bta-miR-1343-3p靶向NEDD8調控牛卵泡顆粒細胞增殖與凋亡機制初探
2023-02-16 02:44:52馬曉燕郭翔宇賈凱琪呂麗華
山西農業科學 2023年2期
關鍵詞:檢測
馬曉燕,郭翔宇,賈凱琪,成 穎,呂麗華
(山西農業大學 動物科學學院,山西 太谷 030801)
在哺乳動物生殖過程中,顆粒細胞的增殖和凋亡在卵子發生、卵泡發育和閉鎖中起到至關重要的作用。卵泡發育是一個有序復雜的過程,受到下丘腦-垂體-性腺軸的調控,并呈現周期性變化[1-2]。生長卵泡發育至有腔卵泡之前不依賴促性腺激素,有腔卵泡的發育則依賴促性腺激素的作用,同時受到卵母細胞—顆粒細胞—膜細胞三者的協同調控[3]。卵泡生長過程中卵母細胞以自分泌和旁分泌的方式與卵泡生態體系中的體細胞共同發揮作用,影響卵泡發育進程,并決定其最終命運。顆粒細胞是卵泡體系中數量最多的體細胞,在卵泡發育過程中發揮了至關重要的作用[4]。雌性哺乳動物從早期胚胎性腺發育到性成熟過程中,大量卵泡閉鎖,調控卵泡閉鎖的機制錯綜復雜,調控過程中的些許偏差都會導致卵泡發育阻滯和閉鎖。雌性哺乳動物在生命早期形成的卵巢儲備是相對固定的,卵子儲備量隨著動物排卵而不斷減少。隨著生存環境的改變,越來越多的哺乳動物包括人類都面臨著由于卵泡生長調節異常導致的卵巢早衰致使生育年限縮短,因此,研究卵泡閉鎖的發生機制將進一步解開卵泡閉鎖的未解之謎。
目前,研究尚未揭開胚胎期卵泡閉鎖的發生機制,而生長期卵泡閉鎖的主要原因可歸結為顆粒細胞過度凋亡[5-6]。miRNA是一種內源的非編碼小RNA,目前,在人類中已鑒定出數百種 miRNAs[7-8]。由于這些miRNAs序列在不同哺乳動物間高度保守,因此,可推測哺乳動物體內絕大多數與發育和疾病相關的事件均與miRNAs的調控有關[9]。成熟的miRNA可與其靶基因的3′UTR結合并發揮調控作用[9],此外,miRNA可直接與蛋白質相互作用調控基因翻譯過程[10]并影響表觀遺傳機制[11],也可通過抑制DNA轉錄或破壞mRNA分子的穩定性實現對靶基因的負調控[12]。哺乳動物中,30%以上的蛋白質編碼基因受到miRNAs的調控[13]。目前,在雌性哺乳動物生殖領域對miRNA的研究主要與卵泡發育有關,如顆粒細胞增殖、凋亡和類固醇激素分泌等方面。
TU等[14]研究發現,miRNAs在卵母細胞、顆粒細胞、膜細胞和卵泡液中廣泛表達。從原始卵泡形成,卵巢儲備建立到初情期卵泡在促性腺激素作用下開始發育再到最終排卵,大量的miRNAs參與了上述過程[14-15]。近年來研究發現,多種miRNAs參與了顆粒細胞調控。ZHANG等[16]研究發現,miR-17-5p可通過靶向調控E2F轉錄因子1(Transcriptional factor,E2F1)可影響豬卵泡顆粒細胞的功能,在豬卵泡顆粒細胞中過表達miR-17-5p顯著降低了轉錄因子E2F1的活性,促進了顆粒細胞生長和激素合成過程。