菊芋酶解制備低聚果糖糖漿的工藝優化及功能評價
2023-01-13 13:14:56王彩月田康明
食品工業科技 2023年2期
王彩月,田康明
(天津科技大學化工與材料學院,天津 300457)
菊芋(Helianthus tuberosus L., Jerusalem artichoke),又名洋姜、鬼子姜,是一種多年宿根性草本植物,具有耐寒、耐旱、耐鹽堿等特點,生態適應性強[1]。一次種植可連續多年生長,種植成本低,塊莖產量4~5噸/畝[2]。聯合國糧農組織2020年統計數據顯示,中國菊芋種植面積約1.2萬公頃,占全世界菊芋總種植面積的10.5%。菊芋塊莖含有豐富的果糖多聚物,其中菊糖達干物質的65%~80%[3],菊芋來源的菊糖是由呋喃型D-果糖經β (2→1)糖苷鍵連接而成,終端通過α(1→2)糖苷鍵連接一個葡萄糖,聚合度為2~60的天然果聚糖混合物[4-5],其經菊糖內切酶水解其內部的β-2,1-果糖苷鍵生成低聚果糖(Fructooligosaccharides,FOS),是生產低聚果糖的主要原料之一[6-8]。以菊芋來源的菊糖為原料生產的低聚果糖其聚合度通常為2~10,高于蔗糖為原料經轉苷生產的低聚果糖的聚合度[9-11]。因此,菊芋來源的低聚果糖在調節血糖血脂平衡、維持胃腸道功能、促進鈣吸收、降低肥胖和降低患Ⅱ-型糖尿病風險等方面具有更顯著的作用[12-16]。然而,受限于生產成本和市場需求量,菊芋來源的低聚果糖加工量遠低于蔗糖來源的低聚果糖加工量,現階段需借助合理的新途徑解決菊芋豐產豐收后下游加工的問題。此外,目前大部分研究基于菊芋烘干制粉提取菊糖,再經除蛋白質、果膠等雜質后酶解制備低聚果糖[17],該工藝制備菊芋粉的過程復雜、副產物較多,如何以新鮮菊芋為原料,建立高效酶解工藝,制備高營養價值的低聚果糖糖漿產品,降低生產成本、減少環境污染是目前亟待解決的問題。
為此,本文以鮮菊芋為原料,結合菊芋組成成分特點,系統研究適用于菊芋的酶解工藝,并研究來源于黑曲霉的菊糖內切酶、葡聚糖內切酶、木聚糖酶、聚半乳糖醛酸酶和單寧酶在加工制備低聚果糖糖漿中的作用規律和酶解效果,建立并優化復合酶法制備低聚果糖糖漿的新工藝,以期豐富菊芋綜合加工產品的類型,推動菊芋加工產業的發展。
1 材料與方法
1.1 材料與儀器
菊芋 選擇完整、無病斑、蟲蛀的新鮮菊芋塊莖,購自中國河北省邯鄲市果蔬食品公司;低聚果糖標準品 純度99%,江蘇銳陽生物科技有限公司;乙腈 色譜純,國藥集團化學試劑有限公司;DPPH(1,1-二苯基-2-三硝基苯肼)自由基清除能力試劑盒
