不同地理群體脊尾白蝦親本繁殖特性比較
2023-02-09 00:37:22張年國周裕華周文玉潘桂平侯文杰劉本偉王剛
水產科技情報 2023年1期
關鍵詞:質量
張年國 周裕華 周文玉 潘桂平 侯文杰 劉本偉 王剛
(上海市水產研究所,上海市水產技術推廣站,上海 200433)
脊尾白蝦(Exopalaemoncarinicauda)又名白蝦、五須蝦、迎春蝦等,隸屬于甲殼綱、十足目、長臂蝦科、白蝦屬,主要分布于我國泥沙底質的近岸淺海及河口、半咸水水域中,經過馴化也可在淡水水域中存活[1]。脊尾白蝦肉質細嫩、味道鮮美,深受消費者喜愛,具有較高的市場價值[2]。同時,該蝦具有生長速度快、繁殖能力強、環境適應性廣等特點,因而在我國沿海省份被廣泛養殖,養殖產量連年攀升,現已成為沿海灘涂地區的特色水產養殖品種之一。
目前,脊尾白蝦養殖所需苗種主要通過養殖海區自然納苗、捕撈天然苗或養殖親本自繁等方式獲得,苗種數量和質量受外部環境的影響較大,導致養殖的脊尾白蝦常常因苗種質量問題而表現出生長速度慢、抗逆能力差、個體差異顯著等特征[3],進而影響養殖經濟效益。因此,對脊尾白蝦繁殖生物學進行深入研究尤顯重要。在進行脊尾白蝦苗種繁育時,對親本進行繁殖特性的研究可以更好地揭示其繁殖規律,為人工繁殖提供科學依據。目前有關脊尾白蝦的研究主要集中在基礎生物學及養殖生態學等方面[4-8],關于脊尾白蝦繁育親本繁殖特性的研究尚未見到報道。本試驗擬通過比較不同地理群體脊尾白蝦親本的繁殖特性,探明其絕對繁殖力和相對繁殖力的差異,以期為脊尾白蝦的人工繁育提供基礎數據。
1 材料和方法
1.1 試驗材料
2020年10—12月從不同沿海區域采集脊尾白蝦野生親本。其中大豐群體(DF)采自黃海中部的川東港,如東群體(RD)采自通州灣,啟東群體(QD)采自長江口,奉賢群體(FX)和寧海群體(NH)分別采自杭州灣和三門灣。各脊尾白蝦群體的具體采集信息見表1。

表1 不同地理群體脊尾白蝦的采集和培育情況
每個群體采集50~100 kg蝦,采用活水運輸的方式運至上海市水產研究所啟東科研基地進行越冬培育。整個越冬培育期約為5個月(當年12月至次年4月),越冬期間水溫為8.0~20.0 ℃,放養密度約為22尾/m2,主要投喂冰鮮麻蝦餌料。
1.2 試驗方法
2021年4月下旬,當水溫達到18 ℃以上時,從每個越冬群體中隨機選取體表光潔、肢體完整、無損傷、活力強,且體長在3.5 cm以上的脊尾白蝦親本100尾進行抱卵率和性比分析。首先用吸水紙擦干脊尾白蝦體表水分后用游標卡尺對其進行體長測量,用電子天平進行稱量(精度為0.01 g)。然后根據其第2游泳足基部是否有生殖突起來鑒別雌雄,并將雌性親本分為抱卵和未抱卵2組,測算其性比(雌性♀/雄性♂)和抱卵率(%),計算公式如下。
性比=雌性親本數量/雄性親本數量
(1)
抱卵率(%)=(抱卵親本數量/雌性親本總量)×100
(2)
為測量不同地理群體脊尾白蝦的繁殖性能,在每個越冬群體中另外隨機選取脊尾白蝦抱卵親本30尾,完成生物學測量后,用鑷子將卵全部取下后準確稱量。然后隨機從卵中選取(0.05±0.01)g,計數所取樣品中的全部卵粒數,并參照前期研究[9],分別測算脊尾白蝦個體的絕對繁殖力(F,粒)、體長相對繁殖力(FL,粒/cm)和體質量相對繁殖力(FW,粒/g),計算公式如下。
