基于群體感應的弧菌生物膜形成、耐藥性及其控制研究進展
2023-01-13 13:16:22劉雪晴呂欣然白鳳翎勵建榮
食品工業科技 2023年2期
劉雪晴,呂欣然,2, ,白鳳翎,勵建榮
(1.渤海大學食品科學與工程學院,遼寧錦州 121013;2.大連工業大學,海洋食品精深加工關鍵技術省部共建協同創新中心,遼寧大連 116034)
弧菌是菌體短小,彎曲成弧形,尾部帶一鞭毛的革蘭氏陰性菌,被認為是一種常見的魚類病原菌[1],主要包括霍亂弧菌(V. cholerae)、副溶血弧菌(V.parahaemolyticus)、創傷性弧菌(V. vulnificus)、哈維氏弧菌(V. harveyi)、溶藻性弧菌(V. alginate)等[2-3]。霍亂弧菌、副溶血弧菌與創傷性弧菌是常見的致病性弧菌,在攝入被污染的海產品后可造成霍亂及食物中毒[4-6]。溶藻性弧菌與哈維氏弧菌主要感染蝦及魚類,可導致魚、蝦、貝類等在育苗和養殖過程中暴發疾病甚至大量死亡,以及在生產運輸過程中發生產品腐敗變質,經濟損失嚴重[7-8]。為防止弧菌污染海產品,常使用抗菌藥物,但藥物的大量使用使弧菌的耐藥性明顯增強,致使其污染海產品的概率大大增加,直接或間接導致了水產養殖業、水產品加工業以及其他相關產業的經濟損失[9]。
群體感應(Quorum Sensing,QS)是指細菌通過監測種群群體密度進而相應地調節其基因表達的行為,是細菌群體之間進行通訊和交流的一種機制[10]。一種微生物可擁有多套群體感應系統、產生多種群體感應信號分子,協調或調控微生物的多種生物功能。其中最主要的是群體感應能夠通過特定的調節因子或與其他信號分子共同作用來調節弧菌的生物膜形成和毒力因子的表達[11]。副溶血性弧菌可以產生兩種類型的IV型菌毛:甘露糖敏感型血凝素IV型菌毛(MSHA)和幾丁質調節菌毛(ChiRP)。它們都是生物膜形成所必需的。研究發現,副溶血性弧菌群體感應調節因子AphA和OpaR對副溶血性弧菌MSHA菌毛中編碼菌毛蛋白的mshA1轉錄起相反的調節作用。在低細胞密度(LCD)下,AphA間接抑制mshA1轉錄。在高細胞密度(HCD)下,OpaR激活mshA1轉錄,促進了生物膜的形成[12]。與大部分弧菌不同的是,霍亂弧菌群體感應系統通過調控LuxR同源物HapR,使其能夠與DNA直接結合,抑制其表面多糖合成基因vps的轉錄及表達,從而抑制生物膜的形成[13]。生物膜(Biofilm,BF)是由細菌及其自身代謝產生的胞外基質(Extracellular Polymeric Substances,EPS)(多糖、蛋白質、eDNA)粘附在某些生物或非生物材料表面,保護細菌正常生長的一種膜狀結構群[14]。弧菌生物膜是海產品在捕撈與加工期間形成的環境與細菌之間的屏障,從而降低細菌對一些抗菌藥物的敏感性[15]。有研究表明,大部分弧菌生物膜對β-內酰胺類、氨基糖苷類、大環內酯類、喹諾酮類等抗生素有不同程度的耐藥[16]。