北京市索萊寶科技有限公司;羥基自由基清除和總抗氧化能力檢測試劑盒 上海市生工生物工程股份有限公司;菊糖內切酶(1480 U/g)、單寧酶(147 U/g)、葡聚糖內切酶(1260 U/g)、木聚糖酶(3720 U/g)、多聚半乳糖醛酸酶(10800 U/g) 本實驗室前期構建并保藏菌株,發酵制備酶液[18];MRS培養基的成分為:1%蛋白胨,1%牛肉膏,0.5%酵母粉,0.2%檸檬酸氫二銨,0.5%無水乙酸鈉,0.2%磷酸氫二鉀,0.005%一水合硫酸錳,0.02%七水合硫酸鎂,0.1%吐溫80;植物乳桿菌(Lactobacillus plantarum)YJG、嗜熱鏈球菌(Streptococcus thermophilus)IFFI 6038、副干酪乳桿菌(Lactobacillus paracei)2001-12 三種菌均由天津科技大學生物催化與生物轉化實驗室保藏;其他試劑 均為國產分析純。
Agilent 1200高效液相色譜儀Shodex RI-201示差檢測器 日本Shodex公司;Prevail Carbohydrate ES 5u糖色譜柱 美國奧泰公司;ZWY-2102C恒溫培養振蕩器 上海智城分析儀器制造有限公司;BAO-80A精密鼓風干燥箱 施都凱儀器設備(上海)有限公司;SP-2012UV型紫外可見分光光度計
上海光譜儀器有限公司;AB204-S型電子分析天平、PHS-2C型pH計 梅特勒-托利多儀器上海公司;1-15臺式高速離心機 德國Sigma公司。
1.2 實驗方法
1.2.1 原料預處理 菊芋塊莖用自來水清洗除去泥土和其他雜質,稱重、切塊,按料水比為1:2(w/w)打漿。
1.2.2 菊芋漿的組分測定 菊糖含量=總糖含量-還原糖含量(苯酚-硫酸法測總糖含量,3,5-二硝基水楊酸法測還原糖含量)[19-21]。以沒食子酸為標準品,用福林酚試劑比色法測定單寧含量[22](具體操作如下:取0.2 mL濃度分別為0.2、0.4、0.6、0.8、1.0 mg/mL的沒食子酸于試管中,加入0.2 mol/L的福林酚試劑0.5 mL,15%的Na2CO3溶液1.0 mL,用蒸餾水補足4 mL。避光反應20 min后,以蒸餾水代替樣液進行調零,在765 nm處測其吸光值)。參考美國國家可再生能源實驗室(NREL)方法,測定纖維素、半纖維素含量[23]。分光光度法測果膠含量[24]。
1.2.3 菊芋酶解用酶的選擇
1.2.3.1 內源性酶對菊芋的酶解效果 為評價內源性酶對菊糖水解和低聚果糖形成的影響,取15 g菊芋漿在50 ℃下水解24 h,每2 h取樣,12000 r/min離心10 min,高效液相色譜法(High performance liquid chromatography,HPLC)測定上清液中葡萄糖、果糖、蔗糖、低聚果糖的含量。
1.2.3.2 內源酶在添加菊糖內切酶條件下對菊糖水解的影響 取30 g菊芋漿用醋酸調pH為5.0,均分為兩份,一份121 ℃高溫滅菌10 min,冷卻至50 ℃,添加15.0 U/g菊糖內切酶(根據菊芋中組分計算酶的理論添加量后加入過量酶);另一份不滅菌,直接加入等量菊糖內切酶,50 ℃酶解12 h,每隔2 h取一次樣品,HPLC測定上清液中葡萄糖、果糖、蔗糖、低聚果糖的含量。
