基于水稻矮稈長粒CSSL-Z688的QTL鑒定及SSSLs培育
2023-01-16 03:18:26李苗苗李儒香秦魚河鄧岢莉余金琎徐光益向思茜楊正林桑賢春凌英華何光華趙芳明
西南大學學報(自然科學版) 2023年1期
李苗苗, 李儒香, 秦魚河, 鄧岢莉,余金琎, 徐光益, 向思茜, 楊正林,桑賢春, 凌英華, 何光華, 趙芳明
1. 西南大學 水稻研究所/農業科學研究院,重慶 400715; 2. 瀘州泰豐種業有限公司,四川 瀘州 646009
水稻是重要的糧食作物,提高水稻產量是每個育種家的不懈追求[1].然而,水稻產量的形成是復雜的,往往與株高、有效穗數、每穗粒數和千粒質量等密切相關.株高決定水稻生物量,植株過高易引起倒伏,從而影響光合效率.因此,合適的水稻植株高度對提高產量至關重要.同時,細胞擴張會影響粒型,粒型影響千粒質量,也是產量的決定性因素之一.然而,這些性狀均由多個基因控制,遺傳機制非常復雜,屬于典型的數量性狀[2-5].隨著分子標記技術的快速發展,研究人員通過構建不同的遺傳群體對水稻農藝性狀進行了QTL定位分析[6].利用傳統分離群體(如F2,F2:3,重組自交系和雙單倍體系進行株高和粒型的QTL定位,鄭躍濱等[7]以短粒普通野生稻突變體和長粒栽培稻品種 KJ01構建的 F2分離群體,檢測到 24個控制水稻粒型的 QTLs ; Zhou等[8]以廣占63-4S和TGMS29為親本構建的F2及其衍生的F2:3群體共檢測到36個粒型QTLs; Liu等[9]利用兩個秈稻品種 Taifeng B和Tesanai 2為親本構建的170個家系的重組自交系(Recombinant Inbred Lines,RILs)群體共檢測到 34個控制水稻粒型的 QTLs; 封功能等[10]利用秈粳交組合Nanjing 11號×Balilla創建的DH群體,鑒定出50個與產量相關性狀的QTLs.然而,由于初級分離群體中個體間遺傳背景的干擾常使QTL定位不準確,也較難直接與育種應用相結合,一定程度上限制了這些QTL的進一步克隆和育種.為了提高QTL的定位準確度,且實現QTL定位和設計育種的一體化,染色體片段代換系(Chromosome segment substitution lines,CSSL)、近等基因系(NIL)等次級作圖群體逐漸受到研究者的青睞.水稻染色體片段代換系是創造自然變異的理想材料,同時又可將多位點控制的復雜性狀進行遺傳分解,尤其定位出的QTL可直接應用于育種實踐,因而是理想的遺傳研究材料[11].姚國新等[12]利用大粒品種SLG-1與小粒日本晴雜交、回交構建的一套關于千粒質量和粒型的姊妹近等基因系(Sister Near-isogenic Lines,SNILs)群體,并檢測到12個QTLs; 游佳等[13]以‘9311’為受體親本,普通野生稻為供體親本創建的染色體片段代換系群體共定位到水稻千粒質量、粒長、粒寬和長寬比等性狀的16個QTLs; Fan等[14]以HHZ為受體親本,BAS為供體親本構建的染色體片段代換系群體,檢測到25個QTLs.本課題組也分別以日本晴和西恢18為受體親本創建了兩套水稻染色體片段代換系,并進行了一些產量相關性狀的QTL定位和QTL聚合分析,如Liang等[15]以西恢18為受體親本和滬旱3號為供體親本構建的7代換片段CSSL-Z563為材料鑒定出11個水稻粒型QTLs; Sun等[16]以西恢18為受體親本和滬旱3號為供體親本構建的6代換片段水稻CSSL-Z431為材料鑒定出13個水稻穗數和粒型QTLs; Wang等[3,17]以日本晴為受體親本和西恢18為供體親本構建的2個CSSL-Z747和CSSL-Z749為材料分別鑒定出46個和15個水稻產量相關性狀QTLs; Ma等[18-19]以日本晴為受體親本和西恢18為供體親本構建的3代換片段CSSL-Z1364和6代換片段CSSL-Z744為材料分別鑒定出8個和17個水稻產量相關性狀QTLs; Wang等[20]以日本晴為受體親本和西恢18為供體親本構建的6代換片段CSSL-Z741為材料鑒定出20個水稻重要農藝性狀QTLs; Zhang等[21]以日本晴為受體親本和西恢18為供體親本構建的3代換片段CSSL-Z741為材料鑒定出7個水稻粒型QTLs.由于這些CSSL所含的代換片段各不相同,且檢出不同性狀的QTL,為以CSSL為平臺的水稻設計育種計劃奠定了良好的基礎.