蔣秀敏等[17]研究發現,miR-483-5p可通過調控細胞外信號調節激酶(Extracellular regulated protein kinases,ERK)的表達調控人卵泡顆粒細胞增殖和凋亡過程以及絲裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK)在各種細胞反應中的作用[18]。
本研究基于前期在牛卵泡顆粒細胞中開展的全轉錄組測序結果,選擇bta-miR-1343-3p為研究對象,進一步研究其在顆粒細胞增殖和凋亡中的調控作用,旨在初步揭示哺乳動物卵泡閉鎖的調控機制。
1 材料和方法
1.1 試驗材料
本研究所用的牛卵巢均來自山西省呂梁市文水縣屠宰場,采集后采用75%酒精消毒并保存于4 ℃ DPBS中,2 h內運回實驗室。
1.2 主要試劑
DMEM/F12基礎培養基(HyClone公司,美國),胎牛血清(FBS)(賽澳美細胞(技術)有限公司,北京),無菌PBS緩沖液(博士德生物,武漢),MTT試劑盒(博士德生物,武漢),Lipofectamine 3000(Invitrogen公司,美國),E.Z.N.A.?Plasmid mini Kit I(Omega BioTek,美國),Mut Express II Fast Mutagenesis Kit(南京諾唯贊生物科技有限公司,武漢),Dual-Luciferase Reporter Assay System(Promega Co. Ltd,美國),ClonExpress?Ultra One Step Cloning Kit(南京諾唯贊生物科技有限公司,武漢),bta-miRNA-1343-3p模擬物與抑制物(漢恒生物科技有限公司,上海),2×Es Taq Master Mix(Dye)(康為世紀生物科技有限公司,江蘇)。
1.3 卵泡顆粒細胞的收集與培養
采用眼科剪分離卵巢皮質中的有腔卵泡。將卵泡用眼科剪剪開,用細胞刮刀輕輕刮取卵泡內壁獲取顆粒細胞并收集卵泡液,1 000 r/min下離心5 min,棄上清。得到的細胞沉淀用新鮮無菌PBS重懸,1 000 r/min下再離心3次,每次5 min,清洗沉淀中的血細胞等雜質。清洗后的細胞沉淀用完全培養基(含10%的FBS和雙抗)重懸,置于10 cm細胞培養皿中,37 ℃、5% CO2下培養24 h后觀察細胞狀態和密度,更換新的完全培養基,待細胞密度達到80%~90%時傳代培養。
1.4 靶基因預測