1.2.3.3 菊芋中其他組分對菊糖內切酶酶解效果的影響 為研究菊芋中單寧、果膠、纖維素、半纖維素的存在對菊糖內切酶發揮作用的影響,因此,采用酶解法先去除該組分,再加入菊糖內切酶,通過低聚果糖含量變化來確定菊芋中各組分對菊糖內切酶發揮作用的影響。具體操作如下:取60 g菊芋漿調pH為5.0,分為4份,分別加入0.2 U/g單寧酶、0.25 U/g聚半乳糖醛酸酶、0.15 U/g葡聚糖內切酶、0.15 U/g木聚糖酶(根據菊芋中組分計算酶的理論添加量后加入過量酶),50 ℃預酶解4 h,每份再加入15 U/g菊糖內切酶,50 ℃下酶解12 h,每隔2 h取一次樣品,HPLC測定上清液中葡萄糖、果糖、蔗糖、低聚果糖的含量,以低聚果糖含量變化來確定菊芋中各組分對應水解酶的添加對菊糖內切酶發揮作用的影響。
1.2.3.4 酶解工藝的建立 根據1.2.3.3的結果確定影響菊糖內切酶發揮作用的物質后,進一步研究各水解酶的最優添加量(根據該組分含量計算水解該酶的理論添加量,在理論添加量的基礎上進行梯度設置),具體操作如下:在菊芋漿中加入不同添加量的水解某物質的酶與15 U/g菊糖內切酶,50 ℃下酶解12 h,用HPLC測定上清液中葡萄糖、果糖、蔗糖、FOS含量,以低聚果糖含量變化來確定該酶的最優添加量。之后,將該酶和菊糖內切酶一同加入菊芋漿中,再加入不同添加量的另一種酶,研究另一種酶的最優添加量,以此類推,逐一研究各水解酶的最優添加量。具體量的設置為:單寧酶、木聚糖酶、葡聚糖內切酶添加量為0.02、0.04、0.06、0.08、0.10,聚半乳糖醛酸酶的添加量為0.01、0.03、0.05、0.07、0.09 U/g。
確定各水解酶的最優添加量后,再研究菊糖內切酶最適添加量和最適作用時間,具體操作如下:加入最優添加量的各水解酶和不同添加量的菊糖內切酶(3.0、6.0、9.0、12.0、15.0 U/g),50 ℃、pH5.0下復合酶解12 h,酶解樣品用HPLC測定上清液中低聚果糖的含量,確定菊糖內切酶的最優添加量;加入最優添加量的各水解酶和最優添加量的菊糖內切酶,每隔2 h取一次樣品,酶解樣品用HPLC測定上清液中低聚果糖的含量,確定菊糖內切酶的最適作用時間。
1.2.4 HPLC外標法測定低聚果糖含量
1.2.4.1 樣品前處理 取酶解樣品于1.5 mL離心管中,12000 r/min離心10 min取上清液,按1:1(v/v)加入無水乙醇,4 ℃下靜置2 h,12000 r/min離心10 min,取上清液,用0.22 μm有機微孔膜過濾,-20 ℃保存,待測。
1.2.4.2 液相色譜條件 高效液相色譜儀Shodex RI-201示差檢測器;色譜柱:Prevail Carbohydrate ES 5 μm(250 mm×4.6 mm);柱溫:35 ℃;進樣量:20 μL;流速:1.0 mL/min;流動相:水/乙腈=35/65(v/v)。
1.2.4.3 水解率計算 以低聚果糖為標準品,對各峰進行定量測定,采用峰面積歸一化法確定低聚果糖中每種物質的相對含量。用以下公式計算水解率:
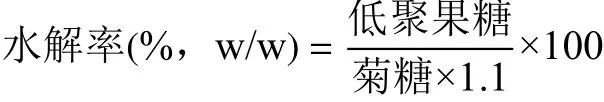
式中:低聚果糖為果二糖、果三糖、蔗果三糖、果四糖、蔗果四糖、果五糖、蔗果五糖、果六糖、蔗果六糖含量的總和[9];1.1為菊糖和低聚果糖的換算系數。
1.2.5 低聚果糖糖漿的制備及其益生功能、抗氧化活性測定
1.2.5.1 低聚果糖糖漿的制備 菊芋在上述最優工藝條件下得到的菊芋水解液,經真空濃縮兩倍體積后制成低聚果糖糖漿。為驗證低聚果糖糖漿的益生元功能,以植物乳桿菌、嗜熱鏈球菌、副干酪乳桿菌為評價菌,研究這三種菌以低聚果糖糖漿為碳源的生長情況(對照為未經水解的菊芋漿原液),評價低聚果糖糖漿的益生元功能。
抗氧化活性測定主要包括DPPH自由基清除能力(DRSA)、羥基自由基清除能力(HRSA)和總抗氧化能力(T-AOC),均按照文獻方法進行[25-27]。對照試驗為菊芋原液。
1.2.5.2 益生功能評價方法 將植物乳桿菌、嗜熱鏈球菌、副干酪乳桿菌分別在以上述制備的低聚果糖糖漿為碳源的MRS培養基和以未經水解的菊芋漿原液為碳源的MRS培養基中,37 ℃培養12 h,每2 h取樣一次,分光光度計測OD600以研究三種菌的生長情況[28]。
1.2.5.3 DPPH自由基清除能力測定 用甲醇溶液制備濃度為0.1 mmol/L的DPPH試劑,取0.1 mL樣品與0.9 mL DPPH自由基溶液混合,將溶液充分混合,并在室溫下暗置30 min。以抗壞血酸為標準品,測定在517 nm處吸光度下降。DPPH自由基清除率計算公式如下:

其中,A0為對照的吸光度,Ai為樣品的吸光度。
1.2.5.4 羥基自由基清除能力測定 羥基自由基清除能力(HRSA)測定的具體方法為:取0.2 mL(10 mmol/L)的FeSO4-EDTA混合液于具塞試管中,加入0.5 mL(10 mmol/L)的脫氧核糖溶液,然后加入0.2 mL樣品溶液,用pH=7.4、0.1 mol/L磷酸緩沖液定容至1.8 mL;再加入0.2 mL(10 mmol/L)H2O2,混勻后置于37 ℃恒溫水浴中反應1 h,然后加入2.8%三氯乙酸溶液1 mL,1%硫代巴比妥酸(TBA)溶液1 mL,混勻后置于沸水浴中反應10 min,冷卻后于520 nm處比色測定吸光值As。不加樣品溶液同上操作處理,所測吸光值Ac。不加樣品且不在37 ℃水浴中反應,其他處理同上,測定空白吸光值A0。羥自由基清除能力HRSA計算公式如下:

1.2.5.5 總抗氧化活性測定 總抗氧化活性(T-AOC)測定的具體方法為:取1 mL樣品與2.5 mL磷酸鹽緩沖液(0.2 mol/L,pH 6.6)和2.5 mL 10 g/L K3Fe(CN)6混合,在50 ℃下反應30 min。之后,加入2.5 mL 100 g/L的三氯乙酸溶液終止反應,混合物1800 r/min離心10 min。最后取上清液2.5 mL,與2.5 mL超純水和0.5 mL 1 g /L FeCl3混合。以抗壞血酸為標準品,在700 nm處測定吸光度。
