水稻光溫敏雄性不育突變體tms3650的鑒定和基因定位
2023-01-18 12:32:56唐杰龍湍吳春瑜李新鵬曾翔吳永忠黃培勁
中國水稻科學 2023年1期
關鍵詞:水稻
唐杰 龍湍 吳春瑜 李新鵬 曾翔 吳永忠 黃培勁
(1海南波蓮水稻基因科技有限公司, 海口 510125; 2海南大學 熱帶作物學院, 海口 510125; *通信聯系人, email: jesontom@126.com)
水稻(Oryza sativaL.)是雌雄同花自花授粉作物。雄性不育(male sterility,MS)是指雌性器官發育正常,而雄性器官由于發育退化或不能產生有功能的花粉或雄配子從而失去生殖功能導致不育的特性[1],它是水稻的一種生理現象,也是作物雜種優勢利用的基礎,在自然界中發生的概率僅為0.13%[2]。早在1876年,達爾文就提出作物雜交后代部分農藝性狀有超越親本的表現[3]。1926年,美國農學家Jones提出水稻也具有雜種優勢[4]。1973年,石明松[5]在晚粳農墾58中發現了3株光溫敏雄性不育株,隨后育成了農墾58S[6],這一新成果為雜交水稻從“三系法”過渡到“兩系法”開拓了新局面。粳稻品種農墾58S和秈稻品種安農S-1[7]、株1S[8]是研究和應用最廣泛的光溫敏雄性不育系,目前生產上應用的“兩系不育系”大部分都是這三個品種的衍生系[9-10]。農業部統計資料顯示,截至2012年,兩系法雜交稻已累計種植3.2×107hm2(4.99億畝)[11]。
“兩系法”的核心是光溫敏細胞核雄性不育系(photoperiod-/thermo-sensitive genic male sterile line),其育性受到溫度和光照長度的影響,具體表現為短日照(或低溫)條件下,雄性正常可育;而長日照(或高溫)條件下,雄性敗育[12]。光溫敏不育系受細胞核基因調控,隨著分子生物學與分子標記技術的發展,截至2021年底,查詢到定位或克隆的光敏雄性不育基因(PGMS)有pms1[13]、pms2[13]、pms3(p/tms12-1)[14-16]、pms4[17]、RMS1[18]、OSWYOXIB[19]和CSA[20]。溫敏雄性不育基因(TGMS)在不同品種中差異很大,遺傳較為復雜,可能不是簡單的遺傳性狀調控[21]。溫敏雄性不育基因有tms1[22]、tms2(ORMDL)[23-24]、tms3[25]、tms4[26]、TGMS[27]、tms5(ptgms2-1(t))[28-29]、tms6[30]、tms6(t)-1[31]、tms6(t)-2[32]、tms8[33]、tms9[34]、tms9-1(PTC1)[35-36]、tms10[37]、tms18(osNP1)[38-39]、TMS[40]、Ms-h[41]、rtms-1[42]和OsUgp1[43],以及光溫敏不育基因(PTGMS)pms1(t)[44]等。pms1和pms3(Os12g0545900)都是非編碼lncRNAs,但pms1為不完全顯性基因且在長日照條件下的PSMS株系中,phasiRNAs偏好性積累,從而造成光敏雄性不育[13];pms3[14-16]為隱性基因,在長日照下特異的RNA分子轉錄量降低,結果造成正處于發育的花藥進入程序化死亡(PCD)從而造成雄性不育。RMS1(Os04g0461000)和CSA(Os01g0274800)都編碼MYB轉錄因子,李莉等[18]通過CRISPR/Cas9敲除RMS1后得到了具有光敏型雄性不育表型的突變體;而CSA是一個反光敏雄性不育基因,在雄性生殖發育過程中調控糖分配并且是種子早期萌發過程中的一種保護機制[20]。OSWYOXIB(Os02g0816900)編碼一個肌球蛋白,可能在花藥細胞的營養運輸中起著重要的作用[19]。tms2(Os07g0452500)是一種血清類黏蛋白(ORM),植物ORMDL蛋白影響了鞘脂平衡,敲除后因為花粉發育異常而影響育性[24]。