江西鉛山紅芽芋蔗糖合成酶基因的克隆和表達分析
2023-02-17 13:38:42張藝欣符卉卉顧佳怡蘭佳欣
江蘇農業科學 2023年2期
張藝欣, 段 菲, 樊 潔, 符卉卉, 顧佳怡, 蘭佳欣, 李 嬌
(1.上饒師范學院生命科學學院,江西上饒 334001; 2.上饒農業技術創新研究院,江西上饒 334001;3.上饒市藥食同源植物資源保護與利用重點實驗室,江西上饒 334001;4.上饒市薯芋類作物種質保存與利用重點實驗室,江西上饒 334001)
江西鉛山紅芽芋(ColocasiaesculentaL. Schoot var.cormosus‘Hongyayu’)為天南星科芋屬多子芋類型草本宿根植物,為江西省鉛山縣“兩紅”(紅芽芋和河紅茶)產業之一,2013年4月15日被核準為國家地理標志農產品,其肉白芽紅,營養豐富,肉質細嫩,口感細松滑,糯香可口,是藥食兼優、老幼皆宜的上好滋補佳品[1-2]。當植物處于逆境時,適度積累蔗糖可保證細胞質膜的穩定性,避免胞內外相關蛋白的降解,還可作為逆境脅迫下植物代謝所需能量的來源[3]。作為蔗糖代謝過程中的關鍵酶,蔗糖合成酶主要負責植物組織細胞壁成分或淀粉合成的調控,直接參與植物的生長發育過程[4]。因此,克隆江西鉛山紅芽芋蔗糖合成酶基因并檢測其組織表達特異性,對于了解江西鉛山紅芽芋糖類代謝及其品質形成具有重要意義。目前,對紅芽芋的研究主要集中在營養成分[5]、品質選育[6]、種植栽培[7]、組織培養[8]等方面。紅芽芋的淀粉含量較高,且淀粉顆粒細小,易被消化[5]。大量研究發現,蔗糖合成酶的功能主要是參與淀粉的合成[9]。目前,關于紅芽芋蔗糖合成酶基因同源克隆和表達特性分析的研究還未見報道,關于紅芽芋蔗糖合成酶cDNA序列及其在紅芽芋不同組織器官中如何表達、如何參與紅芽芋淀粉的合成等問題尚待解決。本研究通過江西鉛山紅芽芋試管苗轉錄組數據庫篩選到江西鉛山紅芽芋蔗糖合成酶基因的核心片段,通過設計簡并引物,用RT-PCR方法克隆江西鉛山紅芽芋蔗糖合成酶的編碼基因,并對其進行氨基酸序列比對、蛋白質二級結構和三級結構預測、系統進化樹分析,利用實時熒光定量PCR技術分析蔗糖合成酶編碼基因在江西鉛山紅芽芋不同器官(根、莖和葉)、不同球莖膨大時期(初期、中期和末期)的表達情況,為進一步研究蔗糖合成酶基因功能及代謝途徑奠定理論基礎,同時為江西鉛山紅芽芋蔗糖代謝和生長發育提供較為豐富有效的科學理論依據。
1 材料與方法
1.1 試驗材料
江西鉛山紅芽芋單芽在MS液體培養基中培養2個月,形成球莖膨大初期的完整試管苗;在MS液體培養基培養4個月,形成球莖膨大中期的完整試管苗;在MS液體培養基培養6個月,形成球莖膨大末期的完整試管苗。
1.2 試驗方法
1.2.1 總RNA的提取和cDNA第1鏈的合成 2020年8月,參試劑盒說明書,用TRIzol提取江西鉛山紅芽芋試管苗的總RNA,RNA濃度和完整性用紫外分光光度計和瓊脂糖凝膠電泳進行檢測。以提取的RNA作為模版,參照試劑盒(M-MLV cDNA第1鏈的合成)說明書進行cDNA第1鏈的合成。以Oligo(dT)18 Primer作為逆轉錄引物(引物序列:5′-G G C C A C G C G T C G A C T A G T A C T T T T T T T T T T T T T T T T T T-3′)。
