水蕹菜和羅莎生菜對(duì)麥穗魚養(yǎng)殖系統(tǒng)水質(zhì)的影響
2023-02-26 06:02:20劉東姣杜新科王世興張冀野劉麗雪申旭紅
水產(chǎn)學(xué)雜志 2023年1期
關(guān)鍵詞:系統(tǒng)
劉東姣,杜新科,王世興,張冀野,劉麗雪,申旭紅
(1.中國(guó)農(nóng)業(yè)大學(xué)煙臺(tái)研究院,山東 煙臺(tái) 264670;2.山東省煙臺(tái)生態(tài)環(huán)境監(jiān)測(cè)中心,山東 煙臺(tái) 264010)
近年來(lái)隨著人民生活水平的提高,對(duì)水產(chǎn)品的需求不斷增加,水產(chǎn)養(yǎng)殖業(yè)總產(chǎn)值穩(wěn)步增長(zhǎng),預(yù)計(jì)到2030 年我國(guó)養(yǎng)殖魚類產(chǎn)量將占到世界魚類總量的62%[1]。但水產(chǎn)養(yǎng)殖的發(fā)展也引起了嚴(yán)重的環(huán)境問(wèn)題。傳統(tǒng)淡水養(yǎng)殖的殘餌及養(yǎng)殖動(dòng)物排泄物會(huì)造成水體氮含量過(guò)高,使水體富營(yíng)養(yǎng)化,影響?zhàn)B殖動(dòng)物健康[2]。生態(tài)可持續(xù)養(yǎng)殖是水產(chǎn)養(yǎng)殖的發(fā)展趨勢(shì)。
魚菜共生是將水產(chǎn)養(yǎng)殖與水耕栽培相結(jié)合的新型養(yǎng)殖模式,養(yǎng)殖水體中魚類排泄物、殘餌等物質(zhì)可轉(zhuǎn)化為蔬菜可吸收的養(yǎng)料,在收獲魚類及蔬菜的同時(shí),改善水環(huán)境污染問(wèn)題[3,4]。魚菜共生系統(tǒng)中植物對(duì)水體的凈化作用是實(shí)現(xiàn)生態(tài)可持續(xù)養(yǎng)殖的關(guān)鍵。目前對(duì)植物凈水已有大量研究,主要集中在石菖蒲(Acorus tatarinowii)、蝴蝶花(Iris japonica)、紫葉酢漿草(Oxalis triangularis)和鳳眼蓮(Eichhornia crassipes)等挺水植物以及番茄、青菜和芹菜等可食用蔬菜[5-9]。魚菜共生體系中對(duì)養(yǎng)殖動(dòng)物的選擇主要集中在可食用經(jīng)濟(jì)魚類,如鱸(Lateolabrax japonicus)、鯽(Carassius crassius)和鯉(Cyprinus carpio)[5-14]。