盡管定位了大量水稻產量相關性狀的QTL,但由于水稻株高、粒型等性狀受多個QTL控制,要實現全基因組水稻分子設計育種,有必要鑒定出更多的有利QTL,且將這些QTL分解到單個單片段代換系(Single segment substitution lines,SSSL)中,以便更精準地研究單基因功能和實現設計育種操作.因此,本研究在6代換片段的染色體片段代換系Z744重要農藝性狀QTL定位的基礎上[19],以受體日本晴與Z744雜交后代中進一步選育的矮稈、長粒水稻染色體片段代換系Z688為研究材料,進行水稻株高、粒型性狀的QTL定位,并進一步培育目標QTL的純合和雜合單片段代換系,研究來自供體西恢18等位基因的加性和顯性效應,為設計育種提供重要遺傳信息.
1 材料與方法
1.1 試驗材料
本研究使用的材料是4代換片段的水稻矮稈長大粒染色體片段代換系Z688.Z688在Ma等[19]培育的6代換片段代換系Z744(平均代換長度為2.72 Mb)的QTL定位基礎上選育而成的.Z744以日本晴為受體親本,西恢18為供體親本,經高代回交和自交,以水稻全基因組選出二者間有多態性的263個SSR標記[22],從BC2F1開始分子標記輔助選擇(Molecular marker-assisted selection,MAS),每代每個株系選20株,繼續MAS選擇,在日本晴/BC2F4的F6代選出了6代換片段的水稻純合染色體片段代換系Z744[19].本研究從受體日本晴與Z744雜交的F2:3群體(經QTL初定位[19])進一步MAS選育出1個純合4代換片段的矮稈長粒染色體片段代換系Z688.選擇過程中,與受體日本晴帶型一致時標“A”,與供體西恢18的帶型一致時標“B”,雜合帶型標“H”.Z688的代換片段鑒定參照Liang等[15]描述的方法,估計代換片段長度計算參照Paterson等[23]的方法進行.
1.2 植物材料與田間種植
2019年在西南大學水稻基地(重慶市歇馬鎮),以日本晴與Z688雜交,收取雜交種并于同年在海南基地種植F1,并收取F1種子.2020年3月8日,在西南大學水稻基地育苗,4月15日,以株距和行距分別為16.67 cm和26.67 cm移栽日本晴和Z688各30株,用于QTL定位的150個單株組成的F2群體于同一試驗田,按常規模式進行田間管理.2021年 3月10日,在西南大學水稻基地對日本晴,Z688和從F2群體選出的10個單株進行育苗,于4月18日,將日本晴,Z688和10個株系(Z1038-Z1047)各移栽30株,按常規方式進行管理.
1.3 Z688的株高、粒型性狀評價
成熟后,平地面收取日本晴和Z688各10株以及150株F2群體.考察株高、10粒長、10粒寬、千粒質量,然后用粒長除以粒寬計算長寬比.具體參照Wang等[20]描述的方法測定,株高用5 m卷尺測定從地面到每株最長穗頂端的距離.每株選30粒飽滿一致的籽粒,隨機將10粒分別按長和寬首尾相連,然后用20 cm直尺測量其總長度和總寬度,重復3次,平均數計為10粒長度和10粒寬度,然后換算為每粒長和寬.谷粒長寬比以粒長與粒寬的比值計算.對于日本晴和Z688的千粒質量,隨機選取3 000粒,然后用電子天平每1 000粒為1組進行測量,重復測定3次.對于QTL定位群體的千粒質量測量,則每株隨機取200粒,用電子天平測量,重復3次,然后乘以5計算.最后,使用Microsoft Excel 2016統計10株日本晴和Z688及F2群體每株性狀的平均值和標準差,并進行t測驗.
1.4 QTL定位
以日本晴與Z688雜交構建的由150個單株組成的次級F2群體作為QTL定位群體,用十六烷基三甲基溴化銨法(CTAB)法提取親本和150個F2單株的DNA,參照Zhao等[24]描述的方法進行PCR擴增和非變性聚丙烯酰胺凝膠電泳.將日本晴帶型、Z688帶型、雙親帶型、缺失帶型基因型賦值,分別用“-1”“1”“0”和“.”來表示.結合150個F2單株對應的表型值,使用SAS9.3統計軟件(SAS Institute Inc,Cary,NC,USA)的混合線性模型(Mixed linear model,MLM)法進行QTL定位[24],以p<0.05為閾值,決定QTL是否存在.