1.3 數據處理
采用EasyChrom進行HPLC數據分析,采用Excel 2016進行數據統計,采用Origin 2018進行繪圖分析,實驗數據以平均值±標準差(Mean±SD)表示。
2 結果與分析
2.1 菊芋漿基本組成成分
分析了菊芋漿的基本化學成分,結果見表1,主要包括菊糖、單寧和纖維素等,本研究所測得的成分為菊芋打漿后的組分含量,換算后在已有研究菊芋組分的范圍內[29]。

表1 菊芋漿中主要成分Table 1 The main components of Jerusalem artichoke pulp
2.2 菊芋酶解用酶的選擇
2.2.1 內源酶對菊芋水解物低聚果糖含量的影響由表2可知,低聚果糖的含量在24 h內由4.17 g/L增加到7.38 g/L(P<0.05),表明菊糖逐漸被水解,菊芋中存在內源性菊糖水解酶;果糖、葡萄糖和蔗糖含量逐漸增加,說明菊芋中存在水解大分子的內源酶;低聚果糖含量在逐漸增加,說明菊芋中存在的內源酶不會將低聚果糖水解。此外,內源酶水解24 h,水解率仍低于15%,說明需要添加外源性菊糖內切酶,加速菊糖的水解。
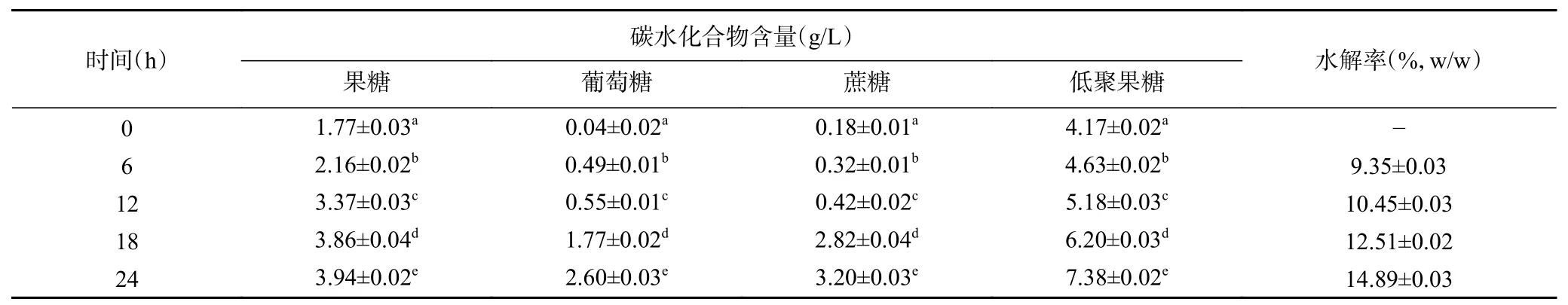
表2 菊芋中內源性菊糖水解酶對低聚果糖得率的影響Table 2 Effect of endogenous inulin hydrolase from Jerusalem artichoke on FOS yield
2.2.2 內源酶對外加菊糖內切酶水解菊糖的影響為探究菊芋中內源酶存在對菊糖內切酶發揮作用是否存在影響,研究了在保留內源酶和內源酶滅活的樣本中加入菊糖內切酶后,將菊糖水解為低聚果糖的情況,結果如圖1所示。由圖可知,當酶促反應由內源酶和菊糖內切酶共同作用時,10 h后低聚果糖的含量最大(17.53 g/L),而單獨使用菊糖內切酶時,10 h后低聚果糖含量基本保持不變(15.60 g/L)(圖1A);內源酶和菊糖內切酶共同作用時,低聚果糖含量比單獨使用菊糖內切酶作用高3.09 g/L,且菊糖水解率比單獨使用菊糖內切酶高12.39%(P<0.05)(圖1B)。說明菊芋中含有一定量的菊糖酶,且內源酶和外加的菊糖內切酶有協同作用,因此,在菊糖內切酶水解菊糖之前,不需要對內源酶進行滅活。

圖1 內源酶在添加菊糖內切酶條件下對低聚果糖含量(A)和水解率(B)的影響Fig.1 Effect of endogenous enzymes on FOS content (A) and FOS yield (B) by the condition of adding endoinulinase
2.2.3 菊芋中影響菊糖內切酶發揮作用的物質 為研究單寧、果膠、纖維素、半纖維素對菊糖內切酶發揮作用的影響,通過加入各組分對應的水解酶,再加入菊糖內切酶,比較產物中低聚果糖含量評價各組分對菊糖內切酶酶解作用的影響,結果如圖2所示。
