富集在生物學過程和分子功能下的氧化還原過程及氧化還原酶活性條目中,在該條目中植物組織抗氧化相關的基因如編碼谷胱甘肽硫轉移酶合成、谷胱甘肽過氧化物酶合成的基因表達上調十分明顯。此外,屬于分子功能的條目大多與離子轉運有關,表明云白1號的離子運輸方式發生了改變。在所有離子轉運途徑中,與鎘離子相關的通路所富集的基因數目最多,其中包含與植物應激反應相關的調控基因如親環素編碼基因,受植物激素及鎘離子誘導的轉錄因子MYB59等,這些基因的表達量均顯著上升。而為植物生長發育提供必要硫化物的硫酸鹽同化途徑關鍵酶 ATP-硫化酶的編碼基因表達量則明顯下降,這些變化趨勢表明在云白 1號種子內與脅迫及生長發育相關的代謝過程與中茶129種子存在較大差異。
2.6 KEGG富集分析
KEGG的富集分析顯示(圖5-A),相比于中茶129,在云白1號果皮中上調的差異基因主要富集于氨基酸代謝,碳水化合物代謝,半乳糖代謝,谷胱甘肽代謝,半胱氨酸和甲硫氨酸代謝,苯丙氨酸、酪氨酸和色氨酸的生物合成等代謝通路中;下調的差異基因顯著富集于光合器官的碳固定,光合作用蛋白,甘氨酸、絲氨酸和蘇氨酸代謝,光合作用,半胱氨酸和甲硫氨酸代謝等通路中。這些通路中富集的差異基因主要在質體/葉綠體中發揮功能,不僅參與了光合作用有關的通路,還參與了在質體/葉綠體中發生的氨基酸代謝、碳代謝等初級代謝過程,這些基因的差異表達極可能與云白1號質體發育的停滯相關。
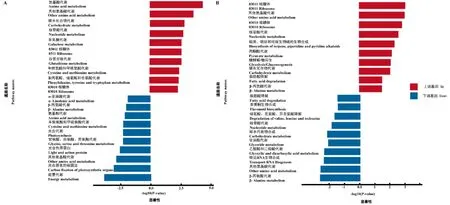
圖5 差異表達基因的KEGG富集分析Fig. 5 The KEGG pathway analysis of DEGs
種子差異基因的KEGG富集結果顯示(圖5-B),云白1號種子中上調的差異基因富集于核糖體,萜類、哌啶和吡啶生物堿的生物合成,丙酮酸代謝,糖酵解/糖異生等通路;下調的差異基因富集于β-丙氨酸代謝,轉運RNA生物合成,乙醛酸和二羧酸代謝,甘油酯代謝等通路。茶樹種子含油率較高,其生命歷程與油料種子相似,常以三酰甘油作為高效儲能形式為萌發提供必要的碳骨架和能量[15]。一旦種子萌發的信號觸發,三酰甘油將經脂肪酸降解途徑轉變為蔗糖,為組織器官的生長分化提供碳源及能量。因此,差異基因所富集的通路可能反映了云白1號與中茶129種子在生長發育進程上的不一致。
2.7 差異基因在KEGG通路中的表達量分析
2.7.1 果皮中與光合作用相關的差異基因
在光反應中(圖6),光合天線蛋白、光系統Ⅰ、光系統Ⅱ、光合電子傳遞鏈以及ATP合成酶的相關調控基因大多在云白 1號果皮內下調表達,這些基因編碼的蛋白包括了捕光色素葉綠素a/b結合蛋白(Light harvesting chlorophyll a/b binding protein,LHC)、鐵氧還蛋白-NADP+氧化還原酶(Ferredoxin-NADP+oxidoreductase,FNR)以及 ATP合成酶亞基蛋白等。光合天線蛋白是光反應中不可或缺的部件,LHC表達量的下調將阻礙光能在類囊體中的吸收和傳遞,而FNR和ATP合成酶所產生的同化力可直接用于暗反應的卡爾文循環,兩者調控基因的下調則意味著碳固定過程產生了一定的變化。