研究發(fā)現(xiàn),不同的水培植物因種植密度、浮床覆蓋比例以及養(yǎng)殖水產(chǎn)品種類的不同,對(duì)氮的吸收效率也不同[5,6];構(gòu)建合理的魚菜共生養(yǎng)殖模式能夠顯著提高羅非魚的生長(zhǎng)速度、成活率和肥滿度[7]。
水蕹菜(Ipomoea aquatica)和羅莎生菜(Lactuca sativa)作為常見(jiàn)的可食用水培植物,已有在草魚(Ctenopharyngodon idella)和金鯽魚菜共生系統(tǒng)的相關(guān)研究[8,9]。水蕹菜和羅莎生菜生長(zhǎng)快,適應(yīng)能力強(qiáng),具有在魚菜共生系統(tǒng)中大規(guī)模應(yīng)用的廣闊前景。麥穗魚(Pseudorasbora parva)為鯉科麥穗魚屬的一種小型淡水魚,對(duì)水質(zhì)變化十分敏感,可作為水環(huán)境監(jiān)測(cè)的指示魚種[10]。隨著居民水產(chǎn)品消費(fèi)的多樣化,麥穗魚逐漸走上了餐桌,作為優(yōu)質(zhì)的觀賞魚餌料,麥穗魚人工養(yǎng)殖已成為一種趨勢(shì)。本試驗(yàn)通過(guò)建立水蕹菜-麥穗魚和羅莎生菜-麥穗魚的魚菜共生系統(tǒng),研究水蕹菜和羅莎生菜對(duì)麥穗魚養(yǎng)殖系統(tǒng)中氮的處理效果,對(duì)研究養(yǎng)殖系統(tǒng)水質(zhì)凈化具有重要意義,以期為進(jìn)一步完善魚菜共生系統(tǒng)的運(yùn)行機(jī)制和理論體系及養(yǎng)殖系統(tǒng)模式提供理論依據(jù)。
1 材料與方法
1.1 材料
麥穗魚捕自煙臺(tái)市牟平區(qū)野外池塘,隨后在實(shí)驗(yàn)室進(jìn)行消毒等處理。選取生長(zhǎng)狀況良好,體長(zhǎng)為(7.3±0.56)cm,體質(zhì)量為(3.88±0.78)g,規(guī)格相近的麥穗魚進(jìn)行試驗(yàn)。
水蕹菜和羅莎生菜購(gòu)于山東壽禾種業(yè),水蕹菜的初始株高為(26.1±1.84)cm,羅莎生菜的初始株高為(13.85±0.92)cm。
試驗(yàn)所用餌料由藍(lán)鯽飼料和拉絲蛋白粉(1∶0.2)配置,制成直徑約1.0 mm 的團(tuán)狀餌料。
1.2 方法