由于生物膜的廣泛性和危害性,研究人員正積極地尋找新途徑來預防和控制弧菌生物膜的形成[17]。本文針對弧菌生物膜與其群體感應系統的相關性,重點闡述了弧菌生物膜形成對耐藥性的影響,并對化學合成類、植物源和微生物源等群體感應抑制劑控制弧菌生物膜的研究現狀進行總結,為預防和控制弧菌生物膜形成奠定理論基礎。
1 弧菌的群體感應系統和生物膜
1.1 弧菌的群體感應系統
弧菌的QS系統調控細菌生物發光、生物膜形成以及毒性基因表達等性狀。弧菌群體感應系統主要包括:a.HAI-1型,該系統由N-酰基高絲氨酸內酯類(N-acylated homoserine lactones,AHLs)信號分子、AHLs合成酶(LuxM蛋白)和AHLs受體(LuxN蛋白)三部分組成,介導細菌種內通訊[18];b.AI-2型,是一種用于細菌種間交流的系統,信號分子是由LuxS蛋白合成的呋喃硼酸二酯(AI-2),當AI-2在菌體外積累到一定閾值時,其與LuxP受體蛋白結合后,再與激酶蛋白LuxQ反應啟動相關毒力基因的表達[18];c.CAI-1型。信號分子為S-3-羥基十三烷-4-酮(CAI-1),CqsA是CAI-1合成酶蛋白,CqsS是CAI-1的組氨酸激酶受體[19]。費氏弧菌和哈維氏弧菌中發現HAI-1、AI-2和CAI-1三種類型QS系統,霍亂弧菌中包含CAI-1和AI-2兩種QS系統,而創傷弧菌只有一條AI-2型QS系統[16]。由此可知,同屬不同種的弧菌之間群體感應系統存在一定的差異性。
1.2 群體感應系統直接介導生物膜形成
弧菌生物膜易在海產品表面及其加工設備中因細菌沉降而形成。其形成過程一般包括4個階段,如圖1所示,即細菌的初始粘附、不可逆附著、成熟和老化階段,該過程是動態變化的,且易受多種因素影響[20]。弧菌QS系統和環境信號應答系統共同調控生物膜的形成和成熟。當弧菌生長在不利的環境條件下時,可誘發其環境信號應答系統,使弧菌分泌QS信號分子,當信號分子累積達到一定閾值時,就會與受體蛋白結合,從而改變細胞內的調控元件轉錄或表達,促進EPS的合成,增強細菌的粘附能力,加速生物膜成熟與結構的穩定。弧菌QS系統中調控生物膜形成的終點控制因子LuxR及其同源物,可通過結合其靶基因啟動子并激活生物膜相關基因的轉錄來調控生物膜的形成[21-22]。