1.5 目標QTL的純合和雜合單片段代換系選育及QTL加性和顯性效應分析
根據2020年QTL定位信息,從F2選擇10個含目標QTL的單株(僅含純合或雜合目標代換標記,其余標記帶型與受體日本晴一致),2021年種成株系(Z1038-Z1045),每個株系取30株的葉片提取DNA,利用 MAS對其中的目標代換標記進行進一步跟蹤篩選,選育出純合的SSSL和雜合的SSSL(H).于2021年7月下旬,收獲10株日本晴和選育出的SSSL及SSSL(H)的所有單株(8~10株),按相同的方法測量株高、粒長、粒寬、長寬比和千粒質量,用于純合目標QTL的加性效應和雜合QTL的顯性效應分析.
1.5.1 基于4個純合SSSLs的QTL加性效應分析
對于純合SSSL(S1-S4),首先假設H0:SSSLi的代換片段不存在控制某一性狀的QTL,然后對每個SSSL和受體日本晴性狀進行t測驗,若p<0.05時,表明SSSLi的代換片段存在控制某一性狀的QTL.根據在特定環境條件下,日本晴的遺傳模型為
p0=μ+ε
和單片段代換系的遺傳模型為
pi=μ+ai+ε
式中,μ為日本晴某一性狀的表型值,ai為QTL的加性效應,pi和p0分別表示SSSL和日本晴的表型值,ε為隨機誤差[15].
我們可估計出該QTL的加性效應(ai)為
ai= (pi-p0)/2
表型差一半被估算為由遺傳引起.
所有計算均在Microsoft Excel 2016中進行.
1.5.2 基于4個雜合SSSL(H)的QTL顯性效應分析
對于雜合SSSL(H),首先假設H0:呈雜合狀態時的SSSLi(H)不存在顯性效應d,即
pi(H)=(p0+pi)/2
式中,pi(H),p0和pi分別為SSSLi(H),日本晴和SSSLi的表型值,然后對pi(H)和(p0+pi)/2做t測驗,若p<0.05,則說明H0不成立,即該代換片段所含控制某一性狀的QTL存在顯性效應d.根據顯性效應原理,雜合SSSLi(H)的顯性效應(di)為
di=pi(H)-(pi+p0)/2
所有計算均在Microsoft Excel 2016中進行.
2 結果
2.1 Z688的代換片段和差異性狀統計分析
Z688含有來自西恢18的4個染色體代換片段,分布于水稻第1,2,6和12染色體上(圖1),其總代換片段的估計長度為8.53 Mb,最長代換片段的估計長度為5.06 Mb,最短為0.55 Mb,平均為2.13 Mb.

每條染色體左側為物理距離(Mb)和定位的QTL,右側為標記名稱和染色體代換片段長度; 框內標記為代換區間.PH為株高,GL為粒長,RLW為谷粒長寬比,GWT為千粒質量.圖1 CSSL-Z688的代換片段及攜帶的QTL
Z688的株高(65.29 cm)顯著矮于受體日本晴(104.11cm),比日本晴降低了38.82 cm(圖2a,d); Z688的長寬比(2.33)比日本晴(2.18)顯著增加了0.15(圖2c,e); Z688的粒長(7.80 mm)比受體日本晴(7.28 mm)顯著增加了0.52 mm(圖2c,f); Z688的千粒質量(24.16 g)比日本晴(21.94 g)顯著增加了2.22 g(圖2g).
Z688的有效穗數、穗長和粒寬等性狀與日本晴差異無統計學意義(統計數據略)(圖2a,b).
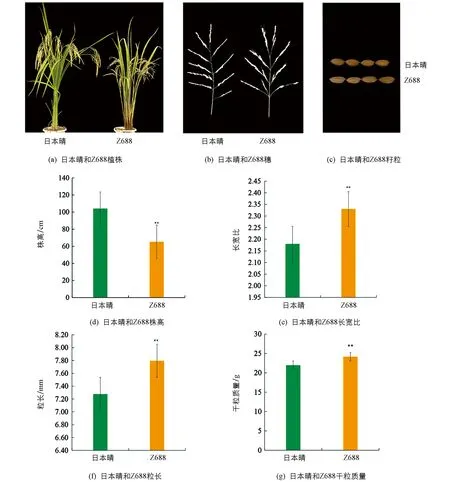
*表示p<0.05,**表示p<0.01,差異有統計學意義.圖2 受體日本晴和CSSL-Z688的表型和差異性狀
2.2 Z688代換片段攜帶的株高和粒型QTL
以日本晴和Z688雜交構建的次級F2群體共鑒定出7個控制Z688株高與粒型性狀的QTLs,分布于第1,第6和第12染色體的代換片段上,解釋了從0.84%到23.94%的表型變異(表1).來自西恢18的主效qPH1-1和微效qPH1-2與Z688的矮稈性狀連鎖,均位于第1染色體的代換片段上,連鎖于不同的分子標記,其加性效應分別使株高減少了3.20 cm和2.16 cm,分別解釋了15.00%和0.84%的株高變異.來自西恢18的2個主效qGL1和qGL6及微效qGL12與Z688的粒長性狀連鎖,其加性效應分別使Z688的粒長增加了0.11 mm,0.09 mm和0.06 mm,分別解釋了23.94%,15.36%,8.83%的粒長變異.來自西恢18主效qGWT1影響Z688的大粒,其加性效應使千粒質量增加了0.66 g,解釋了9.76%的表型變異.此外,qPH1-1,qRLW1,qGL1和qGWT1連鎖于同一標記RM128.