對高通量測序結果進行靶基因篩選分析,結合測序結果,進一步使用TargetScan(TargetscanHuman 7.1:predicted miRNA targets of miR-1343-3p)和miRanda(miRDB-miRNA Target Prediction Database)篩選bta-miR-1343-3p的靶基因。最終選擇與卵泡顆粒細胞存活相關的基因NEDD8,使用RNA Hybird分析bta-miR-1343-3p與NEDD8基因UTR的結合位點。
1.5 NEDD8基因3” UTR雙熒光素酶報告載體的構建
根據NCBI網站公布的牛NEDD8基因mRNA序列,尋找NEDD83” UTR的靶點位置。采用Primer 5.0設計NEDD83” UTR擴增引物,并在引物上下游加入酶切位點與雙熒光素酶報告載體的同源序列。進一步使用CE Design軟件(南京諾唯贊生物科技有限公司)設計同源重組及突變引物序列,如表1所示。顆粒細胞收集,RNA提取與反轉錄程序參考之前的研究[19]。以牛卵泡顆粒細胞cDNA為模 板 ,使 用 2×Es Taq Master Mix(Dye)擴 增NEDD8基因3” UTR,具體步驟參考之前的研究[20],PCR產物采用2%瓊脂糖凝膠電泳鑒定,目的片段回收純化后測序鑒定序列準確性。
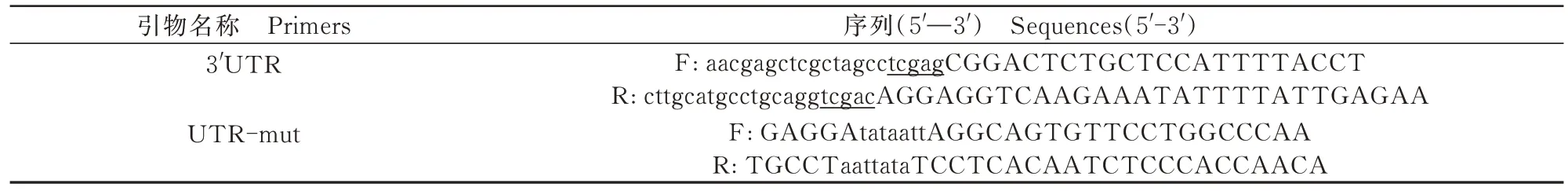
表1 bta-miR-1343-3p熒光定量PCR引物序列Tab.1 Primer sequences for qRT-PCR of bta-miR-1343-3p
將目的片段與線性化的pmir-GLO載體用同源重組酶連接,具體步驟按照ClonExpress?Ultra One Step Cloning Kit說明書(南京諾唯贊生物科技有限公司,武漢)進行,重組產物轉化感受態細胞。經轉化的感受態細胞短暫離心后涂布到LB固體培養基上(含氨芐青霉素),37 ℃過夜培養,挑取單個菌落搖菌測序。將測序結果正確菌液進一步培養,使用E.Z.N.A.?Plasmid Mini Kit I提取質粒DNA,獲得野生型重組質粒。
突變型雙熒光素酶報告載體的構建采用Mut Express II Fast Mutagenesis Kit(南京諾唯贊生物科技有限公司,武漢)進行3” UTR miRNA預測靶點的突變,以連接成功的野生型載體為模板,使用點突變引物引入報告載體序列突變。突變質粒經PCR擴增,重組酶重組后轉化感受態細胞,挑選單克隆菌株培養后測序,測序驗證正確的菌液進一步培養質粒DNA,獲得突變型質粒。
1.6 雙熒光素酶活性檢測