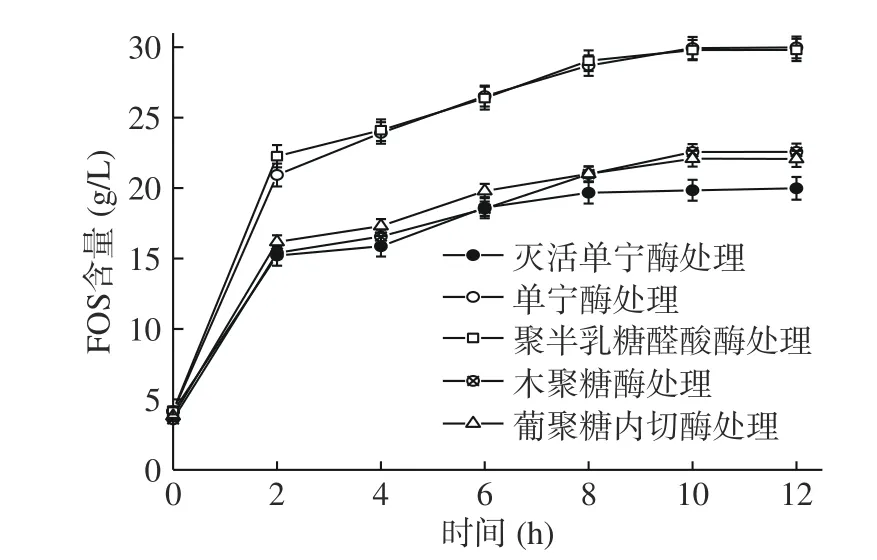
圖2 不同酶預處理對菊糖內切酶水解產生低聚果糖的影響Fig.2 Effects of different enzymes pretreatment on FOS production by endoinulinase hydrolysis
單寧酶預處理使低聚果糖含量由19.98 g/L增加到29.98 g/L(P<0.05),提高50.05%。單寧含有酚羥基、羥基、羧基等多種活性官能團,它可與蛋白質進行共價交聯,從而影響酶的活性[30],因此,菊芋中單寧的存在不利于菊糖內切酶發揮其水解作用,需在加入菊糖內切酶之前預先通過酶解去除單寧。
聚半乳糖醛酸酶預處理使低聚果糖含量達到29.80 g/L(P<0.05),提高49.17%。可能由于果膠的分解使菊芋中菊糖更好地釋放,提高了菊糖內切酶的作用效率;研究表明,果膠的水解產物具有更高的抗氧化活性和營養價值,能澄清果汁并帶來清爽口感[31],因此后續酶解工藝研究加入聚半乳糖醛酸酶。
木聚糖酶預處理使低聚果糖含量達到22.57 g/L(P<0.05),提高12.98%。有研究表明,飲料中半纖維素的水解產物可使果汁澄清,水解阻礙果汁清除物理和化學物的物質或濃縮時引起渾濁的物質,增強漿汁的穩定性[32]。可見,加入木聚糖酶對菊芋漿存在有利影響,后續酶解工藝研究考慮加入木聚糖酶。
葡聚糖內切酶預處理使低聚果糖的含量達到22.09 g/L(P<0.05),提高10.51%。在葡萄酒工業中,葡聚糖內切酶可改善酒體顏色,提高浸漬度、澄清度和過濾性,從而提高了葡萄酒的質量;在發酵產品中加入葡聚糖內切酶,能提高產品質量和產量[33]。可見,葡聚糖內切酶對漿汁類產品澄清度和質量有一定提高,后續研究考慮加入葡聚糖內切酶。
各種組分對應水解酶的添加均表現出促進菊糖內切酶作用的效果。其中,單寧酶的促進作用最為明顯。考慮到其他組分對應的水解酶同樣可能受到單寧存在對酶活的抑制,后續酶解工藝研究中,采用單寧酶單獨預先處理菊芋漿,然后再添加其他組分對應的水解酶和菊糖內切酶進行組合酶解。
2.2.4 菊芋酶解工藝的建立與優化 為充分發揮各組分水解酶與菊糖內切酶組合酶解的效率,對各水解酶的添加量進行了逐一優化。結果如圖3~圖5。
2.2.4.1 各水解酶添加量的優化 由圖3可得出,當單寧酶的添加量為0.08 U/g時,低聚果糖的含量達到29.98 g/L(P<0.05),此時水解率達到60.48%,可能由于菊芋中含有影響菊糖釋放的物質,導致繼續增加單寧酶添加量后低聚果糖含量無明顯增加。因此,后續研究中,均加入0.08 U/g單寧酶預處理4 h,再進行下一步的組合酶解。
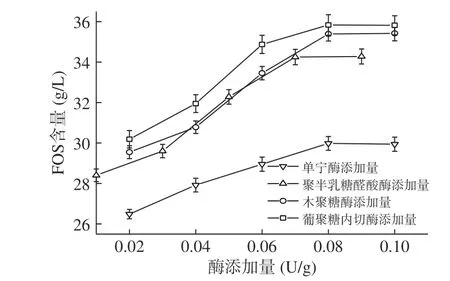
圖3 各水解酶添加量優化Fig.3 Optimization of each hydrolase enzyme addition
