LncRNA 54770.30在黃鱔(Monopterus albus)組織及性腺中表達特征分析
2023-02-12 08:04:48夏雪平連子童田海峰孫敬鋒胡喬木
淡水漁業 2023年1期
夏雪平,連子童,田海峰,李 忠,孫敬鋒,胡喬木
(1.天津農學院,天津 300384;2.中國水產科學研究院長江水產研究所,武漢 430223)
黃鱔(Monopterusalbus),屬于硬骨魚綱(Osteichthyes)輻鰭亞綱(Actinopterygii)合鰓目(Synbranchiformes)合鰓科(Symbranchidae)黃鱔屬(Monopterus),是我國重要淡水特色養殖魚類之一。黃鱔因其肉質滑嫩,含有豐富的蛋白質,而且脂肪含量較低,具有較高的藥用價值,深受國內外廣大消費者喜愛。但是近年來,由于棲息環境的破壞和人類的過度捕撈,野生黃鱔的數量越來越少,為了滿足人們對于黃鱔的需要,黃鱔養殖業的發展和黃鱔人工繁殖技術的日漸成熟。黃鱔屬于雌雄同體魚類,具有典型的性逆轉現象[1]。由于黃鱔具有性逆轉的特點,其生殖性腺發育模式為:雌性發育階段到間性發育階段到最終的雄性發育階段,并且性腺的這一發育過程是單方向進行的不可逆的,即發生性變化后不再由雄性個體逆返為雌性個體[2],這意味著性成熟的雌性黃鱔個體數量無法維持在一穩定范圍內,在一定程度上制約了我國黃鱔養殖業的發展。因此,探究黃鱔性逆轉機制不僅具有非常重要的經濟意義,還能豐富魚類性別調控機制[3]。
生物體內存在兩種不同的RNA,一種是具備編碼蛋白質能力的編碼RNA,即信使RNA(message RNA,mRNA);一種是不具備編碼蛋白質能力的非編碼RNA(noncoding RNA,ncRNA)[4]。在ncRNA中,一般認為長度大于200核苷酸稱為長鏈非編碼RNA(Long non-codingRNA,LncRNA)。根據LncRNA與蛋白質編碼基因的位置關系,可以分為5類,分別是:a.正義;b.反義;c.雙向;d.內含子;e.基因間隔區[5]。LncRNA的作用機制非常眾多且復雜,涉及表觀遺傳學調控、性別分化、細胞內物質運輸和染色體重塑,參與細胞分化、生長發育、應激反應和疾病發生發展等多個方面[6]。黑龍江鱘(Acipenserschrencki)精巢與卵巢的LncRNA研究中發現多條LncRNA靶向多個與性腺發育和配子發生相關的基因[8]。在中華鱉(Trionyxsinensis)的精巢與卵巢的LncRNA研究中,發現多條LncRNA靶基因,包括dmrt1、sox9、cyp19a、sox3等與性別分化相關的基因[9]。