將293T細胞接種至24孔細胞培養板中,培養6~12 h,待細胞匯合度達到60%后轉染。試驗組分為3組:(I)wt載體+mimics NC,wt載體+mimics;(II)mut載體+mimics NC,mut載體+mimics;(III)空載體+mimics NC,空載體+mimics。將上述載體質粒 800 ng和 50 μm mimics使用 Lipofectamine 3000共轉染293T細胞,反應6 h后更換培養基,繼續培養48 h,檢測293T細胞的雙熒光素酶活性。轉染48 h的293T細胞用無菌PBS清洗后加入100 μL 1×PLB,室溫下裂解 15 min,吸取 20 μL 裂解液至96孔酶標板,加入100 μL LARⅡ后迅速在酶標儀中檢測螢火蟲熒光素酶活性,檢測完畢后立即加入100 μL終止液,檢測海腎熒光素酶的活性。
1.7 卵泡顆粒細胞中bta-miR-1343-3p mimics和inhibitors的轉染
將卵泡顆粒細胞均勻接種于12孔細胞培養板中,培養6~12 h,待細胞匯合度達到60%時采用Lipofectamine 3000轉染bta-miR-1343-3p mimics和inhibitors,6 h后更換完全培養基,繼續培養48 h。試驗組與對照組詳細引物序列如表2所示。

表2 bta-miR-1343-3p 模擬物與抑制物序列Tab.2 Sequences for bta-miR-1343-3p mimics and inhibitors
1.8 MTT法檢測顆粒細胞增殖率
將卵泡顆粒細胞均勻接種至96孔板,培養6~12 h。當細胞匯合度達60%時采用Lipofectamine 3000轉染 bta-miR-1343-3p mimics和 inhibitors,繼續培養24、48、72 h后移除培養液。每孔加90.0 μL的新培養基和10 μL MTT,37 ℃恒溫培養箱內繼續培養4 h,再次移除培養液,加入110.0 μL的Formazan溶解液,避光孵育15 min,490 nm波長下測定吸光值。
1.9 qRT-PCR
使用Primer 5.0設計qRT-PCR引物。其中,miRNA下游引物為M5 miRNA qPCR Assay Kit中自帶引物,序列如表3所示。mRNA實時熒光定量流程按照TB GreenTMPremix Ex Taq TMⅡ試劑盒詳細步驟進行。miRNA熒光定量使用M5 miRNA qPCR Assay Kit進行,反應體系為:cDNA 1.0 μL,M5 miRNA qPCR Mixture(2×)10.0 μL,bta-miR-1343-3p Primer(10 μmol/L)0.4 μL,Reverse Primer(10 μmol/L)0.4 μL,加 ddH2O 20.0 μL。反應程序為:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s,60.5 ℃ 1 min,45個循環;95 ℃ 15 s。

表3 熒光定量PCR引物序列Tab.3 Primer sequences of qRT-PCR
1.10 數據分析
