基因工程表達甜味蛋白的研究進展
2023-01-13 13:16:22施天元曹國強
食品工業科技 2023年2期
關鍵詞:生產
施天元,曹國強
(黑龍江省金象生化有限責任公司,黑龍江哈爾濱 150040)
甜味蛋白作為一種由植物提取的蛋白質,具有甜度大、熱量低、安全無毒等特點,早在上世紀就受到了廣泛的關注[1-2]。甜味蛋白是一類擁有巨大潛力的天然甜味劑,可作為蔗糖替代物,它們熱量低,功能活性好,經胃蛋白酶消化可成為必需氨基酸,具有營養價值[3],但由于最初的植物提取存在原料產地限制、提取成本高等問題,限制了其大規模的生產和使用[2,4],利用基因工程異源表達甜味蛋白引起了科學家們的興趣。
目前,甜味蛋白的基因工程改造主要應用于增強甜味蛋白的穩定性、增加甜度、提高產量、降低成本等[5]。但同時,基因工程表達生產甜味蛋白尚存在較多困難與挑戰。如蛋白翻譯后修飾的提取工藝復雜,成本高,價格無法與市場上的人工合成甜味劑相競爭。利用細菌、真菌為原料生產的甜味蛋白很難被大眾接受應用于食品等[6]。基因工程生產的甜味蛋白具有傳統植物原料提取生產無法比擬的優點,包括更高的產率、更低的理論成本和更有針對性的改造,代糖食品已成為新趨勢[7],但是甜味蛋白作為已發現的最甜的物質,現仍未被廣泛應用。本文首次系統的分析了基因工程在生產甜味蛋白中的優勢、應用及相關熱點問題,為未來基因工程改造甜味蛋白提供理論資料。
1 甜味蛋白的種類及理化性質
1.1 索馬甜
索馬甜又稱奇異果甜蛋白、沙馬汀,見表1。最初提取自一種來自西非的植物(Thaumatococcus daniellii)。甜度是蔗糖的3000倍。據報道,索馬甜家族至少存在五種常見的高甜度的索馬甜蛋白成員,包括兩種天然高含量的蛋白產物索馬甜I和索馬甜II,還有三種微量的蛋白產物索馬甜a,索馬甜b和索馬甜c。盡管有些肽環有差異[8],但所有的蛋白都具有相同的分子量大小22 kDa,和相同的等電點pH12[9]。索馬甜I和索馬甜II蛋白都含有207個氨基酸,之間存在五個氨基酸位點的差異,分別是N46K、S63R、K67R、R76Q和N113D[10]。索馬甜蛋白含有16個半胱氨酸形成8個二硫鍵[11]。
1.2 莫乃靈
莫乃靈是一種存在于西非植物(Discorephyllum cumminisii)中的甜味蛋白,見表1。甜度較高,是蔗糖的3000倍。天然莫乃靈由94個氨基酸組成,含有一個半胱氨酸,不存在二硫鍵[11],其分子量為10.7 kDa[4,12]。包含兩個非共價連接的多肽鏈鏈A和鏈B分別包括44個氨基酸和50個氨基酸。單獨的鏈A和鏈B都不具有甜味,見表1[13]。莫乃靈的二級結構包含一個反向平行的β-折疊和一個17個殘基的α-螺旋[12]。因為莫乃靈A鏈和B鏈通過弱共價鍵鏈接,導致其熱穩定性較低[14]。MNEI是莫乃靈的單鏈衍生物,具有相似的甜度[15],相比于莫乃靈,具有更強的熱穩定性[16]。
1.3 馬賓靈