而在卡爾文循環中(圖6),上調的基因主要編碼1,5-二磷酸核酮糖羧化酶(Ribulose-1,5-bisphosphate carboxylase,RCA)、磷酸甘油酸激酶(Phosphoglycerate kinase,PGK),這些酶均處于卡爾文循環的初始反應階段;下調的基因則主要參與編碼卡爾文循環的中間酶,例如3-磷酸甘油醛脫氫酶A/B(Glyceraldehyde-3-phosphate dehydrogenase A/B,GAPA/B)、5-磷酸核糖異構酶、果糖二磷酸醛縮酶、磷酸丙糖異構酶以及轉酮酶等;在非綠色質體中調控3-磷酸甘油醛脫氫酶合成相關的基因則存在兩種相反的表達模式。RCA、PGK是卡爾文循環的關鍵酶,在多數情況下受非生物脅迫的刺激而提升表達量用以維持植物的碳同化速率,云白1號果皮中這兩個酶有關的基因表達倍數的提升可能說明了白化果皮正處于抵抗脅迫的狀態中。
2.7.2 種子中的β-氧化途徑相關的差異基因
云白1號和中茶129種子的差異基因在脂肪酸降解途徑中顯著富集,β-氧化是脂肪酸降解的主要途徑,其產物乙酰輔酶 A可為糖異生和三羧酸循環提供底物,因此脂肪降解途徑對種子的生長發育意義重大。脂肪酸在進入降解途徑前,需要經過酰基輔酶 A合成酶的活化后進入β-氧化途徑(圖7),而呈現下調表達模式的長鏈酰基輔酶 A合成酶極可能刺激β-氧化循環中一系列基因表達量的調整變化,導致長鏈脂肪酸降解過程發生異常。此外,β-氧化循環的限速酶酰基輔酶 A氧化酶、參與反應第二三步驟的多功能蛋白以及直接催化乙酰輔酶A生成的3-酮脂酰-CoA硫解酶等基因表達量均存在一定變化,這可能意味著云白1號種子中的油脂組成發生了改變。
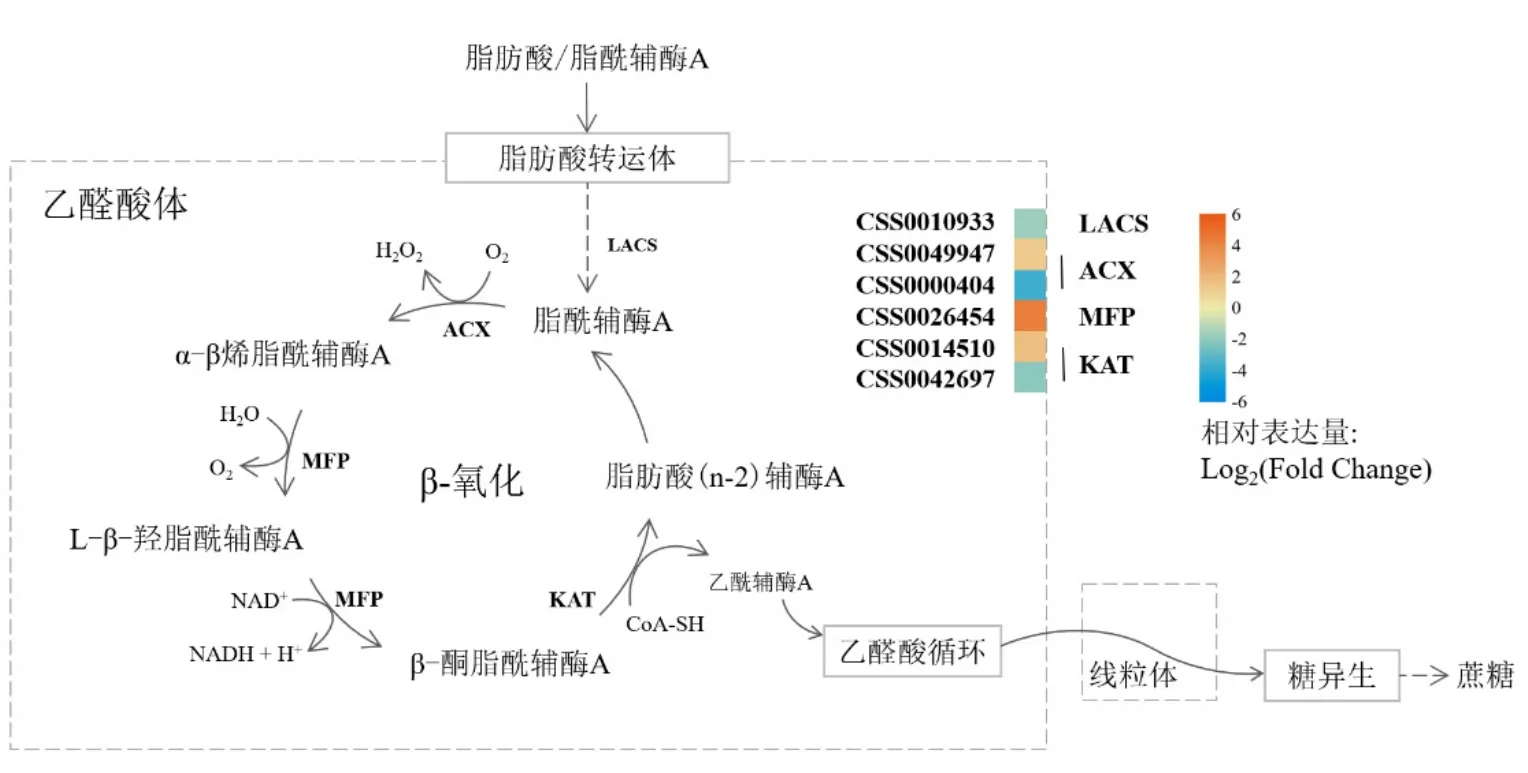
圖7 種子差異基因在脂肪酸降解途徑中的表達情況Fig. 7 The expressions of DEGs of pericarps in the pathway of fatty acid degradation
3 討論
白化突變體的產生大多與葉綠體的非正常發育有關,如擬南芥白化突變體的超微結構顯示,葉綠體缺乏基質和垛疊的類囊體片層,且其內部高度空泡化,聚集了大量球狀結構[16];玉米光合突變體的葉綠體則存在類囊體發育不全、片層結構松散的情況[17];茶樹白化葉片的葉綠體有體積膨大、類囊體片層缺乏或松散、內部空泡化或具有不同程度的降解等現象[18-19]。