miRNA實時熒光定量數據的處理以U6作為內參基因,采用2-ΔΔCt法計算miRNA相對表達量;靶基因表達量檢測采用RPLP0作為內參基因,MTT 結果采用式(1)計算。

雙熒光素酶報告基因檢測數據以螢火蟲熒光素酶的熒光強度與海腎熒光素酶的熒光強度比值作為熒光值。數據采用平均值±標準差表示,所有試驗均重復3次,經SPSS 22.0進行單因素方差分析,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果與分析
2.1 bta-miR-1343-3p靶基因預測結果
采用miRanda軟件預測bta-miR-1343-3p的靶mRNA,為進一步證實預測的準確性,采用TargetScan、miRDB軟件對miRanda預測結果進行驗證,結合2個軟件預測結果發現,NEDD8基因的3” UTR存在bta-miR-1343-3p的種子區結合位點(圖1),RNA hybrid結果如圖2所示。
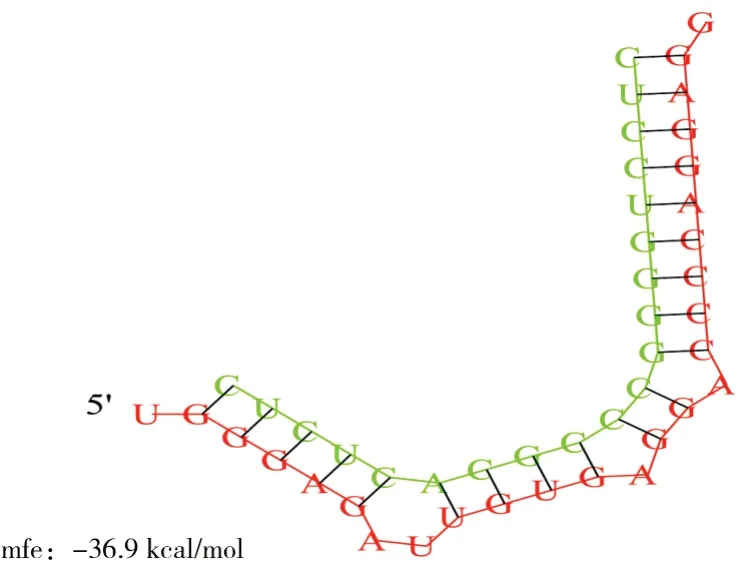
圖1 bta-miR-1343-3p與NEDD8 mRNA 3'UTR 結合位點Fig.1 Binding sites between bta-miR-1343-3p and NEDD8 mRNA 3'UTR

圖2 bta-miR-1343-3p與NEDD8 mRNA 3'UTR的雜交位點Fig.2 Hybridization site of bta-miR-1343-3p and 3'UTR of NEDD8 mRNA
2.2 雙熒光素酶報告基因檢測結果
雙熒光素報告檢測結果如圖3所示。
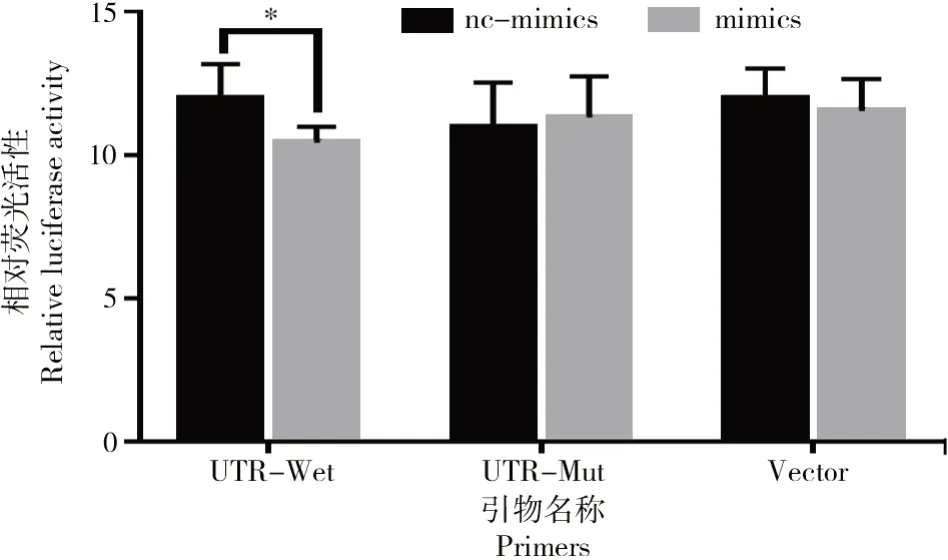
圖3 雙熒光素報告檢測結果Fig.3 Results of dual luciferase reporter assay
共轉染雙熒光素酶報告基因載體與和bta-miR-1343-3p mimics后檢測細胞內螢火蟲熒光素酶和海腎熒光素酶的活性,結果表明(圖3),在野生型報告載體+mimics組中,與對照組相比,bta-miR-1343-3p mimics組的熒光素酶活性顯著低于對照組(P<0.05),而其他組中熒光素酶活性無顯著差異。
2.3 bta-miR-1343-3p對卵泡顆粒細胞增殖的影響
在卵泡顆粒細胞中轉染bta-miR-1343-3p mimics以及抑制物(inhibitors)并采用MTT法檢測細胞增殖情況,結果如圖4所示。
從圖4可以看出,顆粒細胞中轉染mimics后24、48、72、96 h后細胞增殖率顯著下降(P<0.05);轉染inhibitor后,細胞增殖率相比對照組顯著上升(P<0.05)。
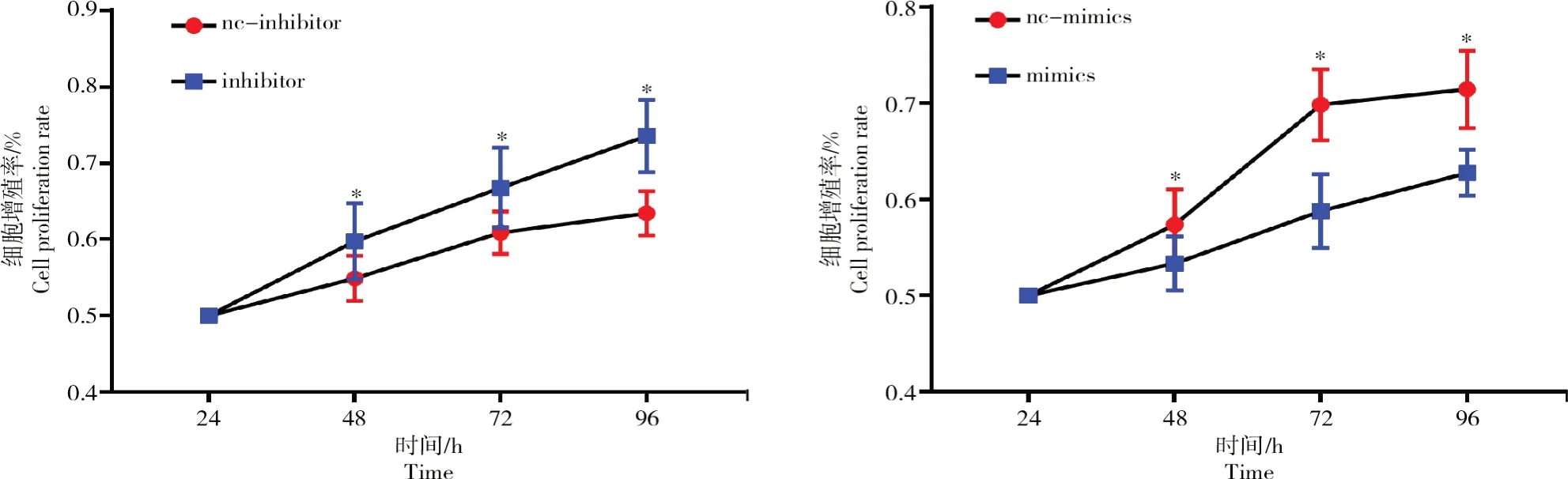