F=W0×S/W1
(3)
FL=F/Lt
(4)
FW=F/Wt
(5)
式(3)~(5)中,S為卵樣本粒數(粒),W1為卵樣本質量(g),W0為個體所抱卵的總質量(g),Lt為樣品蝦體長(cm),Wt為蝦體樣本的總質量(g)。
1.3 數據處理
利用軟件SPSS 18.0和EXCEL 2007對數據進行統計分析,并進行相關分析及回歸分析判定,統計其相關系數,擬合篩選出繁殖力與各生物學指標的最佳回歸方程(R2值最高)。設P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果
2.1 不同地理群體脊尾白蝦繁殖親本的性比和抱卵率
不同地理群體脊尾白蝦繁殖親本的性比和抱卵率情況見表2。所選試驗群體脊尾白蝦的生長年齡相近,均為越冬培育初次性成熟個體。從表2中可以看出,本試驗脊尾白蝦繁殖親本的體長均超過4.0 cm,且除了大豐群體外,其他4組繁殖親本的體長分布均以5.0~6.0 cm為最多,其次為>6.0 cm,體長4.0~5.0 cm最少,而大豐群體則是體長在5.0~6.0 cm的最多,其次是4.0~5.0 cm,體長>6.0 cm的最少。

表2 不同地理群體脊尾白蝦繁殖親本的性比與抱卵率分析
從脊尾白蝦繁殖親本的性比來看,5組群體繁殖親本均為雄性親本多于雌性親本,但組間并無明顯差別,其中啟東群體總性比最高(0.96),寧海群體總性比最低(0.79)。對于抱卵率來說,如東群體和啟東群體的繁殖親本體長在4.0~5.0 cm范圍內的抱卵率均達到50%以上,而其他3組在該體長范圍內的抱卵率最高為20.00%。對于總抱卵率來說,寧海群體最高(40.91%),大豐群體最低(27.66%)。
2.2 不同地理群體脊尾白蝦繁殖親本的繁殖力
通過對不同地理群體脊尾白蝦繁殖親本進行繁殖力分析發現,如東群體繁殖親本的體長和體質量均顯著高于其他4個群體(P<0.05)(見表3)。不同地理群體脊尾白蝦繁殖親本的絕對繁殖力為1 699.10~2 733.72粒,平均2 164.28粒;個體體長相對繁殖力為275.93~404.36粒/cm,平均為344.80粒/cm;個體體質量相對繁殖力在455.31~596.26粒/g,平均545.52粒/g。對于不同地理群體的脊尾白蝦繁殖親本來說,如東群體的絕對繁殖力最高,其次為寧海群體,但二者之間不存在顯著差異(P>0.05),另外3個群體之間的絕對繁殖力亦不存在顯著差異(P>0.05)。各地理群體脊尾白蝦繁殖親本的體長相對繁殖力和體質量相對繁殖力的變化趨勢相近,均為啟東群體最低,如東群體和寧海群體相對較高,但體長相對繁殖力的變化趨勢更為顯著。

表3 不同地理群體脊尾白蝦的繁殖特性
2.3 脊尾白蝦繁殖親本繁殖力與主要生物學指標的相關性分析
不同地理群體脊尾白蝦繁殖親本絕對繁殖力F與體長L、體質量W等主要生物學指標間的相關系數見表4。從表4可以看出,不同地理群體脊尾白蝦繁殖親本的F與L、W均存在著顯著正相關(P<0.05)。除奉賢群體外,其他群體的F與L、W之間均達到了極顯著相關性水平(P<0.01),說明各地理群體脊尾白蝦的絕對繁殖力均隨繁殖親本體長、體質量的增加而增大,其中奉賢群體增加的幅度最小,如東群體次之,啟東群體最大。