本研究通過透射電鏡觀察果皮細胞的超微結構,發現茶樹綠色果皮細胞中僅有少數葉綠體附著于細胞壁,其內部結構雖然還未發育完全,但是處于正常的生長階段;而白化果皮細胞不存在處于生長發育階段或已發育成熟的葉綠體,其內部僅觀測到高度空泡化的質體,這與葉色完全白化突變體的研究結果相似[13-14],表明云白 1號果皮細胞中的質體在發育的某一個階段產生停滯、降解,失去形成葉綠體的功能,因而產生特殊的白化表型。
葉綠體是光合作用的主要場所,光反應和暗反應能為碳代謝和氮代謝提供基本能量和有機物質,碳代謝所生成的中間產物酮酸同時也為葉綠體中谷氨酸轉化成其他氨基酸提供了碳骨架。在茶樹葉色白化突變體中,異常發育或嚴重降解的葉綠體導致光合作用無法正常運轉,而光合碳固定的削弱(碳饑餓)會促使碳-氮代謝的重新平衡,因此相較于正常的綠色品種,白化葉片中的有機酸和氨基酸會存在一定程度的積累[20-21],這也在果皮差異基因的 GO富集和 KEGG富集結果中得以體現。在白化果皮中,與PSⅡ的光損傷蛋白質修復、類囊體形成相關的基因FTSH1、FTSH2的表達量上調,參與光反應的基因LHC、FNR、PSA、PSB的表達量下調,LHC負責吸收、轉移光能至光反應中心,是光系統復合物的重要組成部分,其調控基因的下調意味著光能吸收傳遞過程受阻,長期可能會造成葉綠體內部活性氧的積累,干擾葉綠體的正常發育[22]。在光反應中,FNR是催化線性電子傳遞的最后一步,其主要功能是轉移電子將 NADP+還原為NADPH,為CO2固定提供還原力。因此,FNR的下調會直接影響光合作用,從而導致光合碳固定效率的降低[23-24]。RCA、PGK、GAPA/B是卡爾文循環的關鍵酶,其中RCA同時參與了暗反應和光呼吸兩個代謝路徑,能夠根據CO2/O2的濃度比值調整羧化、還原反應,對凈光合速率有著重要的影響[25];PGK、GAPA/B分別消耗來自光反應的ATP和NAPDH,催化3-磷酸甘油酸生成凈光合產物3-磷酸甘油醛,3-磷酸甘油醛可經過一系列的后續反應轉化為淀粉、蔗糖或脂肪,為植物的生長發育提供基本物質[26]。在云白1號果皮中參與卡爾文循環的大多數基因在轉錄水平上發生了變化,而RCA、PGK上調、GAPA/B下調則可能是白化果皮抵抗脅迫,維持光合碳同化效率的策略。
種子處于植物生命生長發育的一個初始階段,涉及能量的儲存轉化或物質的協調分配,因此,中茶129和云白1號種子的差異基因在碳代謝、脂質代謝、核苷酸代謝等初級代謝通路中有著更為明顯的富集。脂質是茶樹種子儲藏能量的高效形式,種子發芽前,源器官如葉片、果皮的光合產物蔗糖被轉運至種子細胞中,經糖酵解等過程轉化為乙酰輔酶A,為脂肪酸的合成提供前體物質;發芽時,種子中儲存的脂質則將經β-氧化、乙醛酸循環、糖異生等途徑轉變成蔗糖的形式,為即將生長分化的其他組織器官提供碳源。