副溶血弧菌在不同的環境條件下通過切換菌落類型,表現出強的適應機制。其QS通路中luxR的同源基因opaR基因可以正向調節副溶血弧菌菌落不透明性和粘附因子的表達,并影響生物被膜的形成。當opaR過表達時,可以促進菌落混濁,進而促進生物膜的成熟[23]。在創傷弧菌和費氏弧菌中也觀察到類似的現象[24]。以哈維氏弧菌為例,其QS系統調控生物膜形成如圖2所示,在低自體誘導物濃度下,LuxN、LuxQ和CqsS等信號分子受體蛋白將磷酸基團傳遞到共用的磷酸轉移酶的LuxU,進而傳遞到LuxO。磷酸化的LuxO可激活同源群體感應調控型小RNAs(sRNAs)基因(qrr)的轉錄,并與RNA結合蛋白Hfq一起抑制LuxR的mRNA翻譯。在高自體誘導物濃度下,HAI-1、AI-2和CAI-1與相應的受體蛋白結合,使LuxO去磷酸化失去活性,導致sRNAs不能被轉錄,使得編碼LuxR的mRNA得到穩定翻譯,從而調控生物膜的形成。
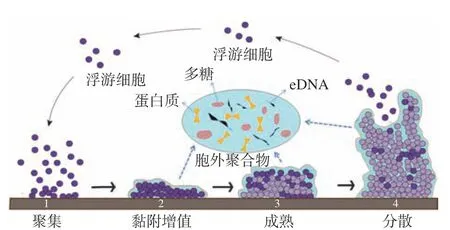
圖1 細菌生物膜形成過程Fig.1 Biofilm formation process of bacterials
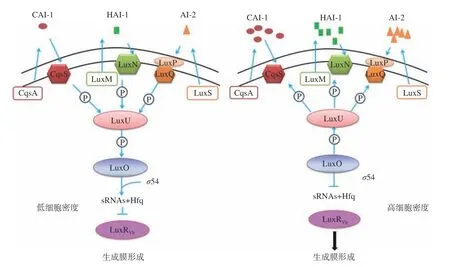
圖2 哈維氏弧菌群體感應系統及調控生物膜形成機制Fig.2 Quorum sensing system and the mechanism of biofilm formation of Vibrio harveyi
1.3 群體感應系統協同其他信號分子介導生物膜形成
除了基于QS系統調控外,弧菌中還存在著同樣可介導生物膜形成的其他信號分子,在生物膜形成階段中有著重要作用。環鳥苷二磷酸(cyclicdiguanylica cid,c-di-GMP)作為內部信使幾乎存在于所有細菌中,其介導革蘭氏陰性菌在運動和附著間的轉換,是生物膜形成的中心調節因子[25]。c-di-GMP通過增加胞外多糖的產生,調節細胞大小,增加細胞聚集,減少鞭毛泳動和eDNA生成來促使細菌生物膜形成[26]。c-di-GMP調控生物膜形成時受控于群體感應系統,Waters等[13]研究發現霍亂弧菌在高細胞密度下,QS調控HapR蛋白合成,它可抑制c -di-GMP合成基因和生物膜轉錄激活因子vpsT的表達,進而抑制生物膜的形成;而低細胞密度時,hapR的轉錄受到抑制,c-di-GMP不斷進行生物合成,濃度增加,誘導生物膜轉錄激活因子vpsT的表達,進而促進生物膜的形成。