油菜糖基轉移酶BnIRX14基因家族鑒定及遺傳轉化
2023-02-21 08:04:44靳豐蔚楊曉明劉婷婷
西北植物學報 2023年1期
靳豐蔚,董 云*,楊曉明,方 彥,劉婷婷,王 毅
(1 甘肅省農業科學院作物研究所,蘭州 730070;2 甘肅農業大學干旱生境作物學重點實驗室,蘭州 730070)
糖基轉移酶是多糖合成中的關鍵酶,參與多種物質的糖基化,催化糖供體和受體結合產生新的化合物,其供體多是激活狀態的核苷二磷酸糖,如UDP-葡萄糖、GDP-甘露糖和UDP-半乳糖等[1]。植物體內存在大量的天然次生代謝物,這些次生代謝物的產生是植物在進化過程中對生態環境適應的結果,是植物生理狀態在代謝水平上的反映[2]。糖基化是這些次生代謝產物的主要修飾之一,糖基化修飾是植物體內普遍存在的生理生化反應,可以提高天然化合物的溶解度和穩定性,使化合物便于儲存在植物細胞中,是調節細胞代謝平衡的重要機制[3];糖基化修飾可以改變化合物毒性和生物活性[4],在植物響應逆境脅迫等方面具有重要作用[5]。
根據序列同源性和催化機制,在碳水化合物活性酶數據庫CAZy(Carbohydrate-Active Enzymes database) 中,根據GT序列相似度、催化底物的特異性和催化產物化學結構,將糖基轉移酶分為114個家族。根據糖基轉移酶三維結構的折疊特征,將糖基轉移酶被分為GT-A、GT-B、GT-C和GT-D等4種類型[5]。糖基轉移酶家族多使用NDP-戊糖和NDP-己糖作為供體底物,催化UDP-糖的糖基轉移酶稱為UDP-糖基轉移酶(UGTs)[6]。
關于UGTs的研究,有學者分別在蕓薹屬植物白菜(Brassicarapa)、甘藍型油菜(Brassicanapus)和甘藍(Brassicaoleracea)中鑒定到140、251和154個UGTs[7],這些糖基轉移酶主要參與合成次生代謝物質響應生物非生物脅迫。水稻超表達OsUGT90A1能保護冷脅迫下細胞膜的完整性[8];而有學者研究UGT84A17能夠調控楊樹對逆境做出積極響應[9];擬南芥UGT85U1/2和UGT85V1通過改變吲哚衍生物參與應答鹽和活性氧脅迫抗性[10];甘藍型油菜UGT84F1和白菜BrGUGT也被發現通過抑制芥子酸來適應非生物脅迫[11]。
木聚糖是存在于植物細胞壁中的一種異質多糖,是植物半纖維素的主要成分。模式植物擬南芥遺傳和生物化學分析表明,一些來自糖基轉移酶GT43家族的BnIRX14 (IRX9I9H/IRX9-L、IRX14、I14H/IRX14-L)和GT47(IRX10、IRX10-L) 家族基因是木糖鍵延伸所必須的。木糖基轉移酶除了參與木聚糖的合成,還在擬南芥[12]和南荻(Miscanthuslutarioriparius)[13]種皮粘液合成中起作用,糖基轉移酶生物功能具有多樣性和保守性,影響植物的生長發育、逆境應答等功能[14]。油菜是中國重要的油料作物,次生物質代謝和抗逆機理解析是油菜育種的重要基礎研究。目前糖基轉移酶調控的油菜抗性研究還相對滯后。甘肅省農業科學院作物研究所前期克隆了一個與甘藍型油菜抗性有關的β-1,4-木糖基轉移酶基因BnIRX14(NCBI登錄號KP100641),發現該基因在細胞質中表達,屬非分泌性蛋白[15]。本研究利用生物信息學方法對甘藍型油菜BnIRX14家族成員進行鑒定,從基因組水平上分析了甘藍型油菜BnIRX14基因數目、進化和結構以及在染色體上的分布,并構建BnIRX14的干擾載體轉化油菜,為進一步發掘BnIRX14基因在油菜抗性育種中的利用價值提供基礎。
1 材料和方法
1.1 材 料
1.1.1 油菜品種供試材料為甘藍型油菜(BrassicanapusL.)春性品種Westar,由華中農業大學生命科學學院提供。