tms5(Os02g0214300)編碼一個保守的RNA酶,在tms5突變體中,由于RNA酶功能缺失導致UbL40mRNA水平上升,mRNA過度積累導致花粉形成缺陷和雄性不育[45]。tms9-1(Os09g0449000)編碼一個PHD鋅指蛋白,該基因在衡農S-1中存在一個T到C的置換,導致了一個氨基酸的變化從而造成雄性不育[35]。tms10(Os02g0283800)編碼一個SERK家族類受體蛋白激酶,表明激酶活性對于高溫下的絨氈層變性和雄性生育力至關重要[37]。tms18(Os10g0524500)是葡萄糖-甲醇-膽堿(GMC)氧化還原酶家族成員[38]。OsUgp1(Os09g0553200)編碼尿苷二磷酸葡萄糖焦磷酸化酶(UDP-glucose pyrophosphorylase,UGPase),該酶通過催化葡萄糖-1-磷酸和UTP形成UDP-葡萄糖和焦磷酸的可逆反應造成雄性不育和育性恢復[43]。此外,tms4[26]和tms9[34]定位于水稻第2染色體,pms2[13]和tms6(t)-1[31]定位于第3染色體,pms4[17]定位于第4染色體,tms6[30]定位于第5染色體,tms3[25]和TMS[40]定位于第6染色體,pms1(t)[44]定位于第7染色體,tms1[22]定位于第8染色體,TGMS[27]和Ms-h[41]定位于第9染色體,tms6(t)-2[32]和rtms-1[42]定位于第10染色體,tms8[33]定位于第11染色體。
本研究團隊通過60Co-γ輻射秈稻品種93-11,分離出了包括株高、分蘗數、株型、穗型、小穗結構、育性、葉色、葉型、抽穗期、苯達松抗性等突變體[46],并克隆了普通核雄性不育基因RMS2[47]、MS26-2245[46]、PTC1-2[48]和MSP1-1972[49]等。在93-11輻射誘變突變體庫后代3650號株系中篩選得到一個雄性不育突變體tms3650,對突變體進行表型鑒定和基因定位研究,以期為進一步研究該基因的功能和調控機理提供理論基礎。
1 材料與方法
1.1 試驗材料
秈稻93-11經60Co-γ輻射誘變得到M0代,輻射后的種子種植于海南省臨高縣試驗田,成熟后分單株收種,共獲得M1代材料約6500份。次年,種植M1代,每個家系種50個單株,在其中3650號株系中發現一種突變體表現為雄性不育,被命名為tms3650。所有水稻材料均按常規田間種植與管理,單株插,株行距16.3 cm×19.8 cm。93-11干種子由湖南省水稻研究所提供。
1.2 花粉育性鑒定
在海南臨高早稻季(2月中旬播種)、晚稻季(7月上旬播種)和湖南望城基地中稻季(5月中旬播種)栽培條件下,抽穗期取突變體tms3650家系不同植株的成熟花粉用1%碘-碘化鉀溶液(0.6%KI,0.3% I2,w/w)進行染色分析。此外,可育植株和不育植株套袋自交,每株3個重復,并考種統計結實率。將tms3650突變體的稻蔸在2015年、2016年、2017年冬季(11月移栽)種植于海南省陵水縣提蒙鎮水稻育種基地,稻蔸圍膜隔離種植,安排稻蔸在1~2月短日照低溫孕穗,對抽穗后的花粉粒進行碘染分析(10倍物鏡,顯微鏡型號Leica DM2500)并統計收種后結實率。陵水轉育后的種子于海南臨高和湖南望城育種基地再次播種,抽穗期取花粉碘染分析。
1.3 突變體育性轉換鑒定
對陵水收獲的轉育種子進行大田種植鑒定和實驗室品種SSR真實性鑒定。在臨高早稻季、晚稻季和湖南望城中稻季各取93-11作對照,轉育的tms3650突變體做待測樣品,編號并單本插秧,在苗期、拔節期、抽穗期、成熟期觀察記錄突變體與對照樣品的差異。根據水稻品種鑒定DNA指紋方法標準(《中華人民共和國農業行業標準: 水稻品種鑒定技術規程》,SSR標記法,NY/T 1433-2014)合成24對SSR標記(附表1),以93-11和轉育后tms3650突變體種子萌發的葉片DNA為模板,進行PCR真實性鑒定。