這一發現證明了在QS信號分子未達到閾值時,c-di-GMP調控著細菌的運動聚集、增殖、營養吸收、合成EPS等生理功能,隨著細胞密度增高并到達閾值,QS系統開始調控生物膜黏附、成熟與分散過程,進而印證了霍亂弧菌中c-di-GMP與雙組分系系統、QS系統等組成了復雜的信號傳遞網絡,共同介導細菌生物膜的形成。
2 生物膜形成對弧菌耐藥性的影響
殺菌劑是指可滅活幾乎所有公認的病原微生物的化學試劑的統稱。抗生素是殺菌劑中的一類,是指微生物在繁殖過程中所產生的具有抗病原體或其它活性的一類次級代謝產物,具有靶向特異性。弧菌成熟生物膜對抗生素或其他類殺菌劑均具有抵抗力。生物膜可阻止殺菌劑進入細胞或在進入細胞前使殺菌劑失去效用,這種方式在對殺菌物質的抗性機制中起到重要作用[27]。因此,研究弧菌生物膜形成對弧菌耐藥性的影響是解決當前細菌耐藥問題的有效途徑。如圖3所示,其影響主要包括以下三個方面。
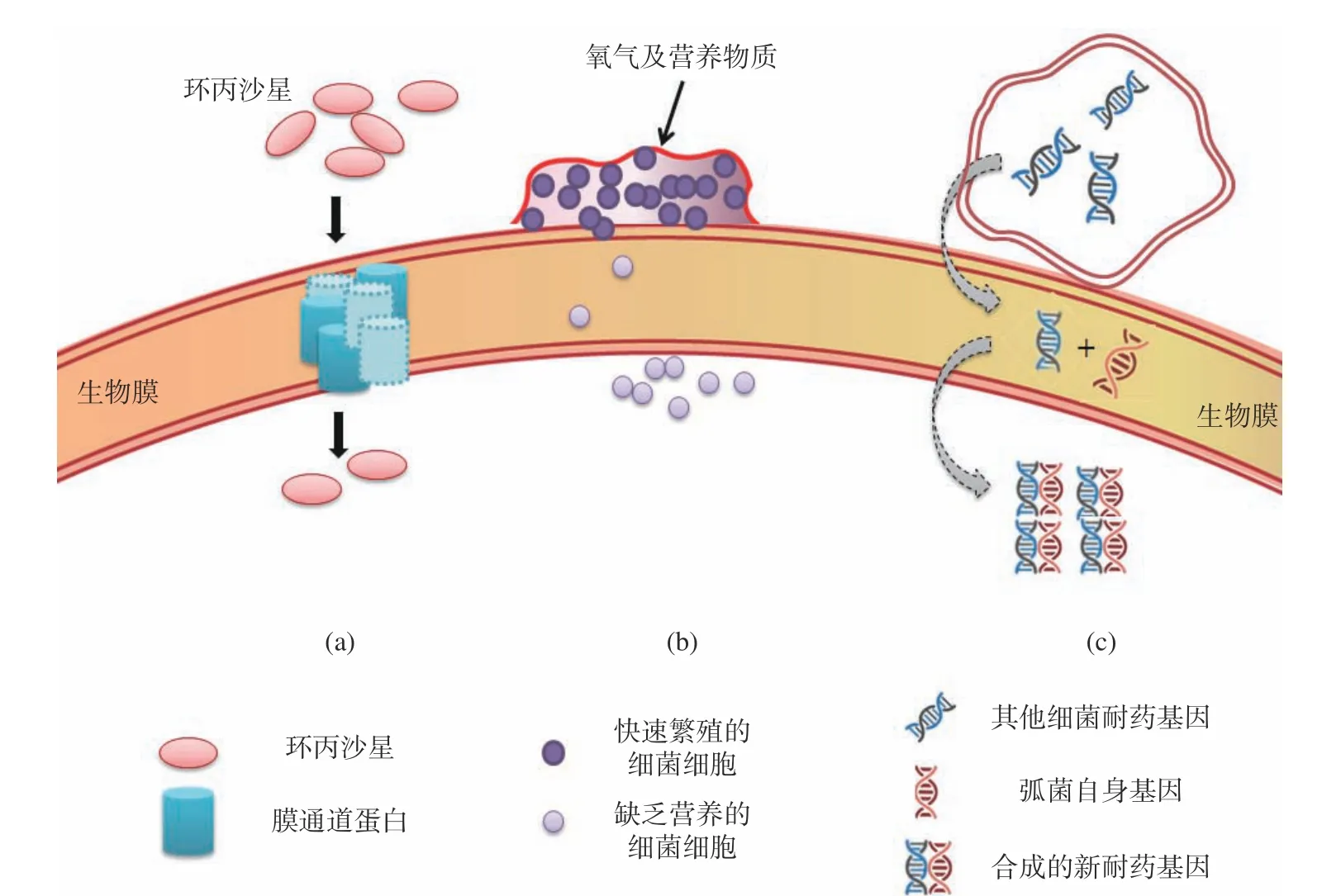
圖3 生物膜對抗生素和殺菌劑的耐藥機制Fig.3 Resistance mechanism of biofilm to antibiotics and fungicides
2.1 生物膜限制抗生素/殺菌劑的滲透
弧菌細胞的多層結構和胞外聚合物(多糖、蛋白、eDNA)構成的復雜而緊密的結構,均可影響抗生素和殺菌劑滲透入胞內。EPS可以通過與殺菌劑進行吸附、靜電或疏水相互作用,限制甚至阻止殺菌劑向細胞的轉運[28]。外膜脂多糖的存在、膜通道蛋白的減少、丟失和替代均可以降低弧菌細胞膜對化合物的滲透性。Das等[29]研究發現,由于霍亂弧菌生物膜功能改變或孔蛋白的缺失,使其對碳青霉烯類、四環素類、氟喹諾酮類、氨基糖苷類和氯霉素產生了耐藥性。Thurnheer等[30]研究表明,在細菌生物膜厚度為150~200 μm時,濃度為25 mg/L的氯滲透深度不超過100 μm。此外,抗生素和生物膜成分之間的潛在相互作用也可能增加生物膜滲透的局限性。如環丙沙星與銅綠假單胞菌生物膜的特定成分結合,使生物膜充當環丙沙星的屏障,使其不能進入細胞,造成細菌耐藥性[31]。Grobe等[32]研究表明,氯、戊二醛和2,2-二溴-3-腈基丙酰胺(DBNPA)與細菌生物膜的成分反應,使殺菌劑濃度減少,對生物膜形成的抑制作用并不明顯,但是這些反應的化學性質仍未確定。
2.2 細菌對亞致死濃度抗生素/殺菌劑的表型適應性