1.2.2 蔗糖合成酶基因的克隆 江西鉛山紅芽芋蔗糖合成酶的轉錄組Unigene序列信息為TRINITY_DN8401_c0_g1,引物用Primer Premier 5.0軟件設計,F:5′-A T G C C G G A A C G C C A G C T G A-3′;R:5′-T C A C T T C T C A T A G A T A C G C T T C A G G-3′。PCR擴增條件:95 ℃ 2 min;95 ℃ 30 s,57.5 ℃ 30 s,72 ℃ 30 s,35個循環;72 ℃ 10 min。用1%瓊脂糖凝膠電泳對PCR產物進行檢測后,將含有目的基因的條帶與pMD19-T載體連接,并用熱激法將其轉化到感受態細胞EscherichiacoliDH5α中,對鑒定正確的陽性轉化子提取質粒后,送至生工生物工程(上海)股份有限公司進行測序。
1.2.3 蔗糖合成酶基因的生物信息學分析 用BioEdit軟件將基因序列翻譯為氨基酸序列,用 ProtParam預測酶的理化性質,用ProtScale預測酶的疏/親水性,用GOR軟件在線預測酶的二級結構,用SWISS-MODEL在線預測酶的三級結構,用WoLF PSORT在線預測基因的表達部位。通過DNAMAN和BioEdit軟件進行氨基酸序列的比對,用MEGA 5.0軟件進行系統進化樹的構建。
1.2.4 蔗糖合成酶基因的組織表達分析 取在MS液體培養基中培養2個月的江西鉛山紅芽芋完整試管苗根、莖、葉以檢測蔗糖合成酶基因在不同器官中的表達情況;取江西鉛山紅芽芋在MS液體培養基中培養2個月(球莖膨大初期)、4個月(球莖膨大中期)和6個月(球莖膨大末期)的完整試管苗的試管球莖以檢測蔗糖合成酶基因在球莖膨大不同時期的表達情況。以上材料各提取得到500 ng RNA,將其反轉錄為cDNA。通過熒光定量PCR(qRT-PCR,SYBR Green I)檢測得出,其內參基因為GAPDH。設計引物(F:5′-G C C C C G T C T T A T T C A T A C C C T C G-3′;R:5′-C A T G C T T T T C C C A T T T T G T C C T C A-3′)。qRT-PCR檢測采用20 μL反應體系,PCR反應程序:95 ℃ 10 min;95 ℃ 10 s,60 ℃ 34 s,95 ℃ 15 s,40個循環。用2-ΔΔCT法計算基因的表達水平。試驗重復3次,所有數據均表示為平均值±標準差,并用SPSS 19.0軟件進行統計分析,用單因素方差分析(One-way ANOVA)檢驗蔗糖合成酶基因組織表達的差異顯著性(α=0.05)。
2 結果與分析
2.1 江西鉛山紅芽芋蔗糖合成酶基因的cDNA序列
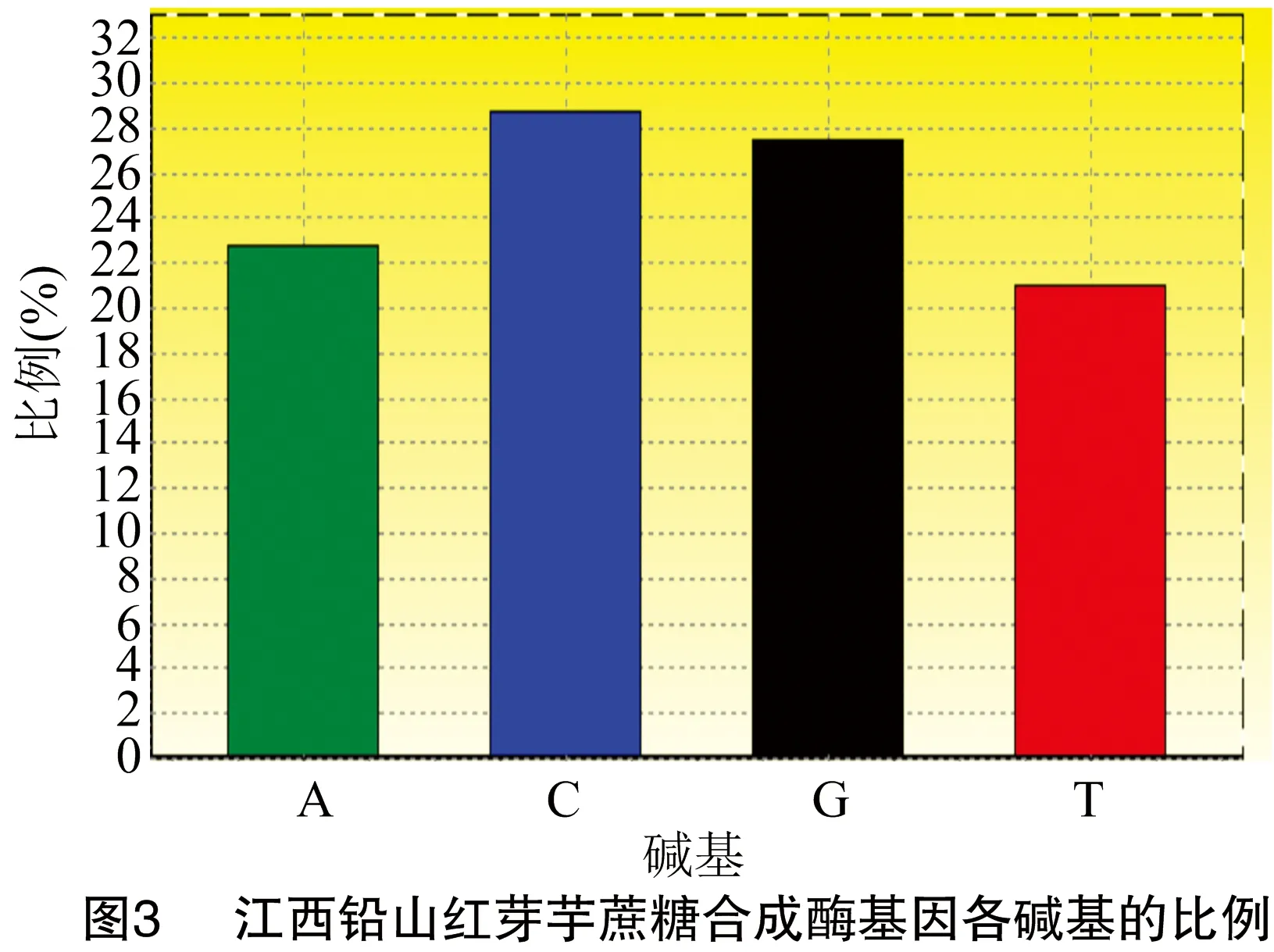
對江西鉛山紅芽芋蔗糖合成酶基因進行PCR擴增,結果見圖1。江西鉛山紅芽芋蔗糖合成酶基因cDNA總長度為2256bp(圖2),G+C含量為56.21%(圖3)。