當聚半乳糖醛酸酶的添加量為0.07 U/g時,低聚果糖的含量達到34.25 g/L(P<0.05),此時水解率達到69.10%,而隨聚半乳糖醛酸酶添加量的增加低聚果糖含量無變化,可能是由于用量增加,酶蛋白使漿汁渾濁[34],影響菊糖內切酶發揮作用。因此,后續實驗聚半乳糖醛酸酶添加量均采用0.07 U/g進行組合酶解。
當木聚糖酶的添加量為0.08 U/g時,低聚果糖的含量達到最大值,為35.39 g/L(P<0.05),此時水解率為71.40%,低聚果糖含量較上一步有所提高,可能是木聚糖的降解使反應體系黏度降低[32],從而促進菊糖內切酶的酶解效率。因此,后續實驗木聚糖酶添加量均采用0.08 U/g進行組合酶解。
當葡聚糖內切酶的添加量為0.08 U/g時,低聚果糖的含量達到最大值,為35.83 g/L(P<0.05),此時水解率達到72.30%,低聚果糖含量進一步提高,可能是纖維素的降解,釋放了細胞內的蛋白、糖等物質,使菊糖更好的釋放[35],此外,葡聚糖內切酶與木聚糖酶、聚半乳糖醛酸酶結合,可改善果汁的黏度[32]。因此,后續實驗聚半乳糖醛酸酶添加量均采用0.08 U/g進行組合酶解。
2.2.4.2 菊糖內切酶添加量優化 為研究菊糖內切酶的添加量,以菊芋漿為底物,添加0.08 U/g單寧酶在50 ℃、pH5.0下酶解4 h后,再加入0.07 U/g聚半乳糖醛酸酶、0.08 U/g木聚糖酶、0.08 U/g葡聚糖內切酶,取不同濃度菊糖內切酶在50 ℃、pH5.0下酶解12 h。圖4表明,菊糖內切酶的用量為12.0 U/g時,低聚果糖含量達到最大值,為38.56 g/L(P<0.05),此時水解率達到77.79%,可能由于菊芋中菊糖沒有全部釋放,此時底物耗盡,繼續增加用酶量低聚果糖含量沒有明顯提高。因此,后續實驗聚半乳糖醛酸酶添加量均采用12.0 U/g進行組合酶解。

圖4 菊糖內切酶添加量優化Fig.4 Optimization of endoinulinase addition
2.2.4.3 菊糖內切酶酶解時間優化 為研究菊糖內切酶酶解時間,以菊芋漿為底物,添加0.08 U/g單寧酶在50 ℃、pH 5.0下酶解4 h后,再加入0.07 U/g聚半乳糖醛酸酶、0.08 U/g木聚糖酶、0.08 U/g葡聚糖內切酶、12.0 U/g菊糖內切酶在50 ℃、pH 5.0下酶解12 h,每隔2 h取樣一次,圖5結果表明,隨酶解時間增加,低聚果糖含量逐漸增加,當菊糖內切酶作用時間為8 h時,低聚果糖的含量達到最大值,為38.89 g/L(P<0.05),此時水解率達到78.46%,繼續延長酶解時間,低聚果糖含量增幅明顯減緩。因此,菊糖酶作用時間為8 h。

圖5 菊糖內切酶酶解時間優化Fig.5 Optimization of enzymatic hydrolysis time of endoinulinase