1.1.2 其他材料農桿菌GV3101、大腸桿菌(Escherichiacoli)感受態細胞DH5α、植物RNAi載體pKANNIBAL、植物表達載體pART27以及Taq DNA聚合酶2×fast pfu master mix、限制性內切酶XhoⅠ、KpnⅠ、XbaⅠ、HindⅢ、NotⅠ,T4連接酶,質粒提取試劑盒Plasmid Mini Kit Ⅰ(OMEGA),基因組提取試劑盒Trans Direct Plant Tissue PCR Ki均購自陜西博瑞德生物科技公司。其他化學藥品均為國產分析純試劑。
1.2 方 法
1.2.1 油菜BnIRX14基因家族成員鑒定根據BnIRX14氨基酸序列,利用Clustal W 默認設置進行多重序列比對,并在MEGA 6 軟件中使用鄰接法(Neighbour-Joining,NJ) 構建系統進化樹[16]。利用GSDS2.0繪制BnIRX14基因結構[17]。通過MEME分析BnIRX14蛋白保守元件[18]。
1.2.2 油菜BnIRX14基因干擾載體構建利用Primer Premier 6.0軟件設計BnIRX14基因靶位點序列擴增引物(表 1),擴增后用百泰克膠回收試劑盒回收目的片段,回收后置-20 ℃保存備用。用XhoⅠ和KpnⅠ雙酶切質粒pKANNIBAL載體,切取載體大片段對應條帶的凝膠,百泰克膠回收試劑盒回收酶切產物。將膠回收的正向片段和線性化pKANNIBAL載體以9∶1摩爾比加到EP管中進行重組反應,混勻后在37 ℃放置30 min進行轉化[19]。進行單菌落PCR陽性克隆鑒定,挑取陽性菌落測序驗證。將測序正確的質粒用XbaⅠ和HindⅢ雙酶切pKANNIBAL正向片段載體,回收大片段。

表1 本實驗所用引物Table 1 Primers in this test
將膠回收的反向片段和線性化pKANNIBAL正向片段載體以9∶1摩爾比加到EP管中進行重組反應,連接、轉化、鑒定方法同正向片段。將測序正確的含有正反向片段的pKANNIBAL載體和表達載體pART27用NotⅠ酶切,T4連接酶連接(圖 1),轉化至DH5α感受態細胞,菌落PCR進行陽性克隆鑒定。

圖1 干擾載體pART27-485-6構建Fig.1 The structure of interference vector pART27-485-6
利用Plasmid Mini Kit Ⅰ提取測序正確的pART27-485-6質粒轉化農桿菌GV3101,轉化參照董云等[19],待單菌落長出后,選3~5個單菌落做菌落PCR驗證,引物見表1,PCR反應體系和條件與靶位點序列擴增相同。
1.2.3 油菜BnIRX14基因干擾載體轉化參照劉宏波[20]以及CARDOZAD等[21]、DE BLOCK等[22]和MAHESHWARI等[23]關于農桿菌介導的轉基因方法。以栽培品種Westar為受體材料,用轉化了pART27-485-6質粒的農桿菌GV3101侵染油菜下胚軸,當抗性芽在誘導分化培養基上長到2 cm時,切芽并將其轉入生根培養基中,待形成完整小植株時切取葉片,使用Trans Direct Plant Tissue PCR Kit試劑盒提取基因組DNA,用4對基因特異引物(表 1)檢測陽性植株。參照方彥等[24]方法,依據BnIRX14基因的cDNA全長序列設計特異RT-PCR引物,Actin作為內參基因對陽性植株的花、角果、葉片進行實時熒光定量PCR分析,實時熒光定量PCR引物見表 1。
2 結果與分析
2.1 油菜BnIRX14基因家族成員鑒定
根據前期獲得的BnIRX14基因序列[15],在PFAM數據庫中預測基因保守結構域,結果表明,BnIRX14包含一個糖基轉移酶家族結構域。根據保守結構域在甘藍型油菜基因組注釋序列中檢索家族基因,得到11個家族成員。
氨基酸序列系統進化樹分析表明,甘藍型油菜BnIRX14家族成員被分成3個亞家族(圖2,A),第一個亞家族包括IRX9-L-1、IRX9-L-2、IRX9-L-4、IRX9-L-5、IRX9-L-7、IRX9-L-8,第二個亞家族包括IRX9-L-3、IRX9-L-6,第三個亞家族包括IRX14-L-1、IRX14-L-2、IRX14-L-3。其中,有8個BnIRX14基因家族成員分別分布在6條染色體上(圖 2,B),分別為A07、A08、A09、C03、C07和C09染色體,除C07染色體有3個基因外,其余染色體均有1個基因,根據染色體定位分布信息和序列比對分析,發現IRX14-L-2和IRX14-L-3串聯重復。第一亞家族基因有3~4個外顯子,第二亞家族基因有7~9個外顯子,第三亞家族基因有2~4個外顯子,IRX14-L-1只有2個外顯子,且內含子最短(圖 3,A)。