馬賓靈,也叫馬檳榔甜蛋白,是從亞熱帶植物(Capparis masaikai)種子中分離得到的,見表1。馬賓靈的甜度是蔗糖的100倍。馬賓靈是由A鏈和B鏈組成的二聚體,A鏈由33個氨基酸組成,B鏈由72個氨基酸組成。共有四種同分異構體,1、3、4型均不能穩定存在,只有2型是熱穩定的,2型馬賓靈蛋白也是目前有報道的研究最多的馬賓靈蛋白[6,17],其甜味能在80 °C條件下保持至少2 h[2,4]。馬賓靈的分子量為12.4 kDa[4]。2型馬賓靈前體含有155個氨基酸殘基,包含一個20個氨基酸的信號肽、一個15個氨基酸的N端擴展序列、一個14個氨基酸的A鏈和B鏈之間的連接多肽和一個C端的擴展氨基酸[18]。馬賓靈在已知的六種甜味植物蛋白中表現出最顯著的熱穩定性和耐酸性[19]。
1.4 布拉齊因
布拉齊因,也叫布那珍甜蛋白,存在于一種西非常綠灌木(Pentadiplandra brazzeana)的紅漿果中[20],見表1。布拉齊因具有很高的溶解性,在pH大幅度變化和高溫情況下都很穩定[21]。布拉齊因比2%的蔗糖溶液甜2000倍,比10%的蔗糖溶液甜500倍。它的味道更像蔗糖而不是索馬甜[22]。它的甜度在80 ℃ 4.5 h條件下也不會被破壞[23]。布拉齊因由54個氨基酸組成,分子量為6473 Da[24]。果實中還存在一種微量的布拉齊因蛋白,由53個氨基酸殘基組成,甜度是主要成分的兩倍[22]。結構區別為主要存在形式在氨基末端有焦谷氨酸,而微量形式在氨基末端缺少焦谷氨酸[25]。布拉齊因的二級結構包含一個α-螺旋(殘基21~29)和三個反向平行的β-折疊(鏈I,殘基5~7;基鏈II,殘基44~50;鏈III,殘基34~39)。四個二硫鍵相互交聯成次級結構[26]。潘塔亭(pentadin)也是一種甜味蛋白[27],但有人認為它只是布拉齊因的二聚體形式[22,27]潘塔亭由54個氨基酸組成,甜度為蔗糖的500倍。潘塔亭的甜味很容易喪失[2]。
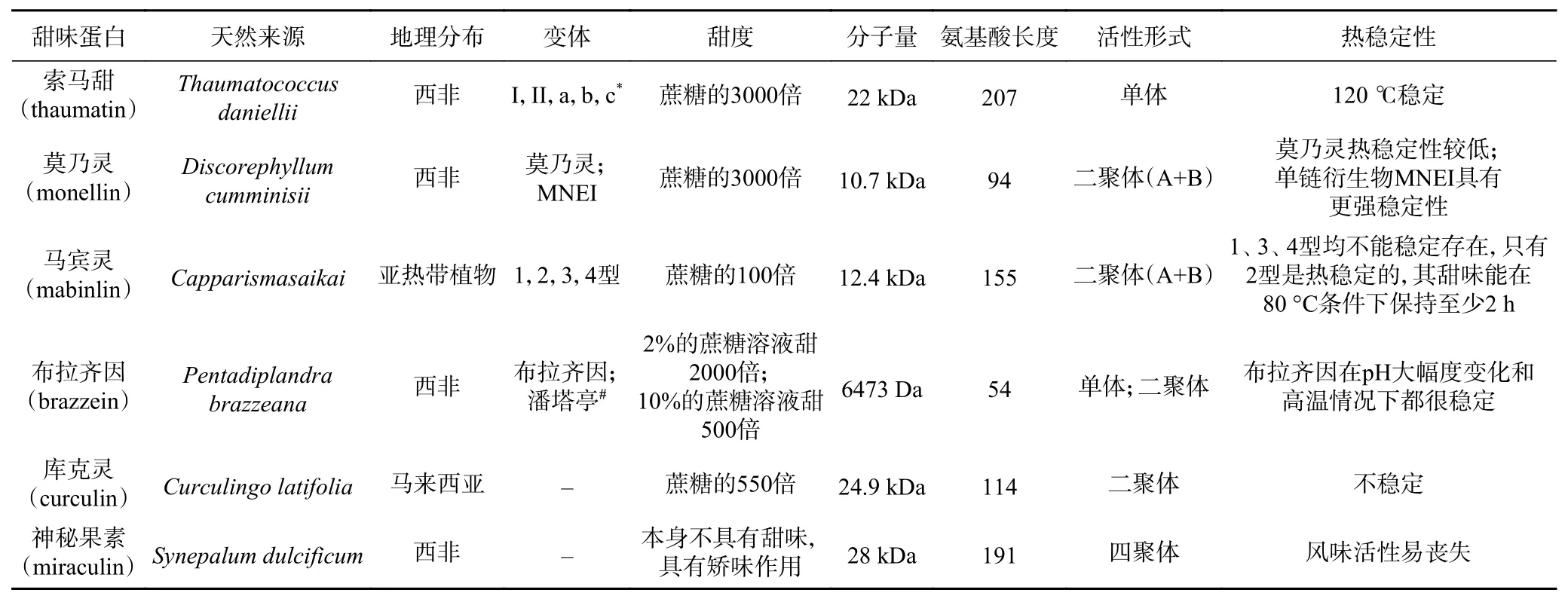
表1 不同甜味蛋白特性對比Table 1 Comparison characteristics of different sweet proteins
1.5 庫克靈
庫克靈,也叫仙茅甜蛋白,存在于馬來西亞的一種植物(Curculingo latifolia)中,見表一[28]。甜度是蔗糖的550倍。其分子量24.9 kDa,由114個氨基酸組成。庫克靈是一種由兩個基礎亞基組成的同源二聚體[10]。庫克靈不穩定,在50 ℃及以上溫度活性降低[2]。庫克靈也是一種矯味蛋白,能夠使酸味物質變甜[7]。甜味在口腔內可維持幾分鐘;甜味在口腔內消失后若喝水,又會恢復,并可維持5 min左右[27]。Neoculin與庫克靈具有相似的甜度,甜度大約是蔗糖的500倍[29]。與庫克靈的區別是Neoculin是一種由一個酸性亞基(NAS)和一個基礎亞基(NBS)組成的異源二聚體。
1.6 神秘果素
神秘果素,也叫奇果蛋白,存在于一種熱帶西非灌木(Synepalum dulcificum)的果實中,見表1。神秘果素本身不具有甜味,但可以使酸味或苦味物質嘗起來具有甜味[30],包括鹽酸、草酸、乳酸、甲酸、醋酸、檸檬酸等。增甜效果取決于酸的酸度和pH[31]。濃度為4×10-7mol/L時神秘果素即能被感知,其甜度相當于0.4 mol/L的蔗糖溶液[32]。在1988年,Theerasilp和Kurihara成功地分離出純化的神秘果素甜味蛋白,并測定其分子量為28 kDa。神秘果素是一個具有191個氨基酸的單鏈多肽,具有兩個通過二硫鍵交聯的糖基化位點,N42和N186[30]。