表4 脊尾白蝦個體繁殖力與體長、體質量的相關系數情況(n=30)
用線性函數、二次函數、對數函數、指數函數和冪函數等分別對不同地理群體脊尾白蝦的絕對繁殖力F與體長L、體質量W等指標進行回歸分析,結果發現,二次函數方程可以更好地反映出脊尾白蝦絕對繁殖力與體長之間的相互關系(見表5)。除奉賢群體外,其他4組的絕對繁殖力與體長均有顯著相關性(P<0.05),其中大豐、啟東、寧海群體均極顯著相關(P<0.01)。從絕對繁殖力與體質量的關系來看,其最佳回歸方程為指數函數,除奉賢群體相關不顯著外(P>0.05),其他4組群體均極顯著相關(P<0.01)。
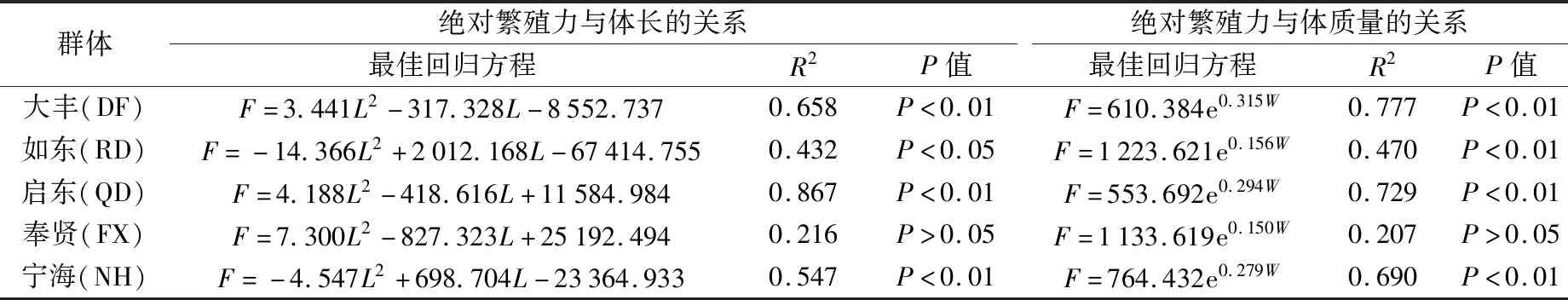
表5 不同地理群體脊尾白蝦抱卵親本繁殖力與體長、體質量的關系
2.4 不同地理群體脊尾白蝦抱卵親本的生物學最小型
不同地理群體脊尾白蝦抱卵親本的最小型生物學信息見表6。從表6可以看出,在5組脊尾白蝦抱卵親本中,奉賢群體最小抱卵個體的體長為5.14 cm,體質量為2.17 g,是所測樣本的最小群體;其次為大豐群體;如東群體最小抱卵個體的生物學指標最大,其抱卵個體最小型體長為6.11 cm,體質量為3.61 g。奉賢群體的生物學最小型雖是所測群體中最小個體,但其絕對繁殖力卻是5組中最大的,說明各地理群體脊尾白蝦的受精卵粒徑差異較為顯著,奉賢群體的受精卵粒徑相對其他群體偏小。

表6 不同地理群體脊尾白蝦抱卵親本生物學最小型的生物學信息
3 討論
3.1 脊尾白蝦繁殖親本的繁殖力分析
水生動物的個體繁殖力作為衡量一種物種繁殖狀況的重要指標,是反映種群繁衍模式和判定物種生存能力的重要指標,體現了物種或種群對環境變化的適應性[10-11]。本研究中,不同地理群體脊尾白蝦繁殖親本的個體繁殖力存在稍許差異,測得絕對繁殖力變幅為1 699.10~2 733.72粒,體長相對繁殖力平均值為344.80粒/cm,體質量相對繁殖力平均值為545.52粒/g。本試驗結果稍高于普通池養的脊尾白蝦,如于天基等[12]對山東日照地區池養脊尾白蝦進行繁殖特性研究發現,脊尾白蝦抱卵量在237~2 212粒,平均體質量相對繁殖力為259.69粒/g。李明云[13]對浙江鄞州池養脊尾白蝦的研究發現,其體質量相對繁殖力為310.53~767.93粒/g,平均為512.7粒/g。出現這種情況的原因可能與脊尾白蝦所處的養殖環境有關。本研究中的脊尾白蝦為遴選出的不同地理群體的野生種群,具有優良的遺傳性狀,因而其繁殖力也較普通池養脊尾白蝦高。裴倩倩等[6]對長江口脊尾白蝦野生種群的研究發現,其平均絕對繁殖力為(1 761±1 064)粒。在本研究中,同處于長江口的啟東群體的絕對繁殖力為(1 699.10±424.03)粒,與之相差不大,表明相同地理位置的脊尾白蝦野生種群具有較強的遺傳穩定性,其繁殖特性并未隨著時間的變化而發生較大改變。