圖4 bta-miR-1343-3p對牛卵泡顆粒細胞增殖率的影響Fig.4 Influences of bta-miR-1343-3p on proliferation rate of bovine follicular GCs
2.4 bta-miR-1343-3p對卵泡顆粒細胞增殖與凋亡相關基因的影響
在牛卵泡顆粒細胞中轉染bta-miR-1343-3p mimics和inhibitors后bta-miR-1343-3p結果如圖5所示。
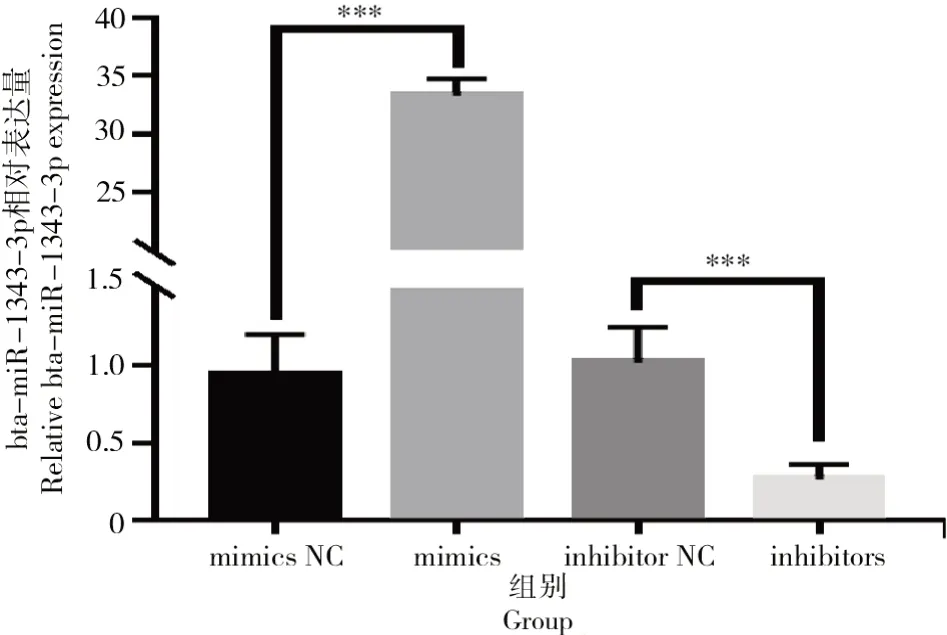
圖5 轉染mimics與inhibitors之后bta-miR-1343-3p相對表達量Fig.5 Relative expression level of bta-miR-1343-3p after mimics and inhibitors transfection
由圖5可知,與NC相比,mimics組中bta-miR-1343-3p的表達量極顯著高于對照組(P<0.001);轉染 inhibitors后,相比對照組,試驗組中bta-miR-1343-3p表達量極顯著下降(P<0.001)。
采用qRT-PCR檢測轉染bta-miR-1343-3p mimics和inhibitors后,卵泡顆粒細胞中NEDD8基因的表達情況如圖6所示。
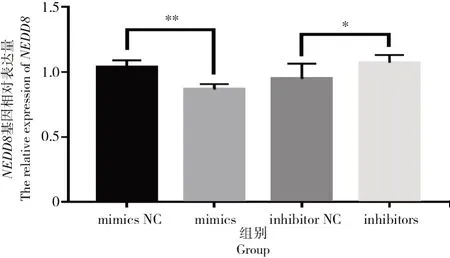
圖6 轉染mimics與inhibitors之后NEDD8相對表達情況Fig.6 Relative expression level of NEDD8 after mimics and inhibitors transfection
由圖6可知,相比對照組,轉染mimics的細胞中NEDD8基因的表達量極顯著下降(P<0.01),而轉染inhibitors細胞的NEDD8基因的表達量顯著上升(P<0.05)。
按照上述方法對細胞增殖與凋亡相關基因進行表達量的驗證,結果如圖7所示,轉染bta-miR-1343-3p mimics和inhibitors后,細胞周期相關基因CDK1與CDK4在過表達bta-miR-1343-3p后表達量顯著下降(P<0.05);相反,抑制bta-miR-1343-3p表達后,CDK1與CDK4基因的表達量顯著上升(P<0.05)。檢測細胞凋亡相關基因Bax與Fas后發現(圖8),過表達bta-miR-1343-3p后Bax表達量顯著上升(P<0.05),Fas表達量升高但差異不顯著,但在抑制bta-miR-1343-3p表達后,試驗組中Bax與Fas表達量均極顯著下降(P<0.01)。
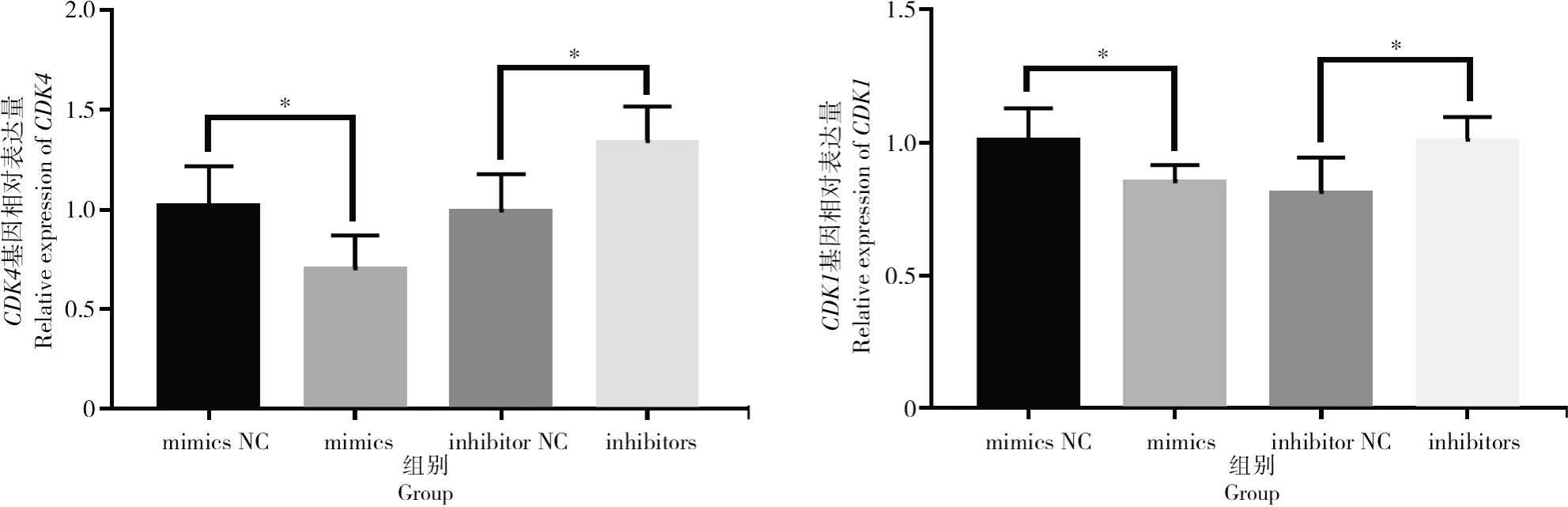