在qPH1-1的代換區間內有已克隆基因OFP3和OsHXK6,在qPH1-2的代換區間內有已克隆基因OsERF3和PSD1,在qGWT1的代換區間內有已克隆基因OsMKKK55,OsMKKK70和OsMKKK62(表1).
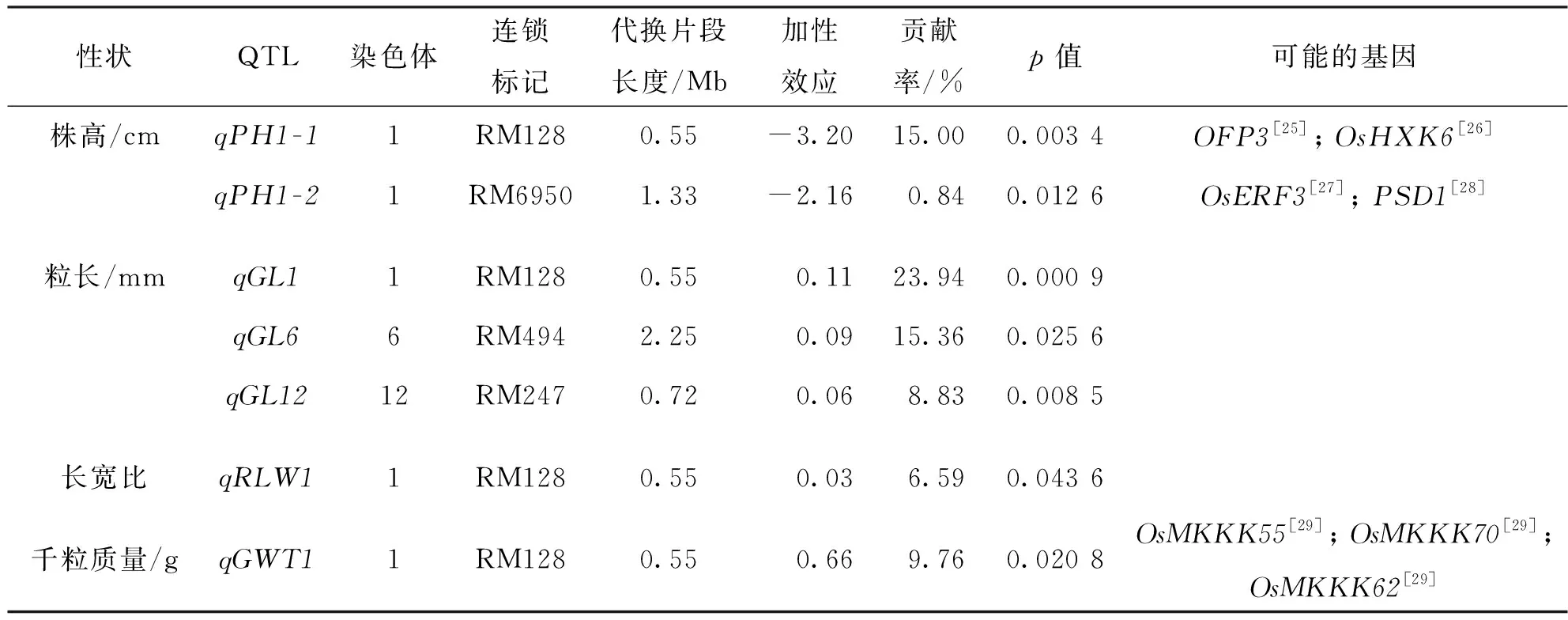
表1 Z688代換片段攜帶的水稻株高和粒型性狀QTL
2.3 目標QTL的次級單片段代換系(SSSLs)的培育及加性和顯性遺傳效應分析
基于QTL定位,通過MAS在F3群體選育出5個純合單片段代換系SSSLs(S1-S5)和5個對應的雜合單片段代換系(S1(H)-S5(H)).S1的代換片段為RM403-RM128-RM1268,位于第1染色體上,估計代換長度為0.67 Mb; S2的代換片段為RM128-RM6950-RM5389,位于第1染色體,估計代換長度為1.82 Mb; S3的代換片段為RM3294-RM6378-RM2483,位于第2染色體上,估計代換長度為0.55 Mb; S4的代換片段為RM400-RM7412-RM439-RM494-RM20769,位于第6染色體上,估計代換長度為2.20 Mb; S5的代換片段為RM6288-RM247-RM491,位于第12染色體上,估計代換長度為0.72 Mb(表2).