細菌的表型適應是由于細菌對其微環境條件改變而做出的特定基因表達。從細菌的附著到生物膜三維結構的形成,該過程受細菌適應生長環境的能力影響。研究發現,細菌細胞黏附到生物材料表面后,編碼鞭毛蛋白的基因被抑制,而編碼EPS和粘附素蛋白的基因被誘導,促進生物膜的形成,使細菌產生耐藥性[33]。同時,與浮游細胞相比,粘附于表面的細胞具有更大的殺菌劑抗性[34]。位于生物膜外圍的細胞可以獲得營養和氧氣,而生物膜內部層中的細菌則處于營養貧乏的微環境中,這種化學特異性導致細菌生長和代謝速率發生改變,也增加了細菌對殺菌劑的抗性[35]。
2.3 耐藥基因水平傳遞(horizontal gene transfer,HGT)和基因突變
在細菌內部,細菌可通過整合子將多種耐藥基因重組成為多重耐藥菌,而整合子還能以移動基因盒的形式將新基因重組到細菌染色體或質粒等遺傳物質中[36-37]。研究表明,生物膜是遺傳物質交換的最佳環境,膜內細胞間不斷傳遞耐藥基因盒,增加了細菌的耐藥性[38]。同時,高細胞密度的存在、生物膜內大量DNA的釋放或合適的營養條件都可能會促進基因接合和轉化過程,使細胞獲得新的遺傳物質,增強細菌耐藥性。當霍亂弧菌細胞與其他細菌細胞共享移動整合子時,就很容易發生抗生素抗性基因的傳播[39]。此外,靶基因突變可導致細菌細胞壁上的藥物受體減少,進而達到耐藥效果。喹諾酮類藥物是細菌脫氧核糖核酸回旋酶和脫氧核糖核酸拓撲異構酶IV的抑制劑,可抑制DNA復制。在霍亂弧菌中通過使其亞基基因gyrA和parC染色體發生突變,干擾DNA復制和轉錄功能,導致其對抗生素的親和力降低,從而使霍亂弧菌免受喹諾酮類藥物的影響[40]。
從弧菌耐藥機制中可看出,生物膜在細菌產生耐藥性及耐藥基因傳遞中具有重要作用。因此,控制細菌生物膜的形成成為防控細菌產生耐藥性的重要措施。
3 控制弧菌群體感應的群體感應抑制劑
傳統的抗生物膜劑一般以單個細菌為目標,作用于細菌的細胞壁、細胞膜以及DNA等靶點,易導致其產生耐藥性或演變為超級細菌。因此,為減少和控制弧菌產生生物膜及耐藥性,細菌的QS系統成為新靶點,干擾細菌的QS系統成為解決細菌產生生物膜和耐藥性的新途徑[41]。群體感應抑制劑的作用方式主要包括以下三種:a.抑制QS信號分子合成;b.抑制QS分子-受體相互作用;c.降解QS信號分子[42]。目前,一些化學合成類和天然類群體感應抑制劑已被用于抑制弧菌生物膜的形成[43](表1)。

表1 群體感應抑制劑對弧菌QS和生物膜的抑制作用Table 1 Inhibition effect of quorum sensing inhibitors on QS and biofilm of Vibrio sp.