1.4 遺傳分析及定位群體的構建
正常大田栽培條件下,以突變體tms3650為母本,分別以野生型秈稻93-11、明恢63和粳稻中花11為父本,配制雜交種,觀察F1、F2和F3的表型,并統計野生型可育花粉植株和突變體雄性不育植株的分離比。
1.5 DNA提取及基因定位
在齊穗期觀察群體表型,取穗子包頸、花藥偏白的水稻小穗和對應分蘗的劍葉進行保存,小穗進行育性鑒定后,選不育株葉片采用傳統CTAB法[50]提DNA。根據Michelmore等[51]的BSA極端性狀混合池分析法(bulked segregant analysis)和RCA隱性群體分析法(recessive-class analysis)對tms3650進行基因定位。在Gramene在線網站(http://www.gramene.org/)合成SSR引物用于群體定位,引物交由深圳華大基因科技有限公司合成。PCR使用Biomiga的2×PCR預混合液5 μL,1 μL引物(含各0.5 μL正反向引物),1 μL模板DNA,ddH2O補足10 μL。PCR擴增程序為常規SSR程序。擴增產物使用6%的非變性聚丙烯酰氨凝膠電泳,0.1%AgNO3染色,甲醛和NaOH顯色拍照統計基因型。
利用301對均勻分布于12條染色體上的SSR標記[52]篩選93-11與明恢63之間的多態性,得到64對具有多態性的SSR標記(附表2),多態性較低,表明明恢63與93-11在基因組序列上差異較小。收tms3650和明恢63雜交F1種子,F1使用篩選得到的多態性標記驗證真實性。種植F1得到F2,F2分離得到的61個不育株用作初定位群體,構建F2分離群體的不育株的混合基因池,每個混合池隨機選20株不育株DNA樣本,利用64對多態性標記對所述混合基因池進行分析。用篩選得到緊密連鎖的標記鑒定F2分離群體的可育株,選擇雜合基因型株系收種F2,種植F2種子得到F3分離群體,F3分離得到的883個不育株用作精細定位群體。根據初定位結果繼續發展緊密連鎖標記的SSR標記(附表3),連鎖標記對F3群體進行分子鑒定。
1.6 候選基因分析
從水稻基因組注釋數據庫(http://rice.plantbiology.msu.edu/index.shtml)和NCBI秈稻93-11基因組注釋數據庫(https://www.ncbi.nlm.nih.gov/nuccore/CM012055.1?report=graph)查詢精細定位區間內的ORF并分析基因功能,水稻表達數據庫(https://ricexpro.dna.affrc.go.jp/GGEP/index.php)分析區間內基因表達譜,預測候選基因。下載候選基因全長DNA序列,使用Primer Premier 5軟件設計測序引物,合成的引物同時PCR擴增野生型93-11和突變體tms3650的DNA,擴增產物送深圳華大基因科技有限公司進行測序,測序結果使用DNAMAN軟件進行拼接和比對分析。
2 結果與分析
2.1 突變體表型分析
在湖南和海南種植的野生型及突變體材料除生育期有5~10 d差異外其他農藝性狀基本一致。野生型93-11和突變體tms3650在分蘗性狀(圖1-A)和穗型上(圖1-B)沒有明顯差異,但突變體抽穗期稍遲(比對照93-11晚約5 d)且包頸,穎殼和小花(圖1-C)沒有明顯差異,野生型93-11花藥(圖1-D)大且黃,但突變體tms3650的花藥(圖1-D)白綠且瘦小,花粉碘染結果顯示野生型93-11成熟花粉粒能被碘液染成藍黑色(圖1-E),而tms3650花粉粒不能染色(圖1-F),徹底敗育。

圖1 野生型93-11和突變體tms3650表型鑒定Fig. 1. Phenotypic comparison of wild type 93-11 (WT)and tms3650.