試驗(yàn)于中國(guó)農(nóng)業(yè)大學(xué)煙臺(tái)研究院實(shí)驗(yàn)室內(nèi)完成。麥穗魚、水蕹菜與羅莎生菜在正式試驗(yàn)前先分別進(jìn)行馴養(yǎng)和暫養(yǎng)。正式試驗(yàn)共6 周(42 d),設(shè)水蕹菜-麥穗魚和羅莎生菜-麥穗魚2 個(gè)處理組和1個(gè)單一養(yǎng)殖麥穗魚的對(duì)照組,每組設(shè)3 個(gè)重復(fù)。試驗(yàn)選用9 個(gè)泡沫箱(40 cm×24 cm×16 cm),放置在實(shí)驗(yàn)室靠窗處以保證充足的光照,每個(gè)箱中投放麥穗魚12 尾。搭建水蕹菜和羅莎生菜浮床,浮床面積占水體表面積35%,分別固定在泡沫保溫箱一側(cè),菜根裹海綿固定防止其松動(dòng),用紗布裹住菜根防止麥穗魚啃食。每日于早、中、晚定時(shí)投喂,及時(shí)補(bǔ)充因蒸發(fā)及取樣而減少的水量,保證水體體積恒定。
1.3 樣品收集與測(cè)定
自試驗(yàn)開始,每隔7 d 取一次水樣檢測(cè)水質(zhì),共取得6 組水樣數(shù)據(jù)。分別檢測(cè)了水體pH、溶氧(DO)、總氨氮(TAN)、亞硝酸鹽氮(NO2--N)、硝酸鹽氮(NO3--N)和總氮(TN)含量等指標(biāo),其中,pH 與DO 使用AZ-8403 便攜式多功能水質(zhì)檢測(cè)儀實(shí)時(shí)測(cè)得;TAN 濃度采用HJ 535-2009《水質(zhì)氨氮的測(cè)定納氏試劑分光光度法》測(cè)定;NO2--N 濃度采用GB/T 7493-1987《水質(zhì)亞硝酸鹽氮的測(cè)定分光光度法》測(cè)定;NO3--N 濃度采用HJ/T 346-2007《水質(zhì)硝酸鹽氮的測(cè)定紫外分光光度法》測(cè)定;TN 濃度采用HJ 636-2012《水質(zhì)總氮的測(cè)定堿性過(guò)硫酸鉀消解紫外分光光度法》測(cè)定。
1.4 數(shù)據(jù)處理與統(tǒng)計(jì)分析
采用SPSS 17.0 對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行單因素方差分析,若存在顯著性差異,再進(jìn)行Duncan’s 多重比較以檢驗(yàn)組間的差異顯著性。以P<0.05 作為差異顯著水平,描述性統(tǒng)計(jì)值采用平均值±標(biāo)準(zhǔn)差(Mean±SD)表示。采用Excel 2016 對(duì)數(shù)據(jù)進(jìn)行折線圖繪制。