王偉佳等[7]在大黃魚(Larimichthyscrocea)性腺中挖掘到了3 984個基因位點的5 162條LncRNA,2 782個LncRNA在精巢與卵巢中差異表達高度相關的LncRNA-mRNA 1 227對,并在精巢中發現了多條與雄性性別決定和性腺發育相關基因的LncRNA,說明大黃魚的性別決定及性腺發育和分化也受到LncRNA的調控。LncRNA在魚類性別決定和分化調控中發揮著復雜且重要作用,這也一直是水產動物性別分化研究的熱點之一。本研究通過對黃鱔卵巢、間性性腺、精巢的組織進行轉錄組測序,篩選到一條在黃鱔性腺不同發育時期差異表達的LncRNA 54770.30,通過生物信息學軟件分析其生物學特性,比較分析LncRNA 54770.30在不同組織和不同階段性腺組織表達水平,并定位其在不同階段性腺的表達位置,探究LncRNA 54770.30與黃鱔性逆轉的相關性,以期為揭示LncRNA在黃鱔性逆轉機制的作用提供基礎數據。
1 材料與方法
1.1 樣品的采集及制備
實驗用魚均采自湖北省仙桃市忠善合作社養殖基地,取不同時期的健康黃鱔共100尾,另取健康的60日齡黃鱔160尾,均充氧分箱暫養于長江水產研究所的活體養殖室內。采集3組不同發育時期黃鱔的性腺、心、肝、脾、腎、腦、胃、腸、肌肉和皮膚等組織,投入液氮中,然后轉入-80 ℃冰箱保存用于提取RNA。采集雌性、間性、雄性發育時期黃鱔性腺組織各3組,每組各兩份,一份保存于RNA store Reagent中用于RNA提取;另一份保存于4%的多聚甲醛(pH 7.5)用于制作組織切片,進行后續組織學鑒定及原位雜交實驗。
1.2 甲基睪酮刺激處理
將160尾60日齡的黃鱔隨機等量分為A、B、C和D 4組,在相同室溫環境下分別飼養于容積約為20 L的塑料箱中,每箱注入曝氣自來水約6 L,其中A、B和C 3組為實驗組,將17α-甲基睪酮(17-Methyltestosterone,MT)用無水乙醇溶解至500、1 000和1 500 μg/L的濃度,早晚向實驗組水箱中分別注入不同濃度的MT 400 μL至終濃度分別為100、200和300 μg/L;D組為對照組注入等量的無水乙醇,每日換水一次,1~2 d喂食等量紅蟲漿一次。兩月后采集性腺樣品分別保存于RNA store Reagent與4%多聚甲醛(pH=7.5)中,用于RNA提取、制作石蠟組織切片、HE染色。通過顯微鏡觀察性腺組織形態及細胞結構選取有明顯退化的性腺樣品,用干提取性腺組織RNA。
1.3 黃鱔總RNA提取與cDNA合成
將樣品從-80 ℃冰箱中取出置于預冷的研缽中研磨至粉末狀,取約0.1 g的各樣品組織于1 mL TRIzol中,使用TRIzol(Invitrogen)法提取各組織中的總RNA。1.5%瓊脂凝膠電泳測定RNA完整性,用NanoPhotometer-NP60測定其濃度和純度。根據PrimeScriptTMRT Master Mix試劑盒(TaKaRa)說明書合成cDNA,存于-20 ℃冰箱中備用。