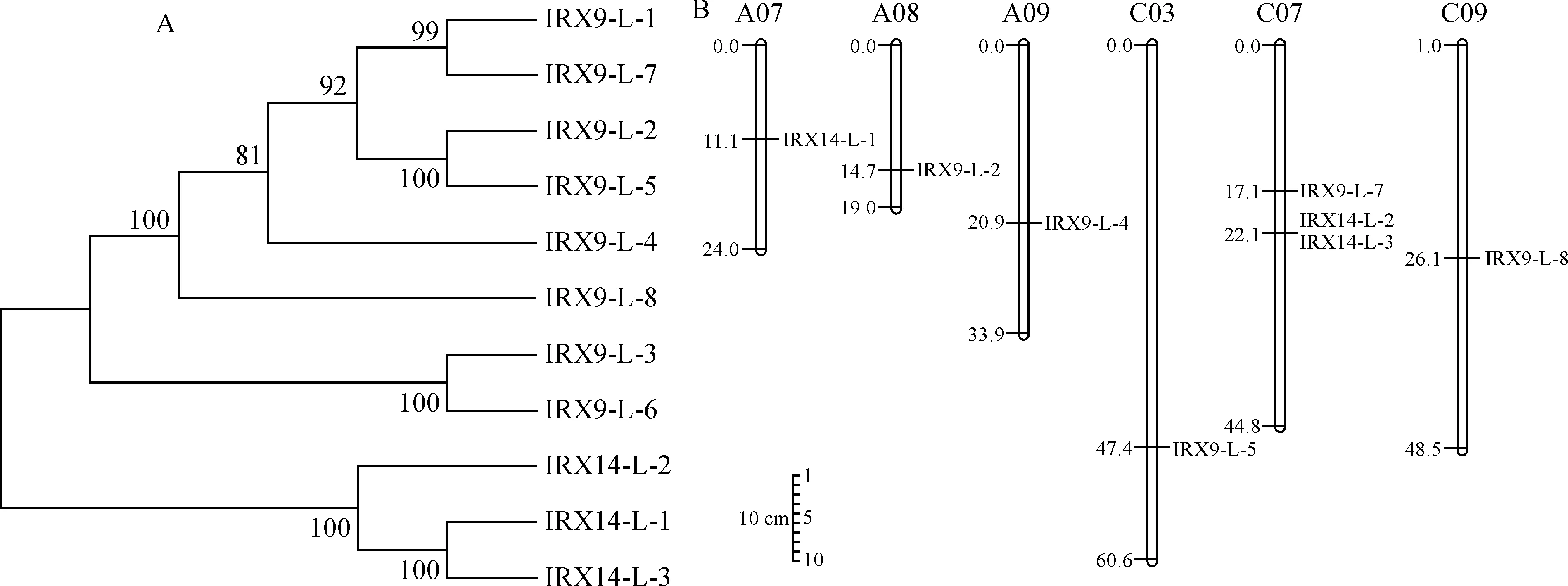
圖2 BnIRX14家族氨基酸序列系統進化樹(A)和染色體分布(B)Fig.2 Phylogenetic tree of amino acid sequences (A) and chromosome distribution (B) of BnIRX14 family members
在BnIRX14蛋白中預測出10個保守元件(圖 3,B),第一亞家族蛋白有2~8個保守元件,其中:IRX9-L-4有2個,IRX9-L-8有5個,其余4個蛋白均有8個相同的元件;第二亞家族蛋白只有1個保守元件;第三亞家族蛋白有1~4個保守元件,其中IRX14-L-2僅1個保守元件,IRX14-L-1和IRX14-L-3存在4個相同元件。元件1除IRX9-L-4和第三亞家族外,在其他兩個亞家族蛋白中均存在;元件2在第一、第三亞家族蛋白中存在;元件3僅在第一亞家族蛋白中存在;元件9、10只有IRX14-L-1、IRX14-L-3存在。3個亞家族在基因結構和保守元件中具有較大特異性。
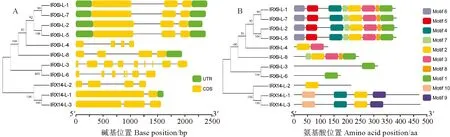
圖3 BnIRX14基因家族結構(A)和蛋白保守元件(B)Fig.3 Family structure (A) and protein conserved element (B) of BnIRX14
2.2 油菜BnIRX14基因干擾載體構建
利用BnIRX14基因靶位點序列設計的正向片段擴增引物F485-6-F/F485-6-R和反向片段擴增引物R485-6-F/R485-6-R進行PCR擴增,擴增產物分別為340和336 bp(圖4,A),膠回收目的片段。XhoⅠ和KpnⅠ雙酶切線性化質粒pKANNIBAL載體,與膠回收的正向片段進行重組后轉化大腸桿菌,用正向片段擴增引物進行單菌落PCR鑒定得到4個陽性克隆(圖4,B)。
XbaⅠ和HindⅢ 雙酶切測序正確的質粒pKANNIBAL 正向片段載體,與膠回收的反向片段進行重組轉化大腸桿菌,用反向片段擴增引物進行單菌落PCR鑒定得到2個陽性克隆(圖4,C)。NotⅠ酶切含有正反向片段的pKANNIBAL載體和植物表達載體pART27,膠回收目的片段后用T4連接酶連接,轉化至DH5α感受態細胞,用反向片段擴增引物單菌落PCR鑒定得到1個陽性克隆(圖4,D)。提取測序正確的pART27-485-6質粒轉化農桿菌GV3101,利用485F/485R進行單菌落PCR,擴增條帶743 bp,驗證得到4個陽性克隆(圖4,E)。

A.正反向片段PCR擴增:1、2.F485-6;3、4.R485-6;B.F485-6連接載體pKANNIBAL菌落PCR鑒定:1-4.pKANNIBAL-F485-6;C.R485-6連接載體pKANNIBAL-F485-6菌落PCR鑒定:1、2.pKANNIBAL- F485-6- R485-6;D.pART27-485-6轉大腸桿菌菌落PCR鑒定;E.pART27-485-6轉農桿菌菌落PCR鑒定;M.DL2000圖4 BnIRX14基因干擾表達載體構建和農桿菌轉化PCR鑒定A.PCR amplification of positive and reverse fragments:1 and 2 were positive fragments F485-6,3 and 4 were reverse fragments R485-6;B.Positive colony PCR identification of the forward fragment F485-6 