冬季移栽陵水南繁后,突變體tms3650花粉粒部分花粉能被碘液染成藍黑色,部分未被染色(圖1-G),經查詢陵水縣2015、2016和2017年1~2 月平 均 溫 度 為17~24℃(https://tianqi.2345.com/wea_history/59954.htm),符合光溫敏雄性不育育性轉育溫度。選擇9個突變體株系進行花粉育性恢復調查及結實率分析(表1),可育花粉和不育花粉數幾乎一致,結實率在30%左右(突變體單株有效分蘗5~6個,穗粒數150粒左右)。上述結果表明突變體花粉的育性在短日照低溫條件下得到恢復,但恢復率不高,還需要繼續尋找合適的臨界轉育溫度和光照條件。
2.2 突變體育性轉換真實性鑒定
轉育后的突變體tms3650種子在海南臨高和湖南望城基地大田種植鑒定,除了穗子包頸和生育期延遲(比對照93-11晚5 d左右),其他農藝性狀和野生型93-11一致,突變體生長整齊一致,無雜株。突變體花粉經鏡檢發現完全敗育。隨機取突變體tms3650不同單株的葉片混樣構建4個DNA池,以野生型93-11的模板DNA為對照。根據24對國標引物PAGE膠檢測結果(圖2),野生型和突變體帶型完全一致,表明所選位點沒有引入外源基因。由此得出結論,在陵水點收獲的突變體tms3650種子為真實轉育自交種,而不是串粉導致。
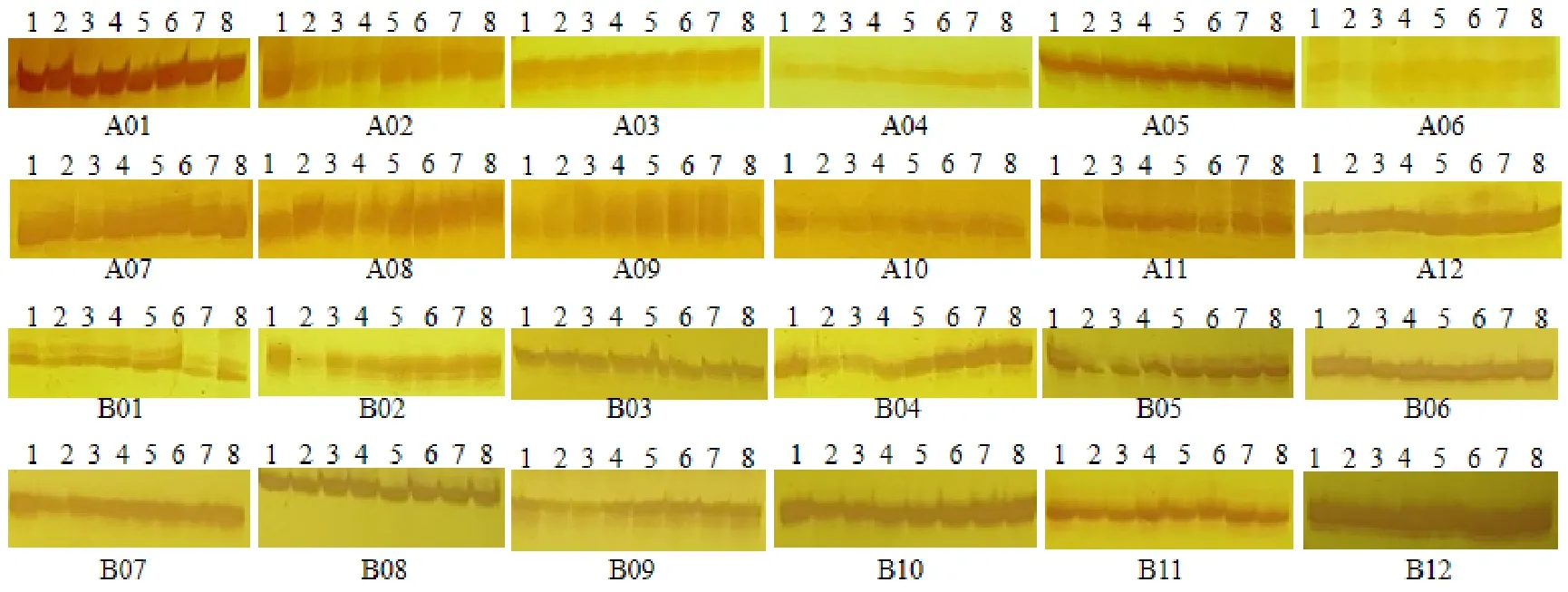
圖2 野生型93-11和突變體tms3650品種真實性鑒定Fig. 2. Authenticity identification of wild-type 93-11 and mutant tms3650.