2.2 江西鉛山紅芽芋蔗糖合成酶的氨基酸序列
通過ProtParam預測,得出江西鉛山紅芽芋蔗糖合成酶的氨基酸序列(圖4)。江西鉛山紅芽芋蔗糖合成酶由751個氨基酸組成,相對分子量為 85 350.64 u,等電點為5.83,為親水性蛋白。各氨基酸的數目和占比如下:丙氨酸(Ala,A),47個,占比6.3%;精氨酸(Arg,R),44個,占比5.9%;天冬酰胺(Asn,N),32個,占比4.3%;天冬氨酸(Asp,D),40個,占比5.3%;半胱氨酸(Cys,C),10個,占比1.3%; 谷氨酰胺(Gln,Q),26個,占比3.5%;谷氨酸(Glu,E),60個,占比8.0%;甘氨酸(Gly,G),51個,占比6.8%;組氨酸(His,H),29個,占比3.9%;異亮氨酸(Ile,I),41個,占比5.5%;亮氨酸(Leu,L),86個,占比11.5%;賴氨酸(Lys,K),36個,占比4.8%;甲硫氨酸(Met,M),16個,占比2.1%;苯丙氨酸(Phe,F),36個,占比4.8%;脯氨酸(Pro,P),37個,占比4.9%;絲氨酸(Ser,S),38個,占比5.1%;蘇氨酸(Thr,T),32個,占比4.3%;色氨酸(Trp,W),8個,占比1.1%;酪氨酸(Tyr,Y),27個,占比3.6%;纈氨酸(Val,V),55個, 占比7.3%。帶負電殘基總數(Asp+Glu)為100個,正電荷殘基(Arg+Lys)總數為80個。估計半衰期:30 h(哺乳動物網織紅細胞,體外)>20 h(酵母,體內)>10 h(大腸桿菌,體內)。經計算, 江西鉛山紅芽芋蔗糖合成酶失穩指數(Ⅱ)為35.94,表明其在分類上屬于穩定蛋白。


2.3 江西鉛山紅芽芋蔗糖合成酶親疏水性的分析
圖5中高峰值(正值)區域表示疏水區域,而負值的“低谷”區域是親水區域。疏水性結果顯示,最大疏水值為2.5左右,說明在該多肽中該處的疏水性最強;親水峰最大值為-2.5左右,整個蛋白質表現出高度的親水性,說明該蛋白為親水性蛋白。

2.4 江西鉛山紅芽芋蔗糖合成酶二級結構的分析
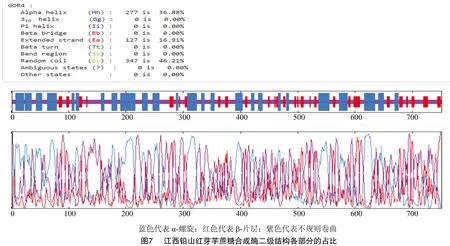
江西鉛山紅芽芋蔗糖合成酶的二級結構(圖6)預測如下:根據GOR預測結果,其二級結構由α螺旋(alpha helix,占比36.88%)、β-片層(extended strand,占比16.91%)、無規則卷曲(random coil,占比46.21%)構成(圖7)。從分布位點上來看,C端主要含有無規則卷曲和α-螺旋,N端主要含有無規則卷曲和β-片層,無規則卷曲、β-片層和 α-螺旋散布于整個蛋白質中。

2.5 江西鉛山紅芽芋蔗糖合成酶三級結構的分析
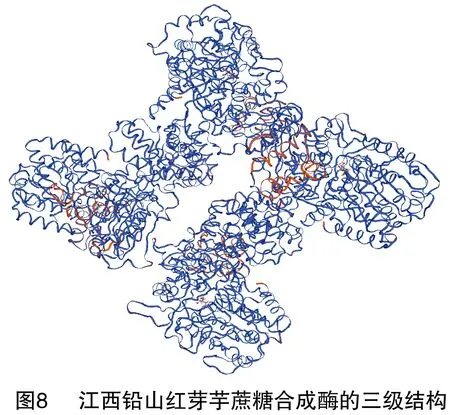
蛋白質的三級結構是指整條多肽鏈的三維空間結構,也就是包括碳骨架、側鏈在內的所有原子的空間排列。SWISS-MODEL是一款用同源建模法預測蛋白質三級結構的全自動在線預測軟件,共有3個工作方式:First Approach mode、Alignment Interface mode、Project(Optimise)mode。SWISS-MODEL預測結果顯示,江西鉛山紅芽芋蔗糖合成酶的三級結構為同源四聚體,詳見圖8。