在2020年定位的7個QTLs中,有6個(qPH1-1,qPH1-2,qGL1,qGL12,qRLW1和qGWT1)可被相應的純合單片段代換系(S1-S5)在2021年驗證.此外,在這5個純合單片段代換系中還新檢測到11個QTLs,分別為qPH2,qGL1-2,qGL2,qGW1,qRLW1-2,qRLW2,qRLW12,qGWT1-2,qGWT2,qGWT6和qGWT12.這些結果表明,由于單片段代換系消除了個體間遺傳背景的干擾,SSSL具有比F2群體更高的QTL檢測效率(表2).
我們還利用5個雜合單片段代換系(S1(H)-S5(H))對QTL的顯性效應進行了分析,結果表明,雜合位點QTL的顯性效應也對表型有較大的影響(表2).
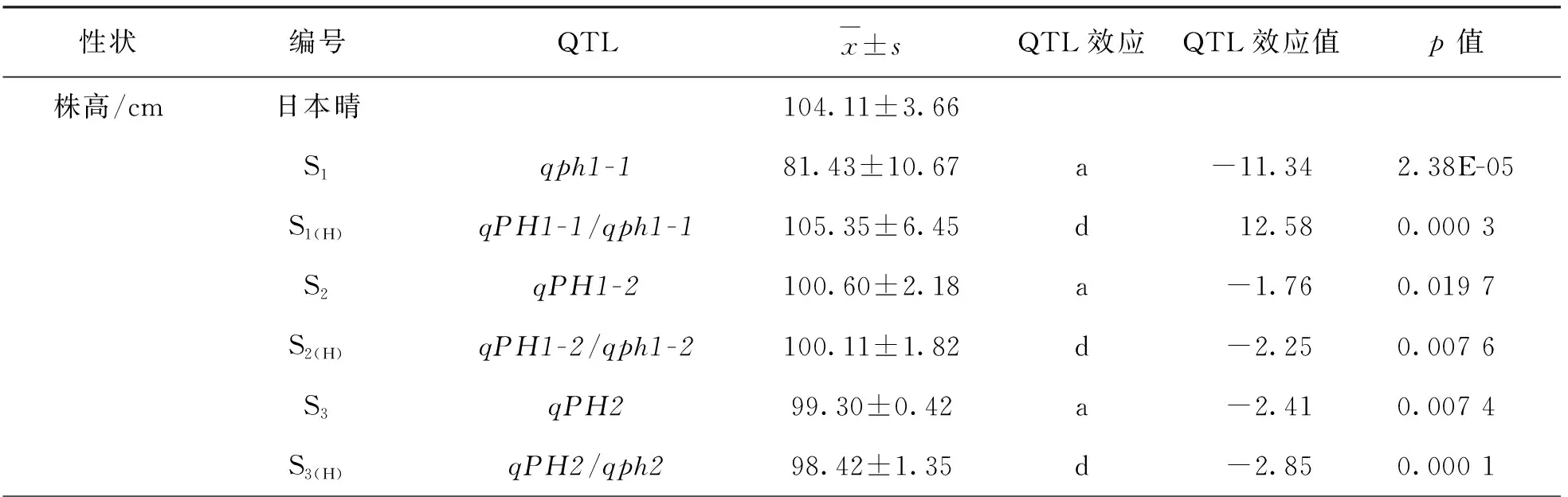
表2 基于5個SSSLi和SSSLi(H)的水稻株高及粒型相關性狀QTL加性和顯型效應
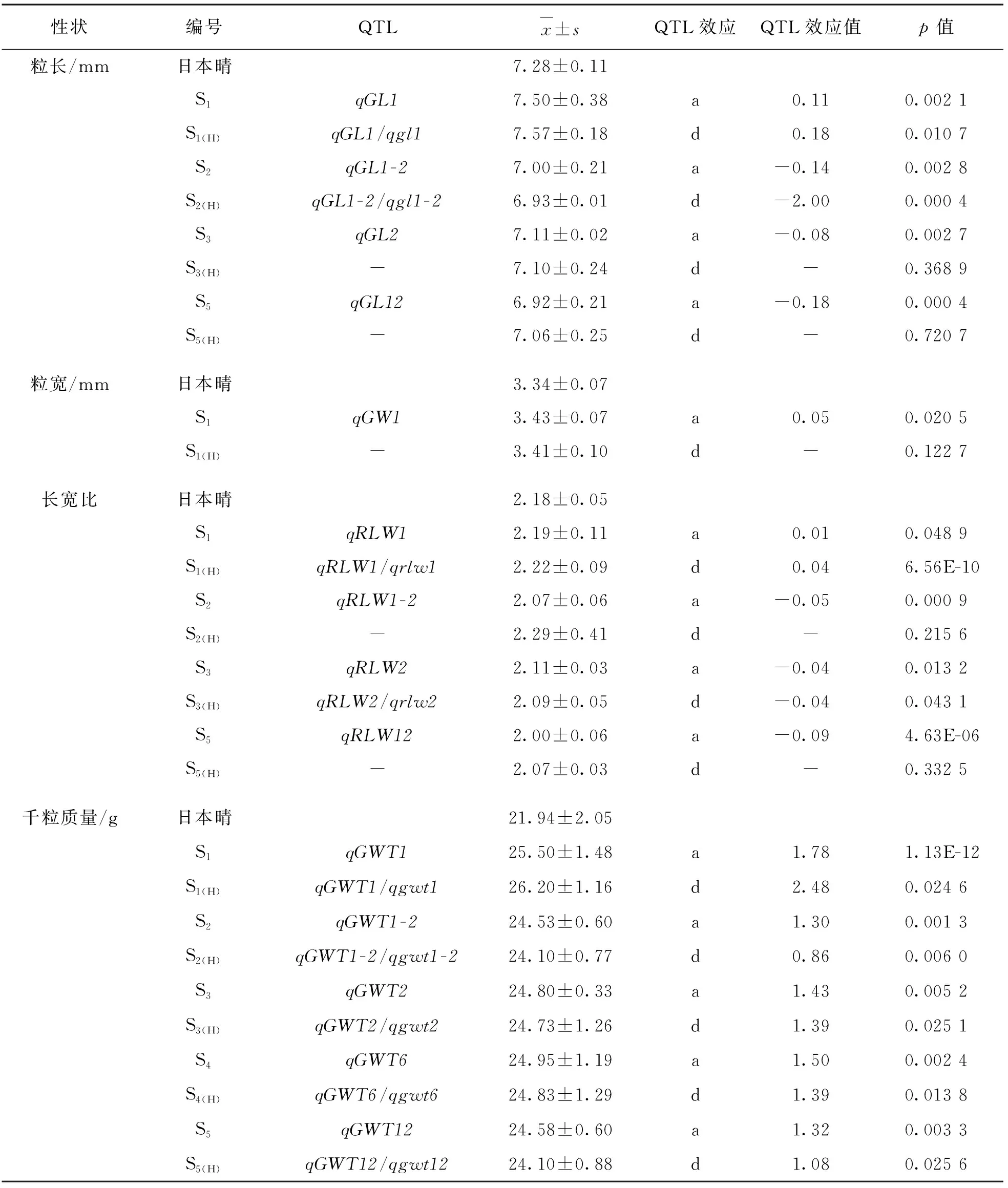
續表2
利用純合的單片段代換系和雜合的單片段代換系可反映出純合位點加性效應和雜合位點顯性效應同時影響株高的遺傳.