圖7 轉染mimics與inhibitors后細胞周期相關基因CDK1和CDK4的相對表達情況Fig.7 Relative expression level of cell cycle-related genes CDK1 and CDK4 after mimics and inhibitors transfection
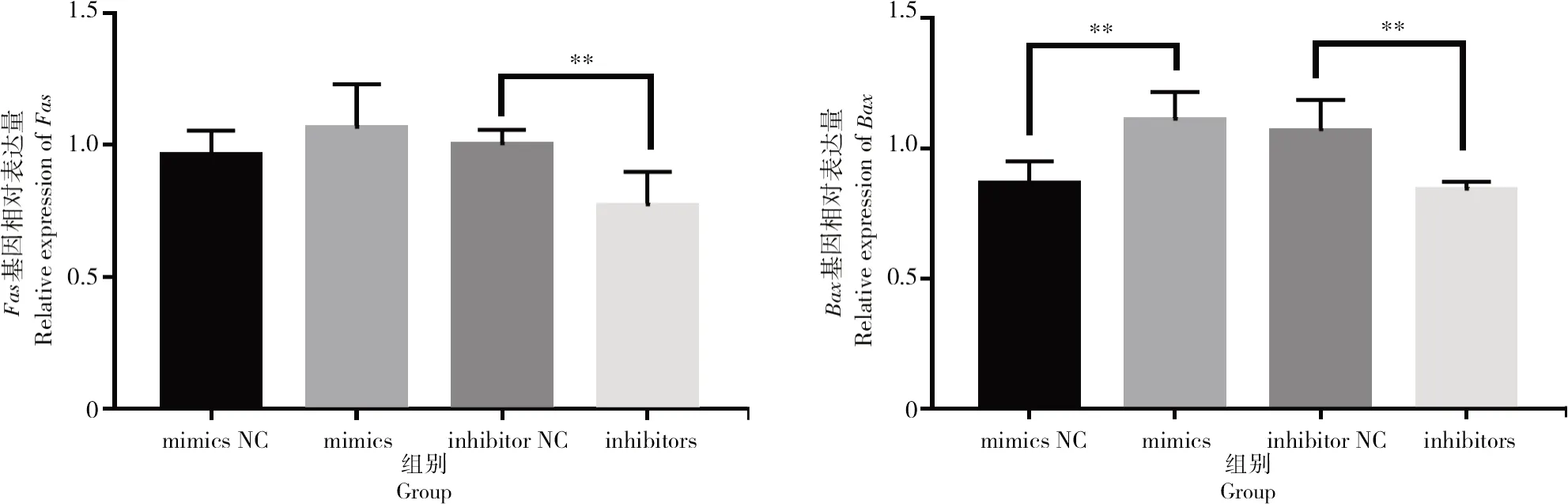
圖8 轉染mimics與inhibitors后凋亡相關基因的相對表達情況Fig.8 Relative expression level of apoptosis-related genes after mimics and inhibitors transfection
3 結論與討論
腔前卵泡的生長不依賴促性腺激素,而有腔卵泡的生長依賴FSH和LH[21-23]。卵泡生長過程中健康的卵母細胞以自分泌和旁分泌方式與卵泡生態體系中的顆粒細胞共同發揮作用,影響卵泡發育進程,并影響卵泡的最終命運。因此,本研究主要圍繞有腔卵泡中bta-miR-1343-3p對卵泡顆粒細胞的影響開展,分離卵巢皮質中大小為4~8 mm的生長期有腔卵泡并進行體外培養。
采用miRanda軟件對bta-miR-1343-3p的靶位點進行預測,發現其與NEDD8基因mRNA的3” UTR存在潛在的結合位點,bta-miR-1343-3p成熟序列在人和小鼠中完全保守。進一步采用TargetScan和miRDB數據庫預測bta-miR-1343-3p的靶基因并得到相同的結果,因此,認為預測結果準確。成熟的miRNA可與Argonaute蛋白組裝成miRNA誘導的沉默復合物(miRISC),并通過miRNA種子區域(2~8位核酸互補配對)和mRNA 3” UTR內的互補位點之間的配對與靶標mRNA結合[24],NEDD8mRNA與bta-miR-1343-3p RNA的分子雜交圖顯示,二者間存在靶向關系。通過構建NEDD8mRNA 3” UTR的雙熒光素酶野生型載體以及種子區突變型載體,轉染293T細胞后證實了bta-miR-1343-3p與NEDD8基因之間存在靶向關系。將bta-miR-1343-3p mimics與inhibitors轉染至牛卵泡顆粒細胞中后,發現btamiR-1343-3p表達量極顯著上升,而轉染抑制物后bta-miR-1343-3p表達量極顯著下降,證明可將bta-miR-1343-3p在卵泡顆粒細胞中過表達與沉默同時發現bta-miR-1343-3p在顆粒細胞中過表達后,NEDD8表達量顯著降低。表明bta-miR-1343-3p與NEDD8基因之間存在負調控作用,btamiR-1343-3p在顆粒細胞中可通過調節NEDD8基因的表達影響卵泡顆粒細胞的功能。