2.6 江西鉛山紅芽芋蔗糖合成酶的亞細胞定位
用WoLF PSORT在線軟件對江西鉛山紅芽芋蔗糖合成酶基因的表達部位進行預測,圖9結果顯示,定位于細胞質中的蔗糖合成酶基因數量為6個,定位于線粒體中的蔗糖合成酶基因數量為4.5個,定位于細胞質_線粒體中的蔗糖合成酶基因數量為3個,定位于葉綠體中的蔗糖合成酶基因數量為1個,定位于細胞骨架_質膜中的蔗糖合成酶基因數量為1個,表明江西鉛山紅芽芋蔗糖合成酶基因主要存在細胞質中。
2.7 江西鉛山紅芽芋蔗糖合成酶的系統進化分析
從構建的進化樹(圖10)可以看出,江西鉛山紅芽芋與芋(Colocasiaesculenta)、眼子菜(Potamogetondistinctus)在一個大分支下,說明江西鉛山紅芽芋蔗糖合成酶在進化上與C.esculentahypothetical protein Taro_012658(MQL80204_c0_g1)、P.distinctussucrose synthase(BAE06059.1)的親緣關系較近,特別是與C.esculentahypothetical protein Taro_012658(MQL80204_c0_g1)具有最高的親緣關系。


2.8 江西鉛山紅芽芋蔗糖合成酶同源蛋白序列的比對結果
江西鉛山紅芽芋蔗糖合成酶同源蛋白的序列比對信息見圖11,其中的“※”號區域是該蛋白家族的保守結構域。由圖11可知,江西鉛山紅芽芋與芋(Colocasiaesculenta)、眼子菜(Potamogetondistinctus)的同源性較高,再次說明江西鉛山紅芽芋蔗糖合成酶在進化上與C.esculentahypothetical protein Taro_012658(MQL80204_c0_g1)、P.distinctussucrose synthase(BAE06059.1)的親緣關系較近,特別是與C.esculentahypothetical protein Taro_012658(MQL80204_c0_g1)具有最高的親緣關系。
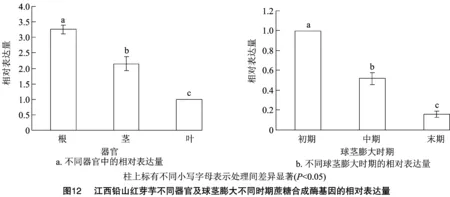
2.9 江西鉛山紅芽芋不同器官及球莖膨大不同時期蔗糖合成酶基因的表達分析
以江西鉛山紅芽芋的GAPDH為內參,利用實時熒光定量PCR分析江西鉛山紅芽芋蔗糖合成酶基因在江西鉛山紅芽芋不同器官中的表達情況。結果顯示,江西鉛山紅芽芋蔗糖合成酶基因在根、莖、葉中均有表達,但在不同組織器官中的表達情況差異顯著(圖12),其中江西鉛山紅芽芋蔗糖合成酶基因在根、球莖膨大初期的相對表達量最高。
3 討論