位于第1染色體的來自西恢18的qph1-1,qPH1-2和位于第2染色體的qPH2的加性效應分別使株高降低11.34 cm,1.76 cm和2.41 cm,qPH1-1/qph1-1的顯性效應使株高增加12.58 cm,暗示來自西恢18的株高qph1-1表現為相對隱性,而來自日本晴的qPH1-1則相對顯性.qPH1-2/qph1-2和qPH2/qph2的顯性效應分別使株高降低2.25 cm和2.85 cm,表明來自西恢18的株高qPH1-2和qPH2等位基因表現為相對顯性,而來自日本晴的qph1-2和qph2相對隱性(表2).這些結果表明,株高的表型是這些QTL的加性效應和顯性效應的疊加效應,揭示了遺傳復雜的數量性狀可通過單片段代換系的構建分解為單位點進行研究.
對于粒長性狀,來自西恢18的qGL1的加性效應在遺傳上使S1的粒長增加0.11 mm,而來自西恢18的qGL1-2,qGL2和qGL12的加性效應使粒長分別減少了0.14 mm,0.08 mm和0.18 mm,qGL1/qgl1的顯性效應使粒長增加了0.18 mm,表明來自西恢18 的qGL1相對來自日本晴的qgl1表現顯性.對于減少粒長的qGL1-2,qGL2和qGL12對應的雜合位點沒有檢出顯性效應(表2).同樣表明,粒長是由多基因控制的數量性狀,其長粒表型是多個QTL加性和顯性效應的疊加.