1.4 熒光定量PCR
根據前期對黃鱔不同發育階段性腺轉錄組數據中獲得LncRNA 54770.30序列,利用Primer 5軟件設計特異性引物(表1),選EF-1α作內參基因,cDNA作模板,每個組織3個樣本,每個反應3個重復。遵照2×T5 Fast qPCR Mix(Vazyme)的使用說明,配成10 μL反應體系。反應在Quant Studio 5實時定量PCR系統上進行,2-△△CT法計算相對表達量;所得數據在統計軟件SPSS 26.0進行單因素方差分析(One-Way ANOVA)和Duncan′s法多重比較分析,當P<0.05差異顯著,用*表示;多重比較中,用不同字母表示差異顯著性。

表1 引物信息Tab.1 Primers used in this study
1.5 組織切片原位雜交
以LncRNA54770.30序列設計原位雜交引物(表1),通過PCR技術對探針片段進行擴增,按膠回收試劑盒(QIAGEN)說明書,對擴增產物進行膠回收,檢測DNA片段完整性與濃度,通過T7體外轉錄試劑盒(Thermo Fisher Scientific)制備探針。選取雌性階段、間性階段和雄性階段黃鱔性腺切片各兩組,首先使用二甲苯溶液進行3次時長為5 min的脫蠟反應,然后將切片置于不同濃度的乙醇(100%,100%,95%,70%,50%)依次浸洗5 min;將脫蠟后的切片置于4% PFA-PBS中固定10 min;PBS洗滌3次后用0.2 mol/L的HCl處理10 min;PBST洗滌3次后用10 μg/mL蛋白酶K消化處理。加入預雜交液(含tRNA、肝素),70 ℃預雜交2~5 h;加入預雜交液(含反義RNA探針100~200 ng),70 ℃過夜;50%預雜交液(無tRNA和肝素)+50% 2×SSC,70 ℃,15 min;2×SSC,70 ℃,2×30 min;0.2×SSC,70 ℃,2×30 min;1×MAB(100 mmol/L馬來酸,150 mmol/L NaCl,pH7.5)室溫5 min。室溫用Blocking Buffer(10%山羊血清,2%BMB,1×MAB)封閉4 h。抗體雜交:加Blocking Buffer 1 000倍稀釋的抗體(Roche Applied Science,Germany),4 ℃過夜,PBST室溫沖洗樣品。按照BCIP/NBT堿性磷酸酯酶顯色試劑盒(Beyotime)說明書進行顯色處理;顯色后PBS沖洗終止顯色;4% PFA-PBS固定10 min,PBST洗滌3×5 min;待切片稍干后用甘油封片,稍后進行拍照。
2 結果與分析
2.1 LncRNA 54770.30在不同組織的表達分析