2 利用基因工程生產甜味蛋白的優勢
2.1 甜度方面
利用玉米表達系統生產甜味蛋白布拉齊因,表達量高,純化出的玉米布拉齊因蛋白甜度高達蔗糖的1200倍[33]。而另一方面,天然索馬甜引起甜味味覺感受的濃度為50 nmol/L[2,34]。通過基因工程改造,最甜的索馬甜突變體D21N,31 nmol/L濃度就能引起味覺[34]。MNEI兩個雙突變體E50N/Y65R和E2N/E50N均表現出比單突變體E50N更高的甜度[35]。用大腸桿菌表達的Neoculin不僅能誘發甜味反應,而且能在水或檸檬酸存在情況下進行矯味[29]。
2.2 蛋白活性方面
神秘果素在室溫收獲后的2~3 h內,風味活性就會喪失[36]。因此,神秘果素的天然來源是有限的。人們利用外源的大腸桿菌、酵母菌、米曲霉和植物等宿主,一直在嘗試利用基因工程技術生產神秘果素,利用轉基因生菜,成功表達了具有甜味活性的重組神秘果素蛋白,以生菜為宿主表達神秘果素蛋白不僅解決了神秘果素易喪失風味的問題,而且由于其食品來源消除了人們對其安全性的擔心[37]。利用帶有KEX2剪切位點的α-淀粉酶分別與兩個蛋白亞基融合,可成功獲得與天然Neoculin具有同樣活性的重組Neoculin[27]。布拉齊因需要四個二硫鍵才能激活產生強烈的甜味,而利用畢赤酵母獲得的布拉齊因的不僅具有活性且在產量方面具有優勢[21]。
2.3 蛋白表達量與轉化率方面
酵母,主要為畢赤酵母(Pichia pastoris)和釀酒酵母(Saccharomyces cerevisiae),在產量方面由于其為真核微生物,在合成和分泌方式上具有優勢。索馬甜用合成DNA在釀酒酵母中生產,表達水平可達140 mg/L。畢赤酵母生產索馬甜產量也可達到70 mg/L[18]。在畢赤酵母中共表達索馬甜與分子伴侶使產量至少提高了兩倍[38]。另外,乳酸乳球菌也可用于表達索馬甜[2]。莫乃靈可以除了可用畢赤酵母和釀酒酵母表達外,還可以用產朊假絲酵母(Candida utilis)表達[39]。利用產朊假絲酵母生產的莫乃靈產量甚至可以超過天然提取的莫乃靈[6]。索馬甜在米曲霉(Aspergillus oryzae)、婁地青霉(Penicillium.roquefortii)、黑曲霉變種(Aspergillus. nigervar.awamori)中均能成功表達[40-41]。在酸性蛋白酶缺陷的黑曲霉變種中,gdhA啟動子誘導和適宜的N源C源條件下,索馬甜的產量可達105 mg/L[40]。同樣的宿主中過表達pdiA基因,產量可達150 mg/L[41]。
在番茄植株中表達并純化出了與天然蛋白甜味相同的索馬甜II,重組的索馬甜II與天然索馬甜II的口感特征沒有明顯區別且具有加工生產發面的優勢[42]。轉基因的番茄也是最適合生產重組神秘果素的宿主,以幼苗子葉為外植體的農桿菌介導轉化方案中,番茄轉化效率超過40%[43]。
目前,評價因素定量化篩選常用的方法有:層次分析法、信息熵法、主成分分析法、聚類分析法、灰色關聯分析法、神經網絡法、粗糙集屬性約簡法。充分考慮核電廠主控室龐大的系統環境,采用層次分析法。目前層次分析法主要采用領域專家意見,主觀賦值通常需要有一定的先驗知識,評判結果具有主觀隨意性,且核電廠交互界面的設計需要考慮到界面設計師以及核電專家兩方面人士的意見,評判側重點容易產生分歧且存在主觀性,故本文采用基于粗糙集理論的層次分析法,自動獲取評價因子。