來自西恢18的qGW1的加性效應使S1的粒寬增加0.05 mm,而在相應的雜合位點無顯性效應(表2),Z688的粒寬也與日本晴的差異無統計學意義.
對于水稻谷粒長寬比,來自西恢18的qRLW1的加性效應使長寬比增加0.01,而來自西恢18的qRLW1-2,qRLW2和qRLW12的加性效應分別使谷粒長寬比減少0.05,0.04和0.09.qRLW1/qrlw1的顯性效應使谷粒長寬比增加0.04,表明來自西恢18的增加谷粒長寬比的qRLW1相對來自日本晴的qrlw1表現顯性.qRLW2/qrlw2的顯性效應使谷粒長寬比減少0.04,表明來自西恢18的減少谷粒長寬比的qRLW2對日本晴的qrlw2也表現相對顯性.另外,在其他雜合位點并未檢出顯性效應(表2).
對于千粒質量,來自西恢18的qGWT1,qGWT1-2,qGWT2,qGWT6和qGWT12的加性效應分別在遺傳上使千粒質量增加1.78 g,1.30 g,1.43 g,1.50 g和1.32 g,qGWT1/qgwt1,qGWT1-2/qgwt1-2,qGWT2/qgwt2,qGWT6/qgwt6和qGWT12/qgwt12的顯性效應分別使千粒質量增加2.48 g,0.86 g,1.39 g,1.39 g和1.08 g(表2),表明來自西恢18的增加千粒質量的qGWT1,qGWT1-2,qGWT2,qGWT6和qGWT12相對日本晴的相應等位基因表現顯性.這些結果進一步驗證了千粒質量是由微效多基因控制的數量性狀,但可通過構建單片段代換系加以分解研究.
3 討論
水稻產量主要由3要素構成:有效穗數、每穗實粒數和千粒質量[30].千粒質量主要由粒長、粒寬和粒厚決定.此外,水稻株高與產量的形成密切相關[31].本研究選育了1個以日本晴基因組為遺傳背景含來自西恢18基因組的4代換片段的水稻CSSL-Z688,與日本晴相比,Z688的籽粒顯著變長,千粒質量顯著增加,其千粒質量的顯著增加主要由籽粒長度變長引起,與Ma等[19]鑒定的水稻染色體片段代換系Z744的長粒表型相似,但Z688的水稻株高顯著變矮.矮稈突變的發現曾引起第一次綠色革命,矮稈基因是寶貴的遺傳資源.選育高度適中的水稻品種,不易倒伏,灌漿良好,是水稻理想的高產株型[32].因此,Z688為研究株高和粒型基因提供了良好的遺傳資源.水稻染色體片段代換系,與受體親本相比,僅存在少量代換片段的差異,因而減少了遺傳背景的干擾,是QTL準確定位的良好材料[24].通過染色體片段代換系的構建可將由多基因控制的數量性狀進行分解,進而挖掘出更多的有利等位變異,為目標基因的遺傳解析和水稻分子聚合育種奠定良好的基礎[33-34].
經日本晴/Z688構建的次級F2群體的QTL定位,發現7個控制株高和粒型性狀的QTLs,其中2個(qPH1-1和qPH1-2)使株高變矮,3個(qGL1,qGL6和qGL12)使粒長增加,1個(qRLW1)使谷粒長寬比增加和1個(qGWT1)使千粒質量增加.此外,我們進一步分離出5個純合單片段代換系(S1-S5)和5個雜合單片段代換系(S1(H)-S5(H)).在5個純合單片段代換系中,6個QTLs能夠被驗證,此外還新檢出了11個QTLs.與前人研究結果相比,發現編碼卵形家族蛋白的OFP3[25]與編碼己糖激酶的OsHXK6[26]與株高qPH1-1的連鎖標記RM128分別相距0.16 Mb和0.29 Mb; 編碼乙烯應答轉錄因子的OsERF3及編碼光周期敏感的矮稈基因PSD1與株高qPH1-2的連鎖標記RM6950分別相距0.84 Mb和0.63 Mb[27-28]; 編碼絲裂原活化蛋白激酶的OsMKKK55,OsMKKK70和OsMKKK62與千粒質量qGWT1的連鎖標記RM128分別相距1.80 Mb,1.81 Mb和1.82 Mb[29].qGL1和qGWT1也被Ma等[19]利用日本晴為受體的水稻6代換片段CSSL-Z744 鑒定出.同時,也與李鋒等[35]定位的qGL1-1,江靜[36]定位的qGL1-1位于相近區間,兩者qGL1-1可分別解釋1.80%,16.2%的表型變異[35-36].而在本研究中qGL1對粒長的貢獻率達到23.94%,表明qGL1可在多年多點、多個遺傳背景下被檢出,是遺傳穩定的QTL,而且當遺傳背景純化后,其加性效應會更大.qGL6還能被沈文強等[37]、Zhang等[21]用不同的水稻染色體片段代換系檢測到.qGL12與王大川等[3]、丁膺賓等[38]定位的QTL區間相近,表明這些QTL遺傳穩定,在基因設計育種中有重要的價值.qGW1,qGL1,qGL6,qGL12,qRLW1,qRLW1-2和qRLW12還尚未見報道,可能是新鑒定的QTL,這些研究結果為后續功能研究奠定了良好基礎.