通過qRT-PCR對黃鱔LncRNA 54770.30進行不同組織的表達分析,結果顯示LncRNA 54770.30在各組織中均有表達(圖1),肌肉中表達量最高,且顯著性高于精巢與肝,在其它各組織中表達較低且無顯著性差異。
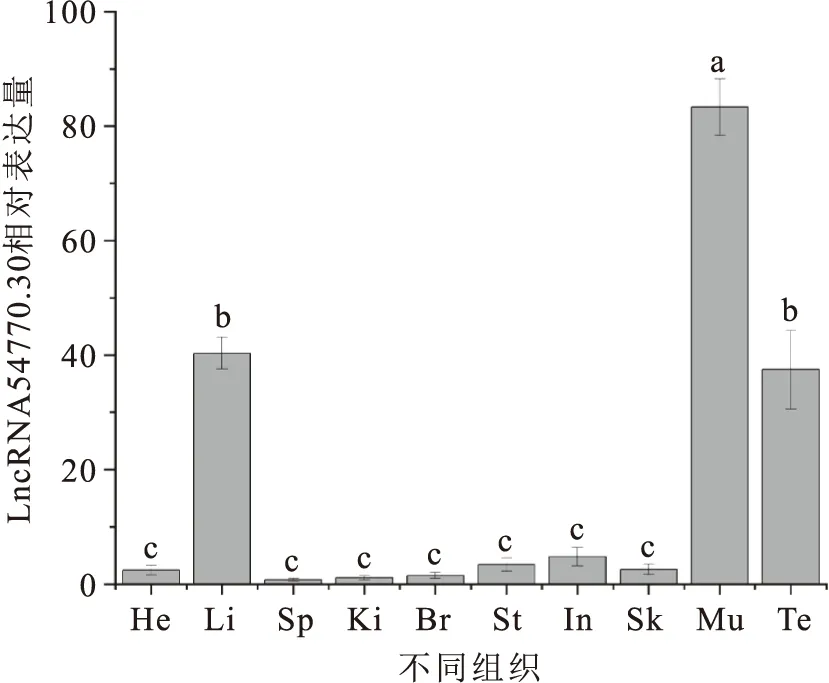
圖1 LncRNA 54770.30在不同組織中的相對表達量Fig.1 Relative expression levels of LncRNA 54770.30 in different tissues
2.2 LncRNA 54770.30在不同階段性腺的表達分析
黃鱔不同階段性腺的表達結果顯示(圖2),LncRNA 54770.30在黃鱔精巢中表達量最高、且顯著性高于雌性卵巢與間性性腺,在黃鱔雌性卵巢至間性性腺中表達量呈上升趨勢,但無顯著性差異。
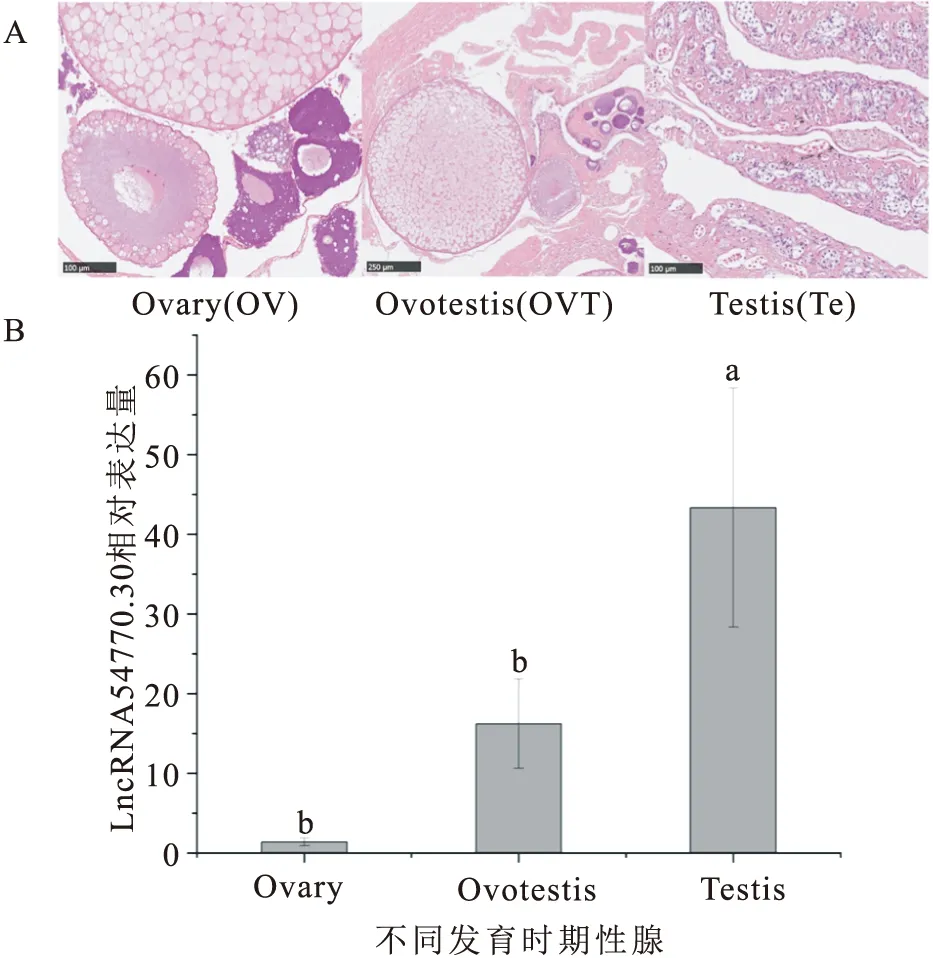
圖2 LncRNA 54770.30在不同發育時期性腺中的相對表達量Fig.2 Relative expression levels of LncRNA 54770.30 in gonads at different developmental stages
2.3 LncRNA 54770.30組織表達定位
通過RNA原位雜交定位LncRNA 54770.30在黃鱔卵巢、間性性腺和精巢中的位置表達。結果顯示,反義RNA探針原位雜交的卵巢、間性性腺和精巢組織中均可檢測到陽性信號(圖3B、C、D),正義探針均檢測不到陽性信號(圖3A)。LncRNA 54770.30在卵巢組織中,主要在初級卵母細胞和II期卵母細胞的細胞質和顆粒細胞中均有表達;在間性性腺中,主要在初級卵母細胞的細胞質、顆粒細胞以及體細胞中表達;在雄性組織中陽性信號在初級精母細胞及次級精母細胞中表達明顯。
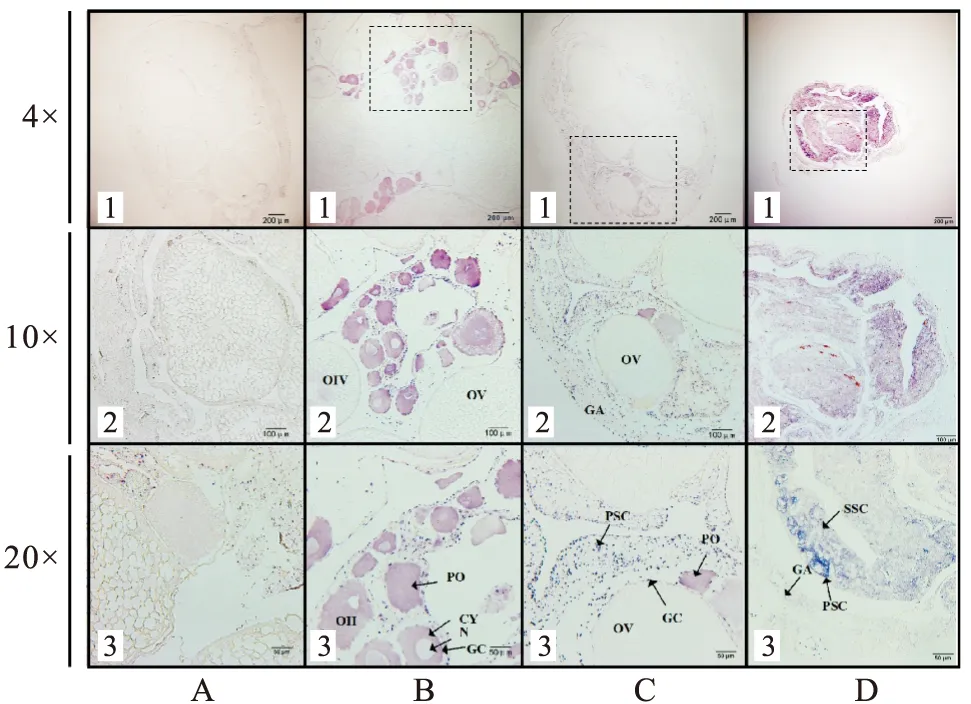
圖3 LncRNA 54770.30在不同發育時期性腺中的表達定位Fig.3 Localization of LncRNA 54770.30 in gonads at different developmental stages
2.4 MT處理后LncRNA 54770.30性腺中表達模式分析
對60日齡的黃鱔進行MT處理兩月后,HE染色切片觀察結果顯示:MT組大量卵母細胞退化,與對照組相比卵母細胞的濾泡膜與卵母細胞的胞質有明顯分離的現象,并伴隨著中空小葉出現且生殖脊明顯增粗,但未見明顯解體的卵巢(圖4A)。根據其HE染色切片從MT組選取退化較明顯的樣品提取RNA合成cDNA并進行qRT-PCR,結果顯示(圖4B)LncRNA 54770.30在卵巢中表達量降低,且與對照組表達量存在顯著差異。