2 結(jié)果與分析
2.1 麥穗魚成活率
水蕹菜組和羅莎生菜組麥穗魚的存活率分別為86.11%和83.33%,顯著高于對(duì)照組(69.44%)(P<0.05)(表1)。
2.2 不同養(yǎng)殖系統(tǒng)pH 的變化情況
各系統(tǒng)pH 隨時(shí)間變化先下降再升高,三組pH均在6.66~7.77 之間波動(dòng),各組間無(wú)顯著差異(P>0.05)(圖1)。
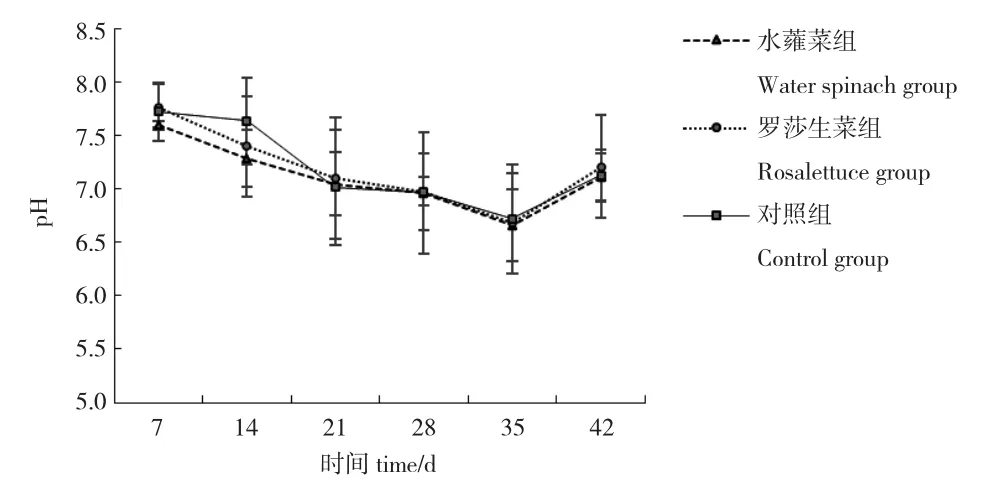
圖1 各系統(tǒng)pH 的變化情況Fig.1 Variation in pH values in different systems
2.3 不同養(yǎng)殖系統(tǒng)DO 的變化
如圖2 所示,在整個(gè)養(yǎng)殖周期內(nèi),對(duì)照組DO 濃度整體較水蕹菜組和羅莎生菜組高。其中,對(duì)照組DO 濃度顯著高于水蕹菜組(P>0.05);與羅莎生菜組相比,除第7 d 和第35 d 差異不顯著,其他結(jié)果均有顯著差異(P>0.05)(表1)。對(duì)照組DO 濃度分布在4.15~8.25 mg/L 之間。水蕹菜組和羅莎生菜組DO 濃度均在養(yǎng)殖前期(7~35 d)降低,養(yǎng)殖后期(35~42 d)升高,DO 濃度分別分布在2.30~5.90 mg/L和4.10~6.30 mg/L 之間。對(duì)比水蕹菜組和羅莎生菜組,除第28 d 和第42 d,羅莎生菜組DO 濃度均顯著高于水蕹菜組(P<0.05)。
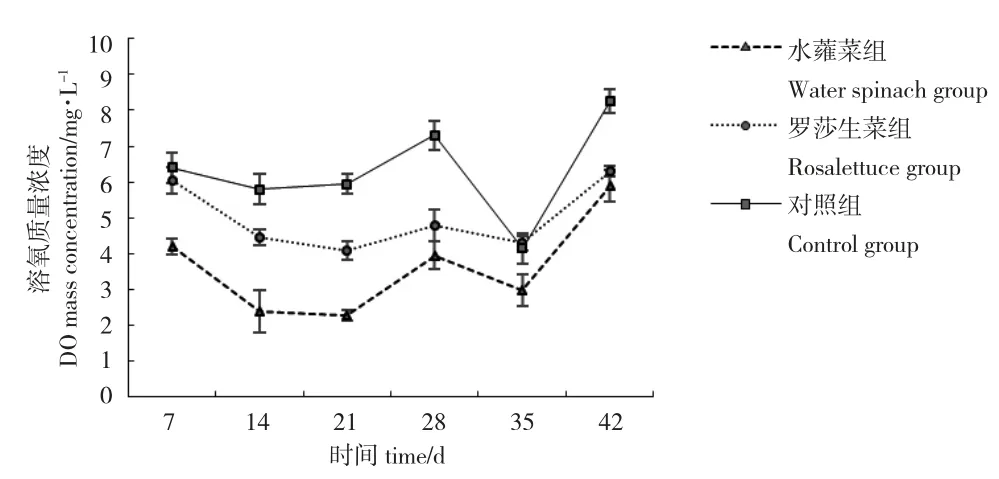
圖2 各系統(tǒng)溶氧的變化情況Fig.2 Variation in DO concentrations in different systems
2.4 不同養(yǎng)殖系統(tǒng)TAN 變化規(guī)律
水蕹菜組和羅莎生菜組TAN 濃度范圍分別為0.41~3.91 mg/L 和0.21~3.61 mg/L,這兩組TAN 濃度變化趨勢(shì)接近,在7~21 d 下降趨勢(shì)明顯,分別比養(yǎng)殖初期下降88.57%和94.26%,在21~42 d 維持在較低水平。對(duì)照組TAN 濃度分布在0.52~5.30 mg/L之間,在7~14 d TAN 濃度明顯上升,14~28 d 下降,但仍顯著高于水蕹菜組和羅莎生菜組(P<0.05),35~42 d 維持在較低濃度(圖3)。整個(gè)試驗(yàn)周期,水蕹菜組和羅莎生菜組前期顯著降低了系統(tǒng)中TAN濃度,試驗(yàn)后期TAN 維持在較低濃度,處理效果無(wú)顯著性差異(表1)。