linked to the vector pKANNIBAL:1-4 were pKANNIBAL-F485-6;C.Positive colony PCR identification of the reverse fragment R485-6 linked to the vector pKANNIBAL-F485-6,1 and 2 were pKANNIBAL-F485-6-R485-6;D.Colony PCR identification of plasmid pART27-485-6 transfected Ecoli;E.Colony PCR identification of plasmid pART27-485-6 transfected agrobacterium tumefaciens;M.DL2000Fig.4 PCR identification of BnIRX14 gene interference expression vector and agrobacterium transgenics
2.3 油菜BnIRX14基因RNA干擾表達載體遺傳轉化和表型分析
2.3.1 遺傳轉化通過農桿菌介導侵染油菜下胚軸進行BnIRX14基因干擾表達載體 pART27-485-6的遺傳轉化,侵染油菜下胚軸300個(圖5,A),出愈率67%,分化培養和Kan抗性篩選,獲得抗性芽60個(圖5,B),通過生根培養得到再生植株20個(圖5,C),用4對特異性引物JDF1/JDR1、JDF2/JDR2、JDF3/JDR3和JDF4/JDR4進行PCR檢測后,擴增條帶分別為1 500、1 500、1 000和1 000 bp(圖5,D~G),確定5株為陽性轉化體。

A.GV3101侵染油菜下胚軸共培養;B.誘導分化培養;C.生根培養;D-G.PCR鑒定:M.DL5000;2#、5#、6#、9#、10#。陽性轉化體;N.陰性對照圖5 干擾表達載體 pART27-485-6的遺傳轉化及PCR鑒定A.Coculture of rapeseed hypocotyls infected by agrobacterium Gv3101;B.Resistant buds grown on induced differentiation medium;C.Resistant buds grown in rooting medium;D-G.PCR identification of positive plants:2,5,6,9 and 10# were transformation events;M.DL5000;N.Negative controlFig.5 Genetic transformation of expression vector pART27-485-6 and PCR identification
2.3.2 表型分析通過BnIRX14基因在不同器官中表達量的分析(圖 6)在油菜5個轉化干擾表達載體 pART27-485-6陽性植株的花、角果、葉中,BnIRX14基因表達量均低于野生型,且差異顯著,說明BnIRX14基因在花、角果和葉中的表達受到抑制。BnIRX14基因在野生型油菜的花、角果、葉片中的表達量基本一致,但在油菜轉化干擾表達載體pART27-485-6陽性植株的花、角果、葉片中的表達量是依次降低。表型鑒定發現,編號為9#和10#的陽性轉化株花的柱頭至花柱中央為一孔狀的空腔,子房較野生型明顯膨大,在柱頭表面授粉后,不能結實,表現雌性不育(圖7),其他3個陽性株花器結構發育正常,但植株莖、枝表皮有液體滲出,呈露珠狀粘附在莖、枝表面。

WT.野生型;2#、5#、6#、9#、10#.陽性株系;下同。小寫字母表示差異顯著(P≤0.05)圖6 BnIRX14基因在陽性植株不同器官中的表達WT.Wild type;2,5,6,9 and 10#.The positive plant;The same as below.Different normal letters meam significant difference at 0.05 level (P≤0.05)Fig.6 Expression of BnIRX14 gene in different organs of positive plants