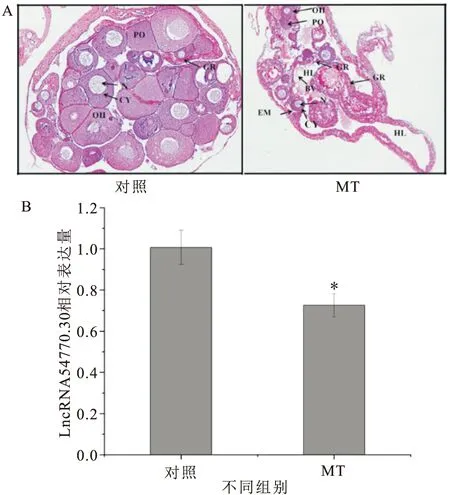
圖4 MT處理后LncRNA 54770.30在性腺中的表達Fig.4 Expression of LncRNA 54770.30 in gonad after MT treatment
3 討論與分析
黃鱔是一種雌雄同體并且雌性先熟的淡水魚類,不同于其他硬骨魚類,黃鱔早期發育為雌性,中間經歷一個雌雄同體的間性發育時期,后期則發育為雄性,該發育過程是不可逆的。黃鱔的所有個體決定性別的遺傳因素是相同的,沒有遺傳因素上的雌雄之分,因而是一種發育上的而非遺傳上的雌雄同體[10]。
LncRNA作為生物中含量較多的非編碼RNA,廣泛參與了各種生物活動的調控,有研究表明LncRNA參與了生物的性腺發育[11]。本研究通過對黃鱔卵巢、間性性腺、精巢轉錄組測序中獲的差異表達LncRNA 54770.30,通過比較分析LncRNA 54770.30在不同組織和不同階段性腺組織表達差異,結果顯示LncRNA 54770.30在肌肉中顯著性高表達(P<0.05),精巢、肝臟中次之,在胃、腸、腦、心、脾、腎、皮膚中低表達(圖1)。對比不同階段性腺表達結果顯示,LncRNA 54770.30在黃鱔雌性卵巢至間性性腺中表達量呈上升趨勢,但無顯著性差異(P>0.05),間性性腺至雄性精巢中表達量持續上升且有顯著差異(P<0.05)(圖2)。可見LncRNA 54770.30在雌性發育時期表達量較低,隨著卵母細胞生長到后期的退化,與此同時精原細胞開始增殖、分化,性腺逐漸發育到雄性階段,LncRNA 54770.30表達量在此程中逐漸上升。在小鼠(Musmusculus)、sox9上游約1.3 kb處存在LncRNA TESCO,通過與轉錄因子sf1的相互作用維持sox9的表達,從而維持雄性小鼠的發育,在小鼠中還發現了位于5號染色體上的LncRNA DMR通過結合dmrt1的3’UTR抑制dmrt1的表達[12]。在果蠅(Drosophilamelanogaster)中,sxlpe是一個位于果蠅X染色體上的雌性性別決定啟動子,可表達R1S、R1A、R2S和R2A四條LncRNAs,其中R2A通過激活sxlpe的轉錄使果蠅發育為雌性,而R1A、R1S和R2S通過抑制sxlpe使果蠅發育為雄性[13]。RONDEAU等[14]在果蠅精巢中的研究也證明了LncRNA在果蠅精子發育中發揮著不可或缺的功能。王偉佳[15]對大黃魚性腺LncRNA的研究中篩選得到了61個在卵巢中特異表達的LncRNA,得到了803個在精巢中特異表達的LncRNA,通過對這些LncRNA的預測獲得多條可能與精子發生發育相關的LncRNA,這些LncRNA可能發揮著促進性腺向精巢分化或維持精巢正常生理功能的作用;同時還發現一條與dmrt1呈現強烈正相關性的LncRNA MSTRG.24346,推測其可能在大黃魚的性別決定中發揮著重要作用。