3 基因工程表達生產甜味蛋白遇到的困難及解決辦法
3.1 甜味缺失
甜味蛋白最重要的是甜味。但1982年,Luppo等[10]第一次在大腸桿菌中表達出的索馬甜蛋白是沒有甜味的。究其原因,主要是由于蛋白質折疊不當或缺乏必要的翻譯后的修飾,因而不能引起甜味反應[6,44]。人們利用外源的大腸桿菌、酵母菌、米曲霉和植物等宿主,一直在嘗試利用基因工程技術生產神秘果素。但異源生產的神秘果素有的喪失了甜味,有的甜味活性很低[18,37,45]。用大腸桿菌表達的庫克靈(NBS-NBS)或庫克靈2(NAS-NAS)兩種同源二聚體都不具有甜味,也沒有矯味作用。2000年在前人基礎上,通過氧化還原性谷胱甘肽系統對重組蛋白進行復性處理,生產出了具有與天然索馬甜相似甜味的甜味蛋白索馬甜[46-47]。利用轉基因植物作為宿主可實現具有甜味活性的神秘果素的生產[30]。MNEI是一種高效的飲料甜味劑,但在半固態食品中使用可能會降低它的甜味,已證實對其進行點突變可以解決這類問題[34,48]。
3.2 成本高
植物提取甜味蛋白由于原料限制存在成本高的問題,限制了其大規模的應用[3,19]。2020年,Park等[49]以胡蘿卜(Daucus carotaL.)愈傷組織為材料,氧化應激誘導SWPA2啟動子驅動各種應激處理下神秘果素基因的表達,采用氣升式生物反應器進行了重組神秘果素的大規模生產研究,為將來利用轉基因胡蘿卜愈傷組織量產神秘果素打下基礎。此外,利用CaMV35S啟動子過表達編碼甜味蛋白基因的轉基因土豆、生菜、黃瓜、草莓等也獲得了成功[18]。部分重組甜味蛋白如莫乃靈仍處于研究階段,也存在生產成本高,在產量和價格上與市場上的人工合成甜味劑相比具有較大差距的問題[1]。利用生物技術生產植物甜味蛋白和矯味蛋白必須達到一定的單產水平才能與天然產品競爭[6]。原核宿主相比植物表達具有操作流程簡單、生長周期短等優點,但是原核宿主表達也存在純化過程繁瑣,純化工藝復雜,純化成本高的問題,而且部分甜味蛋白無法正確折疊,產生甜味[49]。于是人們開展了轉基因植物組織和真菌表達平臺生產甜味蛋白的研究。開發的細胞培養流程簡化了純化過程,提高了蛋白質產量,節約了成本。利用煙草毛狀根培養物成功表達并分泌了甜味蛋白索馬甜I[50]。
3.3 穩定性差
莫乃靈A鏈和B鏈通過弱共價鍵鏈接,熱穩定性較低[14]。基因工程提高部分甜味蛋白如莫乃靈的熱穩定性也是需要解決的問題[1]。MNEI與莫乃靈具有相似的甜度[51],克隆相對簡單,可導入許多宿主微生物中生產應用于食品飲料工業[14]。也可以利用轉基因植物,固相多肽合成等方式生產[18]。相比于莫乃靈,MNEI具有更強的熱穩定性,因此也有著更廣泛的應用。現階段主要用MNEI替代莫乃靈,同時MNEI也被認為是最有前途的甜味蛋白之一[18]。
3.4 安全性