2.3 突變體遺傳分析
突變體tms3650作為母本,野生型93-11,粳稻中花11,秈稻明恢63作為父本雜交發現F1結實正常,說明突變體雌性育性正常且該基因是一個隱性基因。統計F2分離群體中的野生型和突變體株數,經卡方檢驗發現,野生型和突變體分離比符合3∶1(表2),表明tms3650的雄性不育性狀受一對單基因控制。上述結果表明tms3650突變體受一對隱性單基因控制。

表2 突變體tms3650的遺傳分析Table 2. Genetic analysis of mutant tms3650.
2.4 初定位和精細定位
BSA初定位結果表明第3染色體上的SSR標記RM055和RM293與tms3650的突變表型存在連鎖關系(附圖1)。進一步在SSR標記RM055和RM293 附近繼續開發分子標記得到RM15900、RM15915、RM15935和RM15947。利用上述連鎖標記逐個分析明恢63和tms3650的F2群體中的不育株的基因型,得到交換單株分別為18、2、1、0、0和5個。因此將目標基因初步定位于第3染色體長臂上SSR標記RM15900和RM15947之間(圖3-A)。
在標記RM15900和RM15947之間繼續篩選多態性標記,得到RM15919、RM15921、RM15927、RM15931、RM15934和RM15937。通過分析緊密連鎖標記在F3群體883個不育單株的基因型。得到tms3650與上述RM15900到RM15947標記之間的交換單株分別為11、3、2、2、1、0、2、4、4、6個(圖3-B),tms3650與標記RM15931共分離(交換單株基因型和表型列于附表4),最終定位在135.25 kb的RM15927-RM15934區間內(圖3-C)。
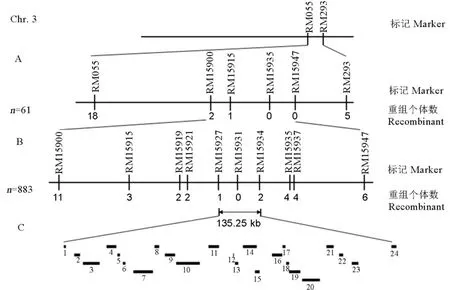
圖3 tms3650突變體的精細定位Fig. 3. Fine mapping of tms3650 mutants.