加性效應和顯性效應是水稻性狀的重要遺傳組成,了解目的基因的加性和顯性效應對作物設計育種有重要意義.本研究結果表明,其株高、粒型等性狀的遺傳受到多個QTLs加性和顯性效應的共同影響,是其加性和顯性效應的疊加.如株高qph1-1,qPH1-2,qPH2的加性效應分別使株高降低11.34 cm,1.76 cm和2.41 cm.qPH1-1/qph1-1的顯性效應使株高增加12.58 cm,而qPH1-2/qph1-2和qPH2/qph2的顯性效應分別使株高降低2.25 cm和2.85 cm.來自西恢18的qGL1的加性效應在遺傳上使粒長增加0.11 mm,而來自西恢18的qGL1-2,qGL2和qGL12的加性效應使粒長分別減少了0.14 mm,0.08 mm和0.18 mm.qGL1/qgl1的顯性效應使粒長增加了0.18 mm.這些結果表明,遺傳復雜的數量性狀可通過單片段代換系的構建分解為單位點進行研究,而且通過多個單基因的聚合可揭示復雜性狀的遺傳組成,并預測聚合基因型的表型.Liang等[15]研究表明,在西恢18的遺傳背景下,聚合加性效應為-0.86 mm的粒長qGL3-2與加性效應為-0.82 mm的粒長qGL3-1產生了-1.02 mm的加性×加性上位性互作效應,最終使該雙基因聚合系產生了比單個單片段代換系更短的籽粒.Zhang等[21]在日本晴遺傳背景下,加性效應為0.30 mm的粒長qGL5和加性效應為0.13 mm的粒長qGL6聚合產生了0.31 mm的加性×加性上位性互作效應,使雙位點聚合系D2的粒長顯著長于僅含單個粒長QTL的單片段代換系S2和S3.Liang等[15]認為以單片段代換系為平臺的多基因聚合系的表現依賴于加性效應與上位性互作效應的代數和與單個QTL加性效應的比較,因此了解單片段代換系QTL的加性效應及其上位性互作效應對植物設計育種有重要意義.此外,顯性效應及顯性×顯性上位性互作效應是雜種優勢利用的遺傳基礎,Zhang 等[21]研究表明,雜種優勢包括單座位的超顯性效應、顯性互補和上位性互作.本研究首次利用雜合單片段代換系分析了水稻株高、粒長、粒寬、千粒質量等性狀單座位的顯性效應,各個性狀都呈現出較大的顯性效應,如qGWT1/qgwt1,qGWT1-2/qgwt1-2,qGWT2/qgwt2,qGWT6/qgwt6和qGWT12/qgwt12的顯性效應分別使千粒質量增加2.48 g,0.86 g ,1.39 g,1.39 g和1.08 g,因此本研究鑒定出的重要QTL的加性和顯性效應,為以后利用這些QTLs進行設計育種提供了可靠的遺傳信息.
4 結論
本研究以受體日本晴和一個4代換片段的水稻矮稈長大粒染色體代換系Z688雜交構建的次級F2群體為QTL作圖群體,鑒定了7個控制株高、粒長、長寬比和千粒質量的QTLs,并進一步培育了目標QTL的5個純合單片段代換系和5個雜合單片段代換系.其中6個QTLs能夠被純合單片段代換系所驗證,此外還在單片段代換系上新鑒定出11個QTLs,分別為qPH2,qGL1-2,qGL2,qGW1,qRLW1-2,qRLW2,qRLW12,qGWT1-2,qGWT2,qGWT6和qGWT12,其中qGW1等7個QTLs尚未被報道.QTL加性和顯性效應表明,株高、粒型、千粒質量等性狀的遺傳受到多個QTL的加性和顯性效應的共同影響,暗示了遺傳復雜的數量性狀可通過單片段代換系的構建分解為單基因進行研究.這些結果為以單片段代換系為平臺的水稻分子設計育種提供了重要的遺傳信息.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中學物理·高中(2016年12期)2017-04-22 11:53:03