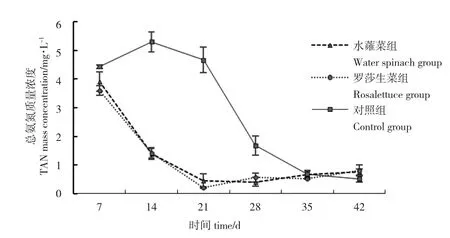
圖3 各系統(tǒng)總氨氮濃度的變化情況Fig.3 Variation in TAN levels in the different systems
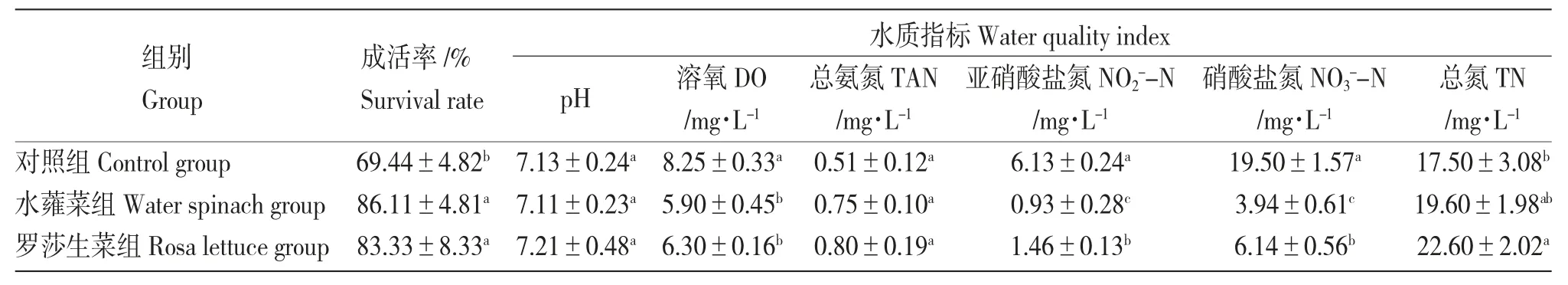
表1 各系統(tǒng)第42 d 麥穗魚成活率和水質(zhì)指標(biāo)Tab.1 The survival rate of topmouth gudgeon and water quality indicators in each system at day 42
2.5 不同養(yǎng)殖系統(tǒng)NO2--N 濃度變化規(guī)律
水蕹菜組、羅莎生菜組和對(duì)照組NO2--N 濃度范圍分別為0.61~4.40 mg/L、0.50~3.75 mg/L 和0.02~6.13 mg/L。在7~21 d 水蕹菜組和羅莎生菜組NO2--N 濃度明顯上升,21~35 d 出現(xiàn)波動(dòng),35~42 d明顯下降(圖4)。
在42 d 取樣時(shí),水蕹菜組和羅莎生菜組NO2--N 濃度維持在較低值,分別為0.93 mg/L 和1.46 mg/L。對(duì)照組從試驗(yàn)開始,NO2--N 濃度一直處于升高的趨勢(shì),在第42 d 達(dá)到最大值6.13 mg/L。試驗(yàn)結(jié)束時(shí)各組NO2--N 濃度差異顯著(P<0.05),濃度由大到小排列為:對(duì)照組>羅莎生菜組>水蕹菜組(表1)。
2.6 不同養(yǎng)殖系統(tǒng)NO3--N 濃度變化規(guī)律
水蕹菜組、羅莎生菜組和對(duì)照組NO3--N 濃度范圍分別為3.42~10.60 mg/L、2.13~8.25 mg/L 和0.98~19.50 mg/L。在7~21 d 水蕹菜組和羅莎生菜組NO3--N 濃度均先上升后下降,在21~35 d 又上升,35~42 d 均下降。對(duì)照組NO3--N 濃度基本保持上升趨勢(shì),其中,在28~42 d 上升幅度較大,最終達(dá)到一個(gè)高值(19.50 mg/L)(圖5)。試驗(yàn)結(jié)束時(shí)各組NO3--N 濃度差異顯著(P<0.05),濃度由大到小為:對(duì)照組>羅莎生菜組>水蕹菜組(表1)。