2.5 候選基因預測
根據水稻基因組注釋數據庫查詢精細定位區間內的ORF,在標記RM15927到RM15934之間135.25 kb物理距離區間內共查詢到24個預測基因(表3),與已克隆雄性不育基因進行比對發現該候選區間內沒有克隆過的光溫敏雄性不育基因(圖4)。水 稻 表 達 數 據 庫 發 現,該 區 間 內LOC_Os03g53650和LOC_Os03g53790在花藥中特異表達,其他基因在花期也有表達。選擇候選基因HTH DNA結合蛋白LOC_Os03g53600,PHD finger家族蛋白LOC_Os03g53630,半胱氨酸合酶LOC_Os03g53650,含有短鏈脫氫酶/還原酶家族結構域的蛋白質LOC_Os03g53690,PHD鋅指家族蛋白LOC_Os03g53700,醛 糖1-差 向 異 構 酶LOC_Os03g53710,SRPK4蛋白LOC_Os03g53720,RNA識別基序LOC_Os03g53770,周質β-葡萄糖苷酶前體LOC_Os03g53790對這些基因編碼區設計引物進行PCR擴增送測序,目前沒有發現造成的突變位點,正在繼續測序驗證上述基因啟動子區及區間內的其他候選基因。
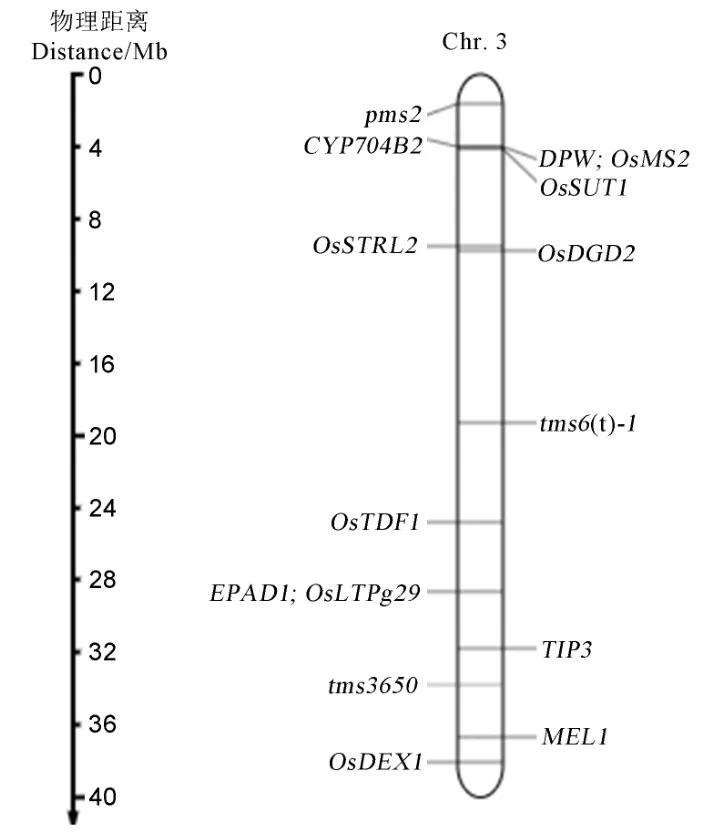