馮博[16]在半滑舌鰨(Cynoglossussemilaevis)的研究中,通過高通量測序分析獲得精巢特異LncRNA 41個,mRNA 18個;卵巢特異LncRNA 33個,mRNA 21個;偽雄魚精巢特異LncRNA 26個,mRNA 12個,發現LncRNA DMRT2-AS在精巢中表達最高,而在卵巢和腎臟中的表達最低,LncRNA DMRT2-AS的表達在雄魚的精巢中的表達均表現出持續增加的趨勢,這些結果表明LncRNA DMRT2-AS可能對dmrt2的表達具有調控作用。彭錕等[17]在羅非魚的研究中,通過轉錄組測序獲得了尼羅羅非魚(Oreochromisniloticus)雌、雄性腺中呈現差異表達的LncRNA,并選取LncRNA TCONS-02477925開展研究,結果表明LncRNA TCONS-02477925在羅非魚卵母細胞發育和性別分化中可能有重要的作用。SONG等[18]在關于鯉魚(Cyprinuscarpio)的研究中通過對干擾igf3前后的性腺組織進行測序,得到了124個差異表達的LncRNA,這些LncRNA可能在鯉魚的性別分化和性腺發育中起到重要的作用。目前LncRNA在黃鱔性逆轉中的研究報導較為罕有,但參照以上LncRNA在其他魚類和其他物種中的研究結果,對比分析本研究結果所展示的LncRNA 54770.30在黃鱔的卵巢和精巢中的表達存在十分顯著的差異且在精巢中的表達明顯高于在卵巢中的表達,可推測LncRNA 54770.30可能在黃鱔的性腺發育和性別分化中起到重要的作用。另外,本研究中黃鱔LncRNA 54770.30在肌肉中顯著性高表達(P<0.05),推測性腺分化與LncRNA 54770.30在肌肉中的高表達有可能存在一定聯系。
原位雜交是指將特定標記的已知順序核酸為探針與細胞或組織切片中核酸進行雜交,從而對特定核酸順序進行精確定量定位的過程。本研究通過RNA原位雜交來定位LncRNA 54770.30在黃鱔卵巢、間性性腺、精巢細胞中的位置表達(圖3),結果顯示在使用反義RNA探針原位雜交的卵巢組織、間性性腺、精巢組織中均可檢測到陽性信號(圖3B、C、D),與本研究qPCR結果中顯示的LncRNA 54770.30在不同發育時期性腺結中均有表達保持高度一致。LncRNA 54770.30在卵巢組織中主要在初期卵母細胞和II、IV時相期卵母細胞的細胞質和顆粒細胞中表達,V時相期卵母細胞的細胞質中無明顯陽性信號;在間性組織中主要初級卵母細胞的細胞質和顆粒細胞中表達,在性腺外膜中有少量表達,在V時相期卵母細胞的細胞質中無明顯陽性信號,陰性對照組中未見陽性信號。