利用基因工程技術生產出的重組蛋白的安全性需經過嚴格的生物學活性檢驗[52]。前期人們在大腸桿菌和酵母中表達了重組甜味蛋白,雖然也進行了小鼠的毒理學驗證,但是并未得到所有人的認可,相當一部分人還是不能接受尤其是在大腸桿菌中表達的甜味蛋白應用于食品[6,53-54]。為甜味蛋白尋找更多被認可的安全表達宿主也是甜味蛋白開發面臨的重要問題之一。還有,利用重組技術生產的植物甜味蛋白或矯味蛋白必須獲得權威性食品安全檢驗機構的認證,例如美國的FDA和歐盟的食品安全審查機構的認可。食品安全認證也是生產食品級甜味蛋白需要攻克的關鍵問題[6]。由于乳酸乳球菌(Lactococcus.lactis)屬于常見的腸道定殖菌,幾百年來一直被奶酪制造工業使用,因此用它生產的MNEI被大多數人認為是安全的可食用甜味蛋白[54]。目前,索馬甜和莫乃靈已通過毒理學試驗和安全試驗,經相關批準用于食物和飼料的增甜劑或調味劑[52,54]。
3.5 大規模生產受限
盡管在一些生物體中成功地表達了重組索馬甜蛋白,但一直缺少大規模的生物生產設施存在。植物的價格,生長周期,種植密度在大規模生產中均需要考慮。此外,由于氣象因素,產品產量和質量方面可能存在大量的批內差異,這也是使用田間種植的植物材料進行這一應用所關注的問題之一。這些變化反過來會導致關鍵設施性能參數的不一致。2021年Kelada等[55]提出了初步的工藝設計,工藝模擬和經濟分析,將利用多種農產品生產重組索馬甜蛋白。未來預計將采用分子手段每年量產50公噸索馬甜II。
另外,酵母是最具工業意義的真核微生物,由于其可用性,主要應用于生物生產,其具有良好的遺傳和生理背景,遺傳操作簡單,生長迅速,具有翻譯后修飾和細胞外分泌的能力。這樣的平臺使其成為表達甜味蛋白的理想系統。因此,利用微生物如絲狀真菌、酵母等對酶的高水平分泌和重組蛋白的大規模生產有一定的好處[2]。
4 基因工程生產甜味蛋白技術的應用
4.1 索馬甜的生產應用
4.2 莫乃靈的生產應用
莫乃靈也是美國食品和藥物管理局(FDA)批準的“公認安全的”(GRAS)食品添加劑[7]。莫乃靈由于高溫不耐受和pH條件的限制[12],其研究一直處于瓶頸狀態。對其進行基因工程改造可提高其穩定性和表達水平,降低其生產成本[57],改良口感,提高產量[47,58]。最初利用基因工程表達的重組莫乃靈蛋白需要生長在含有甲醇的培養基上[59]。在2016年,分子生物學研究表明莫乃靈的N端序列決定了其在酵母菌株中表達的蛋白甜度,成功在畢赤酵母系統中表達了不需要甲醇誘導的莫乃靈蛋白,此項研究表明莫乃靈的量產不再是遙不可及[59]。Jia等[60]通過有效調控甲醇代謝模式和提高能量利用效率,提高了畢赤酵母中低細胞濃度條件誘導的莫乃靈產量。應用Trp啟動子或T7 lac啟動子在大腸桿菌中表達莫乃靈或MNEI蛋白,表達水平高達45%[10,14]。2015年,用乙酸作為碳源工業化生產MNEI在理論上已完全具備條件,并且產量可達177 mg/L[61]。2018年,該實驗室又通過生產乳制品廢液中的乳糖為底物,開發了一種工藝簡單,適合工業規模應用,并且生長參數易于控制的MNEI生產流程[54]。除此之外,煙草葉綠體也可以作為穩定的植物表達平臺來生產三種MNEI蛋白突變體,它們具有更好的口感、更高的穩定性和產量[58]。
4.3 其他甜味蛋白的生產應用