植物光合作用的主要產物為蔗糖,植物經光合作用形成蔗糖后通過葉片源源不斷地向庫器官(塊根、塊莖、球莖、鱗莖等)輸送,到達庫器官后直接或間接參與其生理代謝進程,從而進一步影響植株的生長發育和產量調控[10]。蔗糖到達庫器官后不能直接被細胞吸收利用,必須依靠蔗糖轉化酶或蔗糖合成酶的作用將其分解后才能參與植物的代謝活動[11]。因此,蔗糖合成酶是蔗糖代謝的關鍵調節酶,主要影響植物細胞之間的碳水化合物分配及碳水化合物的結構組成和貯藏[12]。蔗糖合成酶分解蔗糖活性的強弱與植物淀粉貯藏器官庫強度密切相關,包括植物的變態根、變態莖等[13]。在江西鉛山紅芽芋中,蔗糖是葉片光合碳水化合物轉運和分配的主要形式,是江西鉛山紅芽芋球莖淀粉合成的初始原料。本試驗利用同源克隆技術成功克隆了江西鉛山紅芽芋蔗糖合成酶基因序列,該基因cDNA總長度為2 256 bp,G+C含量為56.21%;江西鉛山紅芽芋蔗糖合成酶由751個氨基酸組成,分子量為85 350.64 u,等電點為5.83,為親水性蛋白;江西鉛山紅芽芋蔗糖合成酶二級結構由α-螺旋(36.88%)、β-片層(16.91%)、無規則卷曲(46.21%)構成,三級結構為同源四聚體;江西鉛山紅芽芋蔗糖合成酶主要存在于細胞質、線粒體中,在進化上與C.esculenta、P.distinctus的親緣關系較近,尤其是與C.esculentahypothetical protein Taro_012658(MQL80204_c0_g1)在進化上具有最高的親緣關系。
研究發現,蔗糖合成酶基因在植物中具有組織特異性和時間特異性。白及蔗糖合成酶基因的實時定量PCR結果表明,蔗糖合成酶基因在葉片中的表達量最高,在塊莖中的表達量最低;成熟葉片的表達量高于未成熟葉片的表達量[14]。半定量RT-PCR分析結果表明,蔗糖合成酶基因HbSS1、HbSS2在巴西橡膠樹各組織器官中均有表達,其中蔗糖合成酶基因HbSS1在葉中的表達量最高,蔗糖合成酶基因HbSS2在樹皮中的表達量最高[15]。蔗糖合成酶基因組織表達特異性分析結果顯示,SoSuS1在甘蔗熱帶種Badila莖、葉鞘、葉片中均有表達,且SoSuS1在甘蔗熱帶種Badila莖中的表達量最高[16]。實時定量PCR表達分析結果顯示,蔗糖合成酶LbSusy基因在枸杞莖中的表達量最高,在根中的表達水平較低[17]。半定量RT-PCR表達分析結果表明,蔗糖合成酶基因PhSUS在蝴蝶蘭的組織中均有表達,在花蕾發育中后期、成熟花和花器官的表達量都比營養階段根、葉中的表達量高[18]。蔗糖合成酶ZmSus6、ZmSus7基因在甜玉米的根、莖、葉、玉米芯和籽粒中均有表達,但以葉、根中的相對表達量最高[19]。鐵皮石斛葉、莖、根蔗糖合成酶的半定量RT-PCR檢測結果表明,蔗糖合成酶基因在鐵皮石斛植株中不同部位表達量不同,其表達模式表現為莖>葉>根[20]。玉米蔗糖合成酶的組織表達特異性分析發現,蔗糖合成酶基因ZmSUS1和ZmSUS2在胚和胚乳中大量表達,蔗糖合成酶基因ZmSUS5在所有被檢測組織中的表達量很低,蔗糖合成酶基因ZmSUS3及ZmSUS4在被檢測的組織中均有較高的表達量[21]。本試驗結果與上述結果較為類似。本試驗的實時定量PCR結果顯示,蔗糖合成酶基因在江西鉛山紅芽芋的表達存在器官特異性,在根、球莖膨大初期中的表達量最高。因此,江西鉛山紅芽芋蔗糖合成酶基因的克隆不僅為研究該基因在江西鉛山紅芽芋中的表達調控奠定了基礎,而且豐富了植物蔗糖合成酶分子進化和結構功能研究的材料。