圖4 第3染色體已克隆或定位的雄性不育基因Fig. 4. Male sterility genes cloned or mapped on rice chromosome 3.
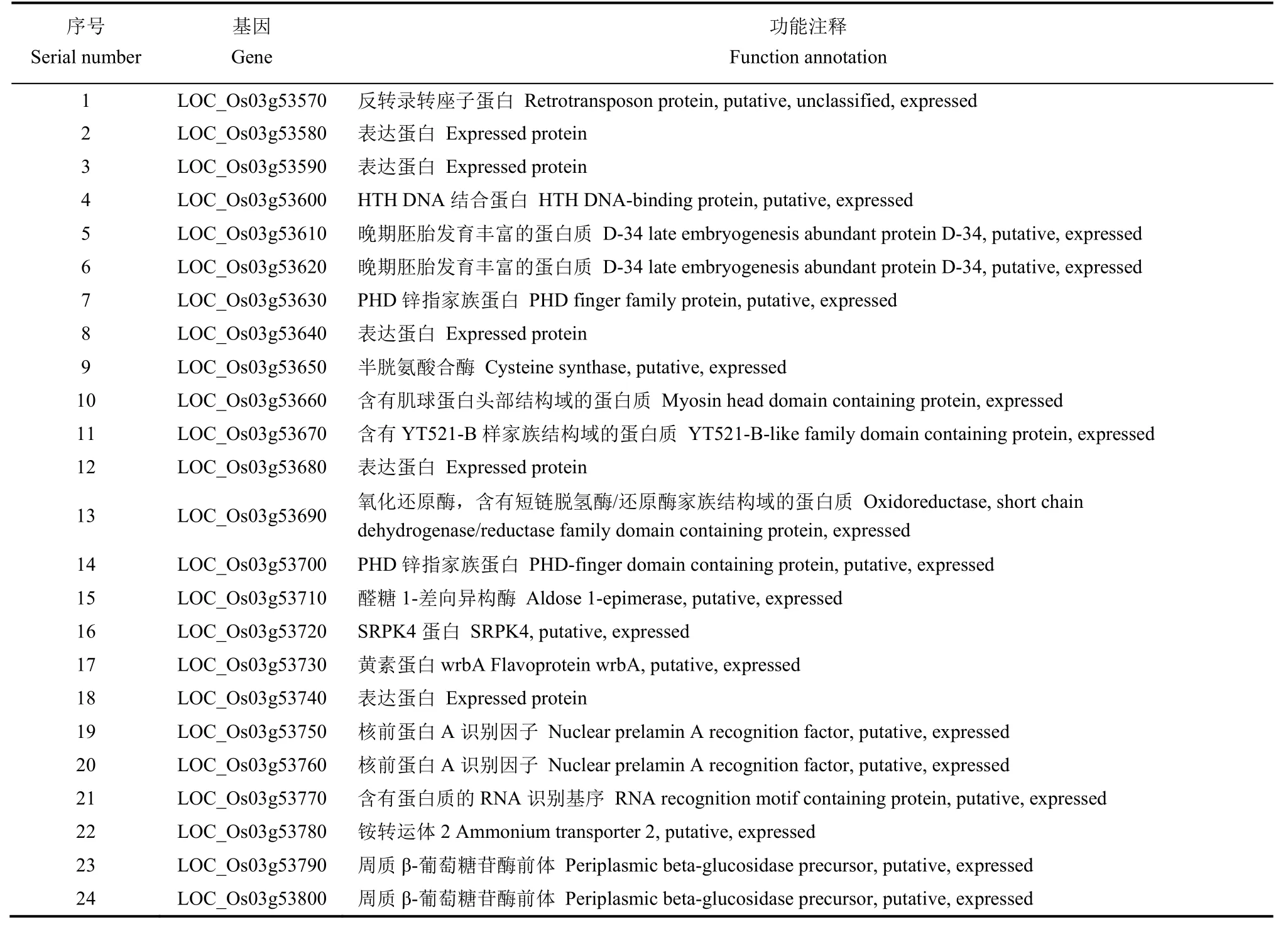
表3 目標區域預測基因Table 3. Predicted gene in target region.
2.6 緊密連鎖標記在不同品種中的多態性驗證
根據精細定位結果,我們得到了和光溫敏雄性不育基因TMS3650緊密連鎖的分子標記RM15931。為了驗證標記在指示tms3650突變體光溫敏雄性不育表型上的特異性,我們分析了在多個品種中該標記的多態性。這些品種包括兩系不育系GD-7S、隆科638S,三系保持系野香B、特B、博II B、H28B,恢復系93-11、明恢63、R51084,常規品種中花11。由圖可知(圖5),RM15931擴增出了156 bp的條帶,其他品種擴增出150 bp的多態性條帶,通過鑒定發現該連鎖標記可用于分子標記輔助選擇育種。
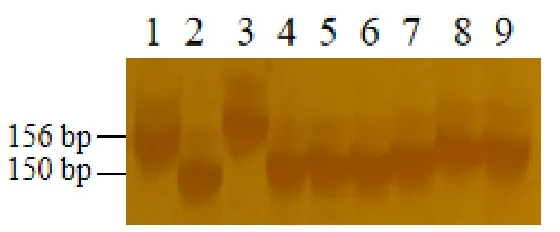
圖5 連鎖標記RM15931親本多態性鑒定Fig. 5. Identification of parental polymorphism of linked markers RM15931.