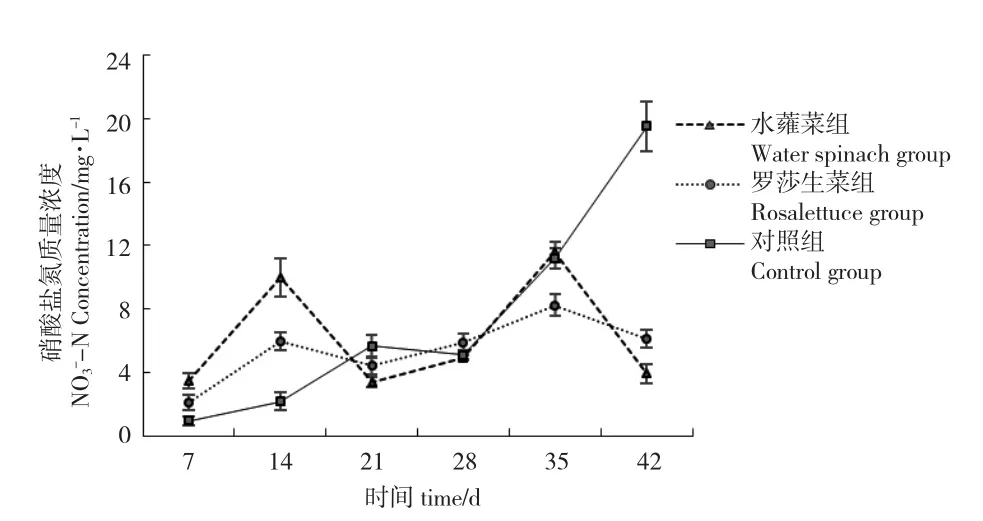
圖5 各系統(tǒng)硝酸鹽氮濃度的變化情況Fig.5 Variation in NO3--N levels in the different systems
2.7 水蕹菜組和羅莎生菜組TN 濃度變化
水蕹菜組和羅莎生菜組TN 濃度范圍分別為4.01~22.10 mg/L 和9.21~22.60 mg/L。水蕹菜組和羅莎生菜組TN 濃度變化趨勢(shì)接近,在7~21 d 先升高后降低,在21~35 d 又均出現(xiàn)上升趨勢(shì)。第35 d 之后,羅莎生菜組TN 濃度繼續(xù)上升,水蕹菜組TN 濃度出現(xiàn)下降趨勢(shì)(圖6)。試驗(yàn)結(jié)束時(shí)水蕹菜組TN濃度低于羅莎生菜組,但差異不顯著(P>0.05)(表1)。
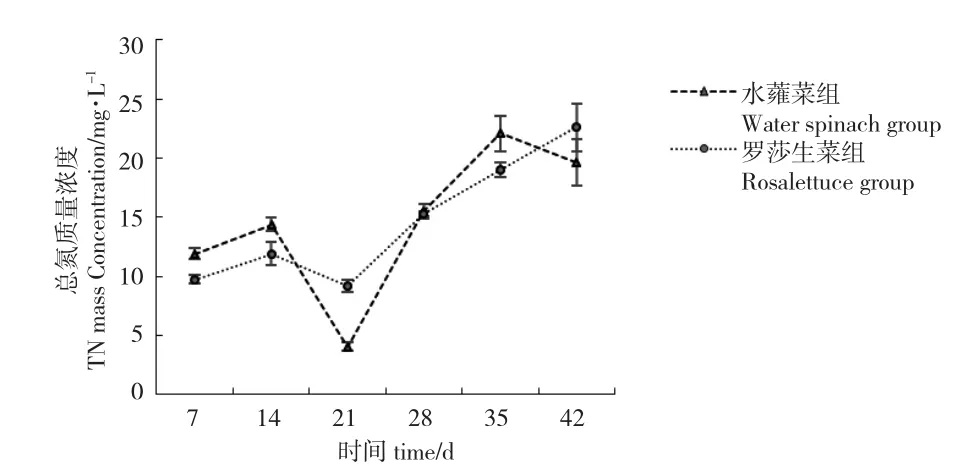
圖6 水蕹菜組和羅莎生菜組總氮濃度的變化情況Fig.6 Concentration changes of TN in two treatment groups
3 討論
3.1 水蕹菜和羅莎生菜對(duì)養(yǎng)殖水體pH 和DO 濃度的影響
麥穗魚適宜生存的pH 為5.50~9.50[11]。本試驗(yàn)各組pH 均在適宜區(qū)間內(nèi),能夠保證麥穗魚正常生長(zhǎng)。本試驗(yàn)中,對(duì)照組DO 濃度除第35 d 略小于羅莎生菜組外,其余均顯著高于水蕹菜組和羅莎生菜組(P<0.05),說(shuō)明生物浮床影響水體產(chǎn)生溶氧。生物浮床遮蔽部分水面,影響藻類的光合作用及水氣交換,導(dǎo)致水體溶氧輸入受限;發(fā)達(dá)的植物根系又為硝化細(xì)菌繁殖提供了良好的場(chǎng)所,促進(jìn)硝化反應(yīng)的進(jìn)行,該過(guò)程進(jìn)一步消耗了水體溶氧[12]。
3.2 水蕹菜和羅莎生菜對(duì)養(yǎng)殖水體TAN 和NO2--N 的影響