A.株系;B.株系花蕾、花器和角果;C.WT柱頭;D.9#陽性轉化株柱頭圖7 轉化干擾表達載體陽性株雌性不育表型A.Strains;B.Flower bud,floral apparatus,silique;C.Chapiter of WT;D.Chapiter of positive strain 9#Fig.7 Female sterility phenotype of rape positive strains of the expression vector transferred
3 討 論
木聚糖是一種存在于植物細胞壁中的異質多糖,約占植物細胞干重的15%~35%,是植物半纖維素的主要成分[25]。β-1,4-木糖基轉移酶是糖基轉移酶家族GT43成員[26],大多以UDP-木糖作為專一性的糖供體,GT43家族中所含成員多與植物的次級代謝和逆境脅迫相關[27]。糖基轉移酶GT43家族成員在模式植物中有許多報道,像擬南芥IRX9、IRX9L、IRX14、IRX14L等[28]都參與了木糖的合成。木聚糖糖基轉移酶在植物生長發育、調節次生代謝及參與相應逆境脅迫有重要意義[29]。有學者從南荻(Miscanthuslutarioriparius)中鑒定到了7個GT43成員[13],在水稻中發現了10個GT43成員[30],這些功能基因冗余具有多樣性。
董云通過RACE技術從甘藍型油菜中克隆到了β-1,4-木糖基轉移酶基因BnIRX14,保守結構域和亞細胞定位表明BnIRX14屬于油菜糖基轉移酶GT43家族成員[15]。本研究通過同源序列比對,在甘藍型油菜中檢索到了11個BnIRX14基因家族成員,分別定位在A07、A08、A09、C03、C07和C09染色體上,其中IRX14-L-2和IRX14-L-3串聯重復,可能來自于遠緣雜交的甘藍基因組。IRX9-L-1、IRX9-L-7、IRX9-L-2和IRX9-L-5結構域十分保守,都存在8個保守的結構域,進化樹結果也表明IRX9-L-1和IRX9-L-7,IRX9-L-2和IRX9-L-5分別聚為一類,說明在進化上的保守性。
通過農桿菌介導的遺傳轉化方法,構建了油菜BnIRX14干擾表達載體轉化油菜下胚軸,通過生根培養得到轉化株系20個。qRT-PCR檢測T0代陽性轉基因油菜,相比野生型BnIRX14在花,角果和葉中均下調表達,其中葉中下調最為明顯,可能的原因在于BnIRX14參與木糖調節的半纖維素的合成,而葉中相比花和角果中本底水平的BnIRX14表達量就低,導致干擾BnIRX14轉基因株系中BnIRX14基因低表達,但具體的原因還需要進一步分子水平的證據。
研究發現干擾BnIRX14轉基因株系9#和10#花器柱頭至花柱中央為一孔狀的空腔,子房較野生型明顯膨大,在柱頭表面授粉后,不能結實,表現雌性不育。其他2#、5#和6#陽性干擾BnIRX14轉基因株系花器結構發育正常,但植株莖、枝表皮有液體滲出,呈露珠狀粘附在莖、枝表面。原因可能是轉基因株系中基因被干擾的程度不同所致,這種情況在轉基因實驗中常常發現[31],不同的轉基因株系因對基因插入位點不同而造成基因表達量不同[32]。qRT-PCR檢測在9#和10#株系角果中BnIRX14基因表達低于其他轉基因干擾株系,也符合觀察到的結實發育的異常。2#、5#和6#轉基因株系植株莖、枝表皮有液體滲出,呈露珠狀粘附在莖枝表面的原因可能在于下調了BnIRX14基因的表達導致次生代謝物質的異常所致,但具體的分子機制和確鑿的證據正在進行后續的研究。本研究在甘藍型油菜中發現了11個BnIRX14基因家族成員,進化上高度保守且存在多樣性,功能缺失研究表明這些木糖基轉移酶家族基因能夠參與雌蕊的發育和次生物質代謝,該研究為油菜雜交育種和抗逆育種提供理論依據。