3.1 化學合成類群體感應抑制劑
化學合成類群體感應抑制劑可通過與信號分子競爭性結合受體蛋白或與信號分子合成酶蛋白靶向結合等方式,干擾細菌的群體感應代謝通路,從而抑制生物膜的合成。化學合成類群體感應抑制劑主要包括AHLs類、AI-2類信號分子的合成類似物及其他小分子物質。Kadirvel等[44]研究表明,合成的4-氟DPD是一種AI-2信號分子類似物,可通過干擾AI-2系統來抑制生物膜的形成,且濃度為10-4mol/L時,對哈維氏弧菌生物膜抑制率高達90%。Rajamanikandan等[45]研究發現硼酸衍生物可與AI-2合成酶LuxP靶向對接,當硼酸衍生物質量濃度為4 μg/mL時,對哈維氏弧菌的生物膜形成抑制率達到64.25%,表明硼酸衍生物通過抑制AI-2的合成來抑制哈維氏弧菌QS通路,進而影響生物膜的形成。Soni等[46]研究發現,合成大麻素HU-210可使哈維氏弧菌BB152的luxS、luxP、luxQ和luxR的表達減少,阻礙AI-2的產生以及受體蛋白對QS信號的接收,抑制AI-2介導的QS級聯反應的活性,從而抑制生物膜形成。Kim等[47]研究表明,1-(5-溴噻吩-2-磺酰基)-1氫-吡唑在創傷弧菌中是一種有效的SmcR(LuxR同源物)抑制劑,與SmcR蛋白結合口袋緊密結合,使蛋白質柔韌性發生改變,從而影響SmcR的轉錄表達,阻礙QS調控創傷弧菌的運動性、毒力因子及生物膜形成。
3.2 植物源群體感應抑制劑
植物提取物中的多酚類、醛類、黃酮類、單寧類、萜類和糖苷類等物質都具有良好的抗生物膜活性[48]。研究表明,從香辛料中提取的大麻酚、肉桂醛、呋喃酮和姜黃素能夠有效地抑制弧菌的QS系統,控制生物膜的生成。Aqawi等[49]研究表明,大麻中存在一種少量非精神活性的大麻酚具有QS抑制活性,可阻止哈維氏弧菌LuxU的去磷酸化,從而干擾群體感應自體誘導物信號的傳遞來抑制生物膜的形成;在濃度為20和50 μg/mL的濃度下,生物膜中細菌量分別減少到58.7%和43.8%。Brackman等[50]研究表明,肉桂醛通過降低LuxR與其靶啟動子序列結合的能力而干擾AI-2的QS,從而抑制了鰻弧菌和創傷弧菌生物膜的形成。從柑橘中分離得到的3,5,7,8,3’,4’-六甲氧基黃酮在濃度為5×10-5mol/L時,通過干擾哈維氏弧菌AI-2系統中的LuxO表達,抑制了約61%生物膜的形成[51]。檸檬苦素是柑橘類分泌的一種具有呋喃環的三萜類化合物,通過影響LuxO的表達,顯著抑制哈維氏弧菌生物膜形成的能力[52]。Srinivasan等[53]研究發現胡椒乙酸乙酯提取物(Piper betle ethyl acetate extract, PBE)對哈維氏弧菌生物膜形成的抑制作用,PBE在亞抑菌濃度為400 μg/mL時,對哈維氏弧菌群體感應介導的生物膜形成的抑制作用達到74%。Karnjana等[54]從紅藻的乙醇提取物中分離純化的N-芐基肉桂酰胺和α-間苯二甲酸的化合物可通過干擾哈維氏弧菌AI-2信號通路抑制生物膜的形成。由于植物源提取物的穩定性強、分子量低,且對人體健康無害,其被認為是理想的天然的QS抑制劑。
3.3 微生物源群體感應抑制劑