作為一種新的泛素樣蛋白,NEDD8蛋白在細胞中對蛋白質的分解與調控作用起著非常重要的作用[25]。NEDD8蛋白調控細胞中多種代謝途徑的方式為Neddylation,與底物結合后可進行復雜的級聯反應[26]。Neddylation激活Cullins的泛素E3連接酶活性后,NEDD化的Cullins蛋白構象發生變化并誘導底物發生泛素化,進而影響細胞內多種生命活動[27]。此外,Neddylation還可與泛素化過程競爭,p53和表皮生長因子(Epidermal growth factor receptor,簡稱為EGFR)上的某些賴氨酸殘基既可以與NEDD8結合又可與泛素結合,抑制或降解底物。研究發現,Neddylation可通過誘導細胞凋亡、衰老和自噬抑制腫瘤細胞的生長[28-29]。卵泡閉鎖過程不僅與卵泡內激素水平有關,也與卵泡顆粒細胞的凋亡高度相關。卵泡閉鎖存在于卵泡發育的整個過程,由于閉鎖卵泡發育階段的不同導致其閉鎖的具體過程存在差異,閉鎖早期均出現卵泡顆粒細胞的凋亡[30]。研究發現,與正常卵泡相比,閉鎖卵泡中凋亡相關基因p53表達量顯著上升[31-32]。p53蛋白在細胞生存與凋亡過程中發揮作用依賴于泛素化、磷酸化、乙酰化等修飾過程[31,33]。此外,p53蛋白的活性由cullin-RING 泛素連接酶MDM2抑制,而MDM2可通過泛素-蛋白酶體途徑降解p53。也有報道表明,MDM2可作為p53的NEDD8-E3連接酶[34],p53蛋白超家族中成員p73也能被NEDD化所抑制活性[35]。
在卵泡顆粒細胞中過表達與沉默bta-miR-1343-3p后檢測細胞的增殖情況,MTT結果表明,bta-miR-1343-3p的過表達會抑制顆粒細胞的增殖,而bta-miR-1343-3p沉默會促進顆粒細胞增殖。這一結果表明,在卵泡顆粒細胞中bta-miR-1343-3p可通過抑制NEDD8的表達促進顆粒細胞的凋亡。這與其他研究中報道的NEDD8誘導細胞凋亡、抑制腫瘤細胞的生長的結果一致[36-37]。進一步在bta-miR-1343-3p過表達和抑制的細胞中檢測CDK1與CDK4基因的表達量,結果發現,過表達bta-miR-1343-3p時細胞周期相關基因表達量顯著下降,抑制bta-miR-1343-3p時表達量顯著上升。細胞凋亡相關基因Bax則在過表達bta-miR-1343-3p后表達量顯著升高,Fas基因表達量有所升高但差異不顯著。上述結果可與之前的MTT結果相互印證,說明bta-miR-1343-3p通過靶向調節NEDD8基因的表達抑制了卵泡顆粒細胞的增殖。
綜上所述,bta-miR-1343-3p靶向的NEDD8基因調控了牛卵泡顆粒細胞的增殖與凋亡,對卵泡發育起著重要的作用。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