至此,本研究獲得富含低聚果糖的菊芋水解液的最優制備工藝為:先加入0.08 U/g單寧酶,50 ℃、pH5.0的條件下酶解4 h,再加入0.07 U/g聚半乳糖醛酸酶、0.08 U/g葡聚糖內切酶、0.08 U/g木聚糖酶、12.0 U/g菊糖內切酶50 ℃酶解8 h。此時低聚果糖的含量為38.89 g/L(P<0.05),水解率達到78.46%。而李鑫等[36]以菊芋渣為原料酶解制備的短鏈低聚果糖質量濃度為16.06 g/L,相比之下,本研究在復合酶的作用下制備的低聚果糖含量較高。
2.2.5 酶法制備低聚果糖糖漿的功能評價 將在上述最優工藝條件下得到的菊芋水解液濃縮為低聚果糖糖漿,并研究該糖漿的益生元功能,以低聚果糖糖漿為碳源,以未酶解的菊芋原液做對照,觀察植物乳桿菌(如圖6A)、嗜熱鏈球菌(如圖6B)、副干酪乳桿菌(如圖6C)的生長情況,由圖可知,這三種菌以低聚果糖糖漿為碳源時,均有明顯的增殖,且生長速率分別提高了33.33%、60.47%、148.15%,最大生物量比陳韞慧等[28]的研究相比效果較好,說明該工藝條件下的低聚果糖糖漿有利于益生菌的生長,具有益生元功能,可作為低糖高營養食品的原料。

圖6 三種菌的生長曲線Fig.6 Growth curves of three species of bacteria
進一步檢測該糖漿組分含量及抗氧化活性,結果見表3,制備的糖漿中低聚果糖含量達到53.73 g/L,單寧和總蛋白含量都有所提高。此外,酶處理可顯著提高菊芋糖漿的抗氧化活性(P<0.05),DPPH自由基清除率、羥基自由基清除率和T-AOC分別達到82.23%、30.47%和2.78 μmol/mL,與標準品維生素C相比,其抗氧化活性處于較高水平。
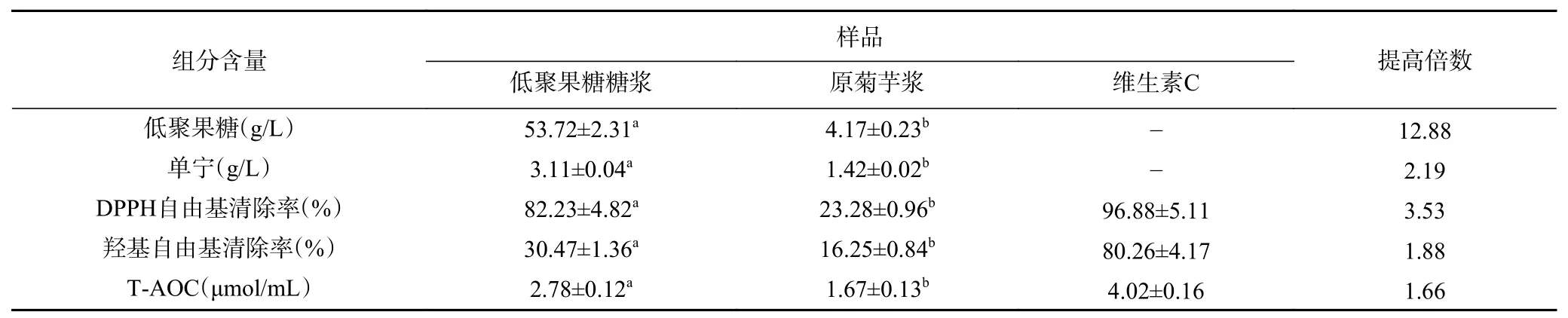
表3 低聚果糖糖漿的組分含量及抗氧化活性Table 3 Composition content and antioxidant activity of oligosaccharides syrup
有研究表明,單寧酶水解單寧的水解產物(表兒茶素、表沒食子兒茶素和沒食子酸)比底物(表兒茶素沒食子酸酯和表沒食子兒茶素沒食子酸酯)具有更高的抗氧化活性[37-38],可見,單寧酶對菊芋原漿的預酶解,既有助于解除單寧對菊糖內切酶的抑制作用,又有助于抗氧化物質的釋放。
3 結論
本研究以新鮮菊芋為原料,通過研究菊芋內源酶、菊粉內切酶、葡聚糖內切酶、木聚糖酶、聚半乳糖醛酸酶和單寧酶在鮮菊芋酶法加工制備制備低聚果糖糖漿中的影響,建立并優化酶法制備低聚果糖糖漿的工藝。結果表明,復合酶解得到的菊芋水解液中,低聚果糖含量達到38.89 g/L,經濃縮制備的低聚果糖糖漿中低聚果糖和單寧的含量分別為53.72和3.11 g/L,DPPH自由基清除率、羥基自由基清除率和總抗氧化能力分別為82.23%、30.47%和2.78 μmol/mL,抗氧化活性較原漿有所提高,此外,能促進益生菌生長,具有益生元功能。菊芋的這種酶處理工藝可能成為未來菊芋種植業綜合加工、提升產業價值的重要方向之一。