不僅僅是索馬甜與莫乃靈,其他甜味蛋白通過基因工程的改造工作也已經在不斷推進。2004年Lamphear等[33]在玉米宿主中表達了布拉齊因蛋白,因含布拉齊因的玉米胚芽粉可直接應用于食品,使這種高甜度的甜味蛋白未來在食品工業中的作用得到看好,并且使用可食用植物作為宿主的優勢在于可以降低純化成本[32]。美國的BioResources International公司也利用傳統的栽培技術種植R.dulifera,并利用重組宿主表達生產神秘果素。作為合作研究,日本的Asahi DenkaKogyo公司和日本物理學和化學研究所正在將庫克靈表達于轉基因植物和大腸桿菌。而布拉奇因作為甜度最大的甜味蛋白,也擁有著極大的潛力,近年來被廣泛研究。通過基因工程改造后的布拉奇因甜度比野生型提高近50%[5]。乳酸克魯維酵母表達系統生產的布拉奇因產量可達107 mg/L[62]。經優化發酵工藝、凈化程序后的布拉奇因可以用作食品、醫藥和生物技術領域的甜味添加劑[23],在食品工業中具有廣闊的應用前景。
5 結語與展望
目前關于甜味蛋白的研究主要集中在甜味活性的保持,降低生產成本,提高產量,生產工藝的優化等方面。研究人員希望能夠找到一種認可度高、高甜度、價格便宜、甜味特性好、性質穩定的甜味蛋白。利用可食用的植物組織生產甜味蛋白在安全性上具有一定優勢,易于被大眾接受。利用微生物生產的甜味蛋白具有價格上的優勢。而無論是利用植物組織生產還是微生物生產,應用基因工程改造都能令其具有更高的甜度和更好的穩定性。
2016年,重組蛋白市場全球價值3.472億美元。根據2018年consistent Market Insights公司提供的信息,2017~2025年全球重組蛋白市場復合年增長率預計為6.2%[2]。甜味蛋白具有甜度高,熱量低,安全,且不會把非天然的代謝產物引入體內的優點。隨著人們健康意識的增強,甜味蛋白未來將長期向好。我國目前減糖市場還屬于起步探索階段,作為功能糖和高倍甜味劑原料的主產國,國內企業發展減糖類產品具有較大的原料優勢。制糖企業可參考國際減糖產品發展趨勢及甜味劑的應用情況提前布局減糖市場[7]。隨著部分甜味蛋白被發達國家批準應用,未來甜味蛋白的國際需求將持續增大,關于甜味蛋白的研究也將越來越多。
甜味蛋白的研究方向一直集中在降低成本、提高轉化率、提高穩定性、擴大高活性甜味蛋白的生產規模上。未來,繼續提高甜度,降低生產成本將是主流方向。此外,為克服原材料的季節性,通過基因工程手段對植物甜味蛋白基因進行克隆,再導入微生物細胞表達,最后利用工業發酵法來大規模生產或將成為未來生產甜味蛋白的主流模式;另外,純化方式的優化也是一大難點。在應用方面,未來也將使用多種甜味劑復合產品改良食品口感。同時,隨著科技的進步,甜味蛋白的研究也將向更為微觀,更為精細發展,現已開展對甜味蛋白活性位點的研究,也就是說,生物工程技術和計算機技術設計相結合改良甜味蛋白也將成為趨勢。
猜你喜歡
江蘇安全生產(2022年9期)2022-11-02 07:01:24
中國化肥信息(2022年7期)2022-08-31 01:28:54
山東冶金(2022年2期)2022-08-08 01:50:42
小學科學(學生版)(2020年10期)2020-10-28 07:52:12
中國化肥信息(2020年7期)2020-03-19 01:54:02
中國軍轉民(2017年6期)2018-01-31 02:22:28
消費導刊(2017年24期)2018-01-31 01:29:23
中國制筆(2017年2期)2017-07-18 10:53:09
現代企業(2015年4期)2015-02-28 18:48:06
汽車零部件(2014年11期)2014-09-18 11:57:16