微生物源的群體感應抑制劑主要來源于酵母、乳桿菌屬(Lactobacillussp.)和芽孢桿菌屬(Bacillussp.)。筆者團隊的研究表明,乳酸乳球菌LY3-1分泌的次級代謝產物肽LQLY3-1可通過競爭性結合信號分子受體LuxN而干擾哈維弧菌QS系統,肽LQLY3-1對哈維弧菌生物膜、溶血素、胞外多糖、蛋白酶等毒力因子的抑制率在21%~66%[55]。此外,團隊研究發現,面包乳桿菌ZHG2-1代謝產物粗提物對銅綠假單胞菌AHLs信號分子的降解率接近100%,在亞最小抑菌濃度(1.0、2.0和3.0 mg/mL)下,對生物膜形成和預先形成生物膜的抑制率和清除率分別為13.40%~47.30%和11.60%~38.70%,同時,抑制細菌蛋白酶、幾丁質酶、綠膿菌素、鼠李糖脂、海藻酸鹽和胞外多糖等毒力因子的產生,使銅綠假單胞菌群體感應調控基因lasI/R和rhlI/R表達下調[56]。上述研究結果說明,乳酸菌來源的淬滅酶、細菌素等代謝產物可通過抑制AHLs合成、競爭性抑制AHLs與受體蛋白結合、AHLs失活等作用方式干擾細菌QS系統。此外,其他研究學者Gutiérrez-Almada等[57]研究發現,濃度為1000 μg/mL地衣芽孢桿菌的乙酸乙酯提取物對C6HSL的降解率為98%,對溶藻弧菌生物膜抑制率幾乎達到100%。微生物源群體感應抑制劑具有易培養、適應性強、無耐藥性等優點,作為一種環境友好的抗生素替代物,可通過干擾弧菌群體感應代謝通路,達到抑制/破壞生物膜的形成和結構的目的。
4 結語與展望
弧菌在水產養殖與海產品生產加工的環境中,可通過不同群體感應信號分子調控基因表達產生毒力因子及成熟的生物膜,進而使弧菌對抗生素/殺菌劑造成了多藥耐藥性。生物膜的形成限制抗菌藥物的滲透,極大地影響殺菌效度,弧菌對亞致死濃度殺菌劑的表型適應性也增加了對抗菌藥物的抗性;同時,耐藥基因水平傳遞和基因突變也給消除弧菌帶來困難。群體感應是控制細菌性食品腐敗和食源性疾病領域的研究熱點之一,通過干擾、抑制、淬滅等多種方式控制群體感應,控制食品腐敗和致病作用是一條有效途徑。因此,基于群體感應減少和控制弧菌產生生物膜及耐藥性,提出一解決細菌產生生物膜和耐藥性的新途徑。本文對弧菌的群體感應作用機制,弧菌生物膜對多藥耐藥性的影響和耐藥機制以及目前新型化學合成類和天然類群體感應抑制劑等進行總結與分析,為水產品養殖與貯藏加工中控制食品腐敗和食源性疾病、挖掘新型綠色健康的群體感應抑制劑方面提供借鑒與參考。
目前,弧菌生物膜的控制技術已經廣泛應用到水產加工與養殖領域中,但還存在許多問題。一方面,化學合成類和天然類群體感應抑制劑的合成與添加過程中,殘留的有毒化學物質對人類健康有潛在危害,且商用群體感應抑制劑價格昂貴。另一方面,在食品加工環境中,有多種不同的微生物附著在食品或器械表面,其中群體感應種間交流AI-2系統可以促進弧菌與其形成耐藥性更強更難除去的混合生物膜。因此,研究各環境條件下混合菌的生物膜控制技術,開發價格低廉、綠色、健康的群體感應抑制劑是未來將要攻克的難題。主要可以從以下幾個方面開展:a.QS抑制劑可以與抗生素或其它活性物質,如抗菌肽、噬菌體等聯合應用,旨在降低細菌耐藥性、減少混合生物膜形成及其對人類健康的危害。b.研究者們對生物膜形成及控制大部分都停留在對其結構構象表層的研究,對其作用機理機制并未進行深層探索,因此可以通過轉錄組學或基因敲除等方式,研究QSI對生物膜相關基因的調控作用,進一步揭示其機理。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
鴨綠江(2021年35期)2021-04-19 12:24:18
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
家庭影院技術(2017年9期)2017-09-26 03:41:45
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