3 討論
雖然“兩系法”雜交水稻一般由隱性核雄性單基因調控,但目前已克隆的光溫敏雄性不育基因還是太少,很多遺傳機理還不是很清楚。林艷等[53]通過TALEN編輯日本晴和明恢86的pms3基因,發現突變體并不產生光溫敏雄性不育表型。王芳權等[54]發現pms3基因在培矮64S和粵光S中并不能獨立起作用,還需要與其他基因共同調控,pms3基因還受到其上游轉錄本AK111270產生的siRNA介導的甲基化調控[55]。Qi等[35]發現tms9-1與明恢63在第3外顯子上有一個T到C的差異導致氨基酸的改變導致光溫敏雄性不育,但Li等[36]發現該位點編碼一個PHD鋅指蛋白基因PTC1,在第2個外顯子T的插入使翻譯提前終止導致其普通核雄性不育,推測突變的位置不同,造成育性影響不同。OsNP1[39]是一個無花粉型普通核雄性不育基因,但tms18與OsNP1在不同位點突變會引起不一致的性狀表型[38]。由此可見,部分基因的不同突變甚至能造成不一樣的表型。因此,挖掘、鑒定新的光溫敏雄性不育基因對進一步闡明光溫敏雄性不育的分子機制具有十分重要的意義。
本研究從93-11的輻射誘變體庫中獲得了一個雄性不育突變體tms3650。表型鑒定發現突變體花藥瘦小且白綠,花粉碘染完全敗育,抽穗期比93-11稍晚5 d左右,其他農藝性狀正常。遺傳分析發現突變體受一個單隱性核基因控制,通過多年次在海南陵水冬季南繁(11月至次年4月)種植tms3650突變體稻蔸,發現在短日照(日照時間小于12 h)低溫(17~24℃)條件下部分花粉轉育并結實,收種繼續種植并通過大田鑒定和分子SSR真實性鑒定其為真實自交種,因此得出tms3650是一個光溫敏雄性不育突變體。通過將突變體和明恢63雜交得到F1,自交后代F2和F3分離群體用來定位,最終將基因定位于第3染色體長臂SSR標記RM15927到RM15934之間物理距離135.25 kb區間范圍內,且與標記RM15931共分離。
通過基因組注釋數據庫發現該區域內有24個預測基因,沒有已克隆的育性相關基因,已定位在第3染色體的光/溫敏雄性不育基因pms2[13]和tms6(t)-1[31],以及定位在第3染色體長臂的普通核雄性不育基因EPAD1[56]、TIP3[57]、OsDEX1[58]和MEL1[59]同tms3650定位區間不在一個位點,說明TMS3650是一個新基因。目前已克隆的光溫敏雄性不育基因功能有非編碼RNA、MYB轉錄因子、肌球蛋白、血清類黏蛋白、PHD鋅指蛋白、葡萄糖-甲醇-膽堿氧化還原酶、尿苷二磷酸葡萄糖焦磷酸化酶基因、SERK家族類受體蛋白激酶。通過對該區間的基因編碼區進行測序暫時沒有找到候選基因,猜測該基因可能在候選基因的啟動子區發生變異或者是其他預測不同代謝路線功能的基因,需要繼續測序并構建不同的群體對定位區間及候選基因加以驗證。
本研究進一步利用緊密連鎖標記對育種中常用的保持系進行多態性驗證,發現標記RM15931在部分常用的保持系、恢復系和常規稻之間都存在多態性,可以用于分子標記輔助選擇育種。但由于突變體在陵水南繁轉育后結實率較低,后續還需要繼續鑒定基因的臨界轉育溫度,并弄清楚該基因是光照敏感還是溫度敏感型雄性不育。本研究為該光溫敏雄性不育基因的分子標記輔助選擇育種及進一步研究該基因的功能和調控機理奠定了理論基礎。
在線輔助性信息:附表1~4和附圖1請見《中國水稻科學》網站http://www.ricesci.cn。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