有研究表明,水生動物的個體繁殖力與其生物學指標呈現不同的擬合關系,其相關性也存在明顯差異[14]。在本研究中,通過統計分析發現,不同地理群體脊尾白蝦的個體繁殖力分別與其體長、體質量存在著顯著的正相關(P<0.05),這表明在實際生產過程中,可根據體長、體質量等指標來大致估算脊尾白蝦的個體繁殖力,從而為其良種繁育和苗種投放提供技術參考。在對不同地理群體脊尾白蝦絕對繁殖力與生物學指標進行擬合時發現,其絕對繁殖力與體長的最佳擬合方程主要為二次函數,這與同為甲殼類口蝦蛄的研究結果[9]相似;而絕對繁殖力與體質量的最佳擬合方程主要為指數函數。這表明在實際生產運用中,應根據不同群體采用相應的函數進行方程擬合,并通過相關性檢驗找出其最佳擬合方程。
3.2 脊尾白蝦繁殖親本的性比分析
水生動物的性比是反映種群結構的重要指標,會在一定程度上影響種群數量的變動,對于繁殖群體來說顯得尤為重要[15]。有研究發現,甲殼動物的性比與物種及季節變化有關,日本沼蝦在繁殖初期雌雄比為1.11∶1,繁殖期為1.44∶1,而在非繁殖期雄蝦則多于雌蝦[16-17]。裴倩倩等[6]對長江口脊尾白蝦野生種群的研究發現,在繁殖期其雌雄比在1.18∶1,非繁殖期為1.05∶1,周年性比為1.12∶1。本研究顯示,5個地理群體脊尾白蝦繁殖期的性比在(0.79∶1)~(0.96∶1),平均性比為0.87∶1,這與其他研究存在一定差異,原因可能與取樣時間有較大關系。在本研究中,不同地理群體脊尾白蝦的取樣時間安排在4月下旬,此時池塘水溫為18~20 ℃,雌性脊尾白蝦的抱卵比例僅在27.66%~40.91%,尚處于繁殖早期,交配率不高,而雄性個體因生殖交配需要,其活動范圍較雌性廣泛,故被隨機抓捕的可能性也比雌性高。具體原因尚需進一步驗證。本研究還發現,不同地理群體的脊尾白蝦雄性個體在交配完成后存在死亡現象,而雌性個體的產后死亡率遠低于雄性,這與周氏新對蝦群體[18]的情況類似。
脊尾白蝦作為甲殼動物的一種,因其一年內可多次繁殖,故其性比發生變化的幾率非常高。若一個群體中雄性個體的數量偏多,常會影響群體的性別結構,進而導致其繁殖潛力出現下降趨勢[19]。本研究中,不同地理群體脊尾白蝦繁殖群體的性比在0.79~0.96,說明用于產卵的雌性親本少于雄性親本,導致群體繁殖潛力也偏小。因此,在養殖脊尾白蝦投放繁殖親本時,應注意調控投放親本的性比,這樣才能在養殖過程中科學地調控養殖密度和產量,有利于培育出大規格商品蝦。
3.3 脊尾白蝦親本的繁殖策略篩選
脊尾白蝦在人工養殖過程中,由于其生殖周期短,在養殖后期常會出現多世代交替的現象,進而造成群體中個體差異大、規格不齊的現象[20]。因此在投放繁殖親本時需進行嚴格篩選,從中挑選出個體規格大、繁殖力高、遺傳背景清晰的蝦作為親本進行培育。由于繁殖力體現了不同種群對環境變動的適應特征,絕對繁殖力體現種群的繁殖潛力,相對繁殖力則表現其繁殖策略[21],因此在篩選繁殖親本時,需綜合考慮親本的生物學規格、繁殖力大小等多種因素,從中篩選出最優個體來進行良種繁育。在本研究中,如東群體抱卵親本的平均生物學規格最大,其次為寧海群體,這兩個群體的絕對繁殖力和相對繁殖力也相對較高,表明如東群體和寧海群體的脊尾白蝦親本具有較好的繁殖潛力,可對其進行重點篩選用于良種繁育研究。
致謝:本研究在脊尾白蝦生物學測量方面得到上海海洋大學洪海新、羅浩煒等同學的大力協助,在此表示衷心感謝。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