在蓖麻和油茶種子中,蔗糖與油脂呈現極顯著負相關,且兩者的動態平衡關系在一定程度上反映著種子所處的生長發育階段[27-28]。本研究中,云白1號和中茶 129種子的脂肪酸合成代謝通路中的基因在表達量上并無差異,但云白1號的種子中β-氧化、乙醛酸/二羧酸循環、糖酵解/糖異生代謝通路相關基因的表達量發生顯著變化,其中乙醛酸循環的差異基因以下調為主,糖酵解/糖異生的差異基因則以上調為主,表明云白1號種子中的脂肪酸降解過程不如中茶 129旺盛。因此,在云白1號中,用于供應種子生長發育的能量要低于中茶129,但如果云白1號用于合成脂肪酸前體所需的蔗糖得到了充足供應,則能在一定程度上滿足其種子后續生長發育的需求。
綠色果皮細胞內含有葉綠體,同樣也能進行光合作用,屬于非葉光合器官。在甘藍型油菜和油菜中,其綠色角果同樣可作為源器官向庫器官(主要為種子)供應光合同化物,角果的不良發育會對種子的含油量、粒重等生理指標產生較大影響[10,29]。然而在大麥中,其穎殼的白化并不會導致大麥的結實率降低和抗病性減弱,但會在一定程度上影響籽粒干物質的積累[9];此外,甘藍種莢白化突變體可產生少量種子,屬于非致死突變[11]。實際上,葉片是光合作用的主要器官,可作為源器官直接向庫器官比如種子提供光合同化物,用于滿足種子的基本生長發育需求,因此果皮的白化并不意味著種子喪失了發芽的能力[30]。蔗糖作為光合作用的主要產物,其分解代謝在碳資源、能量的協調分配中發揮重要作用,其中,蔗糖合酶(Sucrose synthase,SUS)、轉化酶(Invertase,INV)是蔗糖分解代謝的關鍵酶[31]。SUS可調控光合同化物從源器官向庫器官的轉運,在種子發育、響應逆境脅迫等過程中發揮著不可忽視的作用[32];INV催化蔗糖水解成葡萄糖和果糖,可維持細胞滲透壓,并調節蔗糖在植物體中的運輸裝載[33],其活性的缺失會降低光合作用的水平,減少庫器官的庫強[34]。在本研究中,INV的調控基因在云白 1號的種子中特異表達、SUS相關編碼基因的表達顯著上調,說明蔗糖分解代謝在云白1號種子中的活性較高,可能是為了滿足種子的營養物質儲存需求,維持其正常生長發育進程。因此,盡管茶樹白化果皮在轉錄水平上的異常變化可能會對種子不利,導致種子的生長發育機制出現調控差異造成生理指標等方面的變化,但并不一定會影響種子的發芽能力。為了深入了解白化果皮的遺傳特性,本課題組對云白1號的種子進行了播種繁育,目前已獲得健康的一年生幼苗,說明云白1號種子具有正常的生長活力。此外,表型觀察發現,所有一年生幼苗均未出現任何白化變異,初步推測云白1號的白化現象可能是由隱性基因控制。由于茶樹的自交不親和性,云白1號接觸外界綠色茶樹植株傳遞而來的花粉時,其控制白化果皮的基因在子代中大概率為雜合基因,因而云白1號子代果皮表型呈現出綠色的表型,但相關推測需通過進一步的表型觀察與雜交試驗進行驗證。