何智等[19]對不同發育時期黃鱔性腺中Caspase-3定位分析中也有類似情況,推測可能是因為此時的卵母細胞細胞質中充滿了大量卵黃物質和脂質,使細胞質中陽性信號無法觀察到。在雄性組織中,陽性信號在初級精母細胞及次級精母細胞中表達明顯,在性腺外膜中有少量表達。蔣嬌云等[3]的研究中黃鱔的f4基因原位雜交分析結果顯示與半定量具有類似的表達情況,即f4基因從IV期卵巢開始表達,一直持續到后期精巢,間期性腺到后期精巢表達量有增高趨勢,最終性腺性逆轉為精巢。與本研究中LncRNA 54770.30在卵巢至間性性腺和精巢中的表達呈逐漸上升趨勢且在精巢中最高具有相同的表達趨勢。
17α-甲基睪酮(17-Methyltestosterone,MT)是一種人工合成的白色結晶性粉末狀雄性激素,屬于固醇類激素,能夠干擾動物正常的內分泌系統,促進并維持雄性性器官的發育及第二性征的形成和維持,對雌激素產生拮抗作用,抑制雌性性器官的發育,可做為魚類性轉化誘導劑,常用于苗種培育和性別控制等方面的特效藥物據[20]。TANG等[21]報導多種雄激素(甲基睪酮、睪酮和11-酮睪酮)均對成鱔提早性轉化無效,但對黃鱔胚胎后期以及仔鱔進行雄激素處理可以促使黃鱔提早性轉換。本研究通過在飼養黃鱔的水體和餌料中添加甲基睪酮,對仔鱔進行激素刺激處理,通過對比觀察性腺樣品的HE切片,MT組大量II期卵母細胞退化,與對照組相比發現卵母細胞的卵膜與卵母細胞的胞質有明顯分離的現象,并伴隨著中空小葉出現且生殖脊明顯增粗,但未見明顯解體的卵巢。MT處理后LncRNA54770.30表達相較于對照組明顯下降,具有顯著性差異(P<0.05)。卓孝磊等[22]在外源性甲基睪丸酮對黃鱔性腺發育的研究中發現MT可促進雌鱔卵母細胞退化,但不能使卵巢完全解體;MT可促進雌性和間性黃鱔性腺生殖板擴大、延伸并出現中空小葉;MT對雌雄間體早期的黃鱔的雄性生殖細胞的發育有促進作用。KUO等[23]報道了雌雄同體、雌性先熟的藍點鮨(Epinephelinaefario)自然生長需要7年才會變為雄魚,但是用甲基睪酮可誘發2齡魚提前性轉變為雄魚。方永強等[24,25]最初用17α-甲基睪酮研究對赤點石斑魚(E.akaara)性逆轉的影響,并認為17α-甲基睪酮引起石斑魚性逆轉的作用機制為:17α-甲基睪酮抑制卵母細胞生成卵黃,使卵母細胞出現閉鎖,最終退化,并通過刺激生精細胞的增值使精巢增值,激發精子發生機制,最終引起石斑魚性逆轉。綜上所述與本實驗結果可推測甲基睪酮可能抑制黃鱔卵母細胞積累卵黃,阻止了卵母細胞的進一步發育;同時也能刺激原始精母細胞的發育而使黃鱔由雌性發育階段提早進入性逆轉階段,而LncRNA 54770.30在此過程中可能參與黃鱔卵母細胞發育及性逆轉過程中的卵母細胞的凋亡和精母細胞的增殖。