本試驗(yàn)第7~21 d,水蕹菜組和羅莎生菜組TAN濃度顯著下降,對(duì)照組TAN 濃度呈升高趨勢(shì),各組NO2--N 濃度均出現(xiàn)升高趨勢(shì),但對(duì)照組升高幅度小于水蕹菜組和羅莎生菜組(圖3、圖4)。在第14 d 和21 d,對(duì)照組TAN 濃度均顯著高于水蕹菜組和羅莎生菜組(P<0.05),而NO2--N 濃度均顯著低于水蕹菜組和羅莎生菜組(P<0.05)。試驗(yàn)前期的結(jié)果說(shuō)明:該階段麥穗魚處于調(diào)整適應(yīng)期,代謝較快,水體中TAN 濃度偏高。隨著試驗(yàn)進(jìn)行,水蕹菜組和羅莎生菜組植物根系中硝化細(xì)菌的數(shù)量和豐富度達(dá)到一定水平,加速TAN 向NO2--N 轉(zhuǎn)化[10]。
自第21 d 開始,水蕹菜組和羅莎生菜組TAN濃度基本穩(wěn)定,NO2--N 濃度出現(xiàn)波動(dòng)變化,最終維持在較低的濃度,說(shuō)明此時(shí)系統(tǒng)中硝化反應(yīng)正以穩(wěn)定速率進(jìn)行。對(duì)照組TAN 濃度自第21 d 開始下降,NO2--N 濃度持續(xù)升高。對(duì)照組TAN 濃度下降的原因,一是有大量TAN 向NO2--N 轉(zhuǎn)化,導(dǎo)致NO2--N濃度升高(圖4);二是NH3揮發(fā),對(duì)照組水體表面積大,使得NH3揮發(fā)量較大。該結(jié)果說(shuō)明:沒(méi)有蔬菜存在的系統(tǒng),有害氮鹽會(huì)持續(xù)累積,長(zhǎng)時(shí)期的養(yǎng)殖活動(dòng)容易對(duì)魚體生長(zhǎng)產(chǎn)生不利影響(表1)。
3.3 水蕹菜和羅莎生菜對(duì)養(yǎng)殖水體NO3--N 的影響
NO3--N 是TAN 經(jīng)硝化反應(yīng)的最終產(chǎn)物,毒性較低,只有達(dá)到較高濃度才會(huì)對(duì)魚體產(chǎn)生毒害作用[13]。試驗(yàn)初期(7~14 d)水蕹菜組和羅莎生菜組NO3--N 濃度較對(duì)照組明顯升高,說(shuō)明TAN 向NO3--N 轉(zhuǎn)化速率較高,而蔬菜根系對(duì)于NO3--N 的吸收成為加快該反應(yīng)進(jìn)行的促進(jìn)因素。試驗(yàn)初期,蔬菜根系吸收營(yíng)養(yǎng)鹽的能力還沒(méi)有達(dá)到最大化,容易造成水體中NO3--N 累積,因此在第14 d 水蕹菜組和羅莎生菜組NO3--N 濃度顯著高于對(duì)照組(P<0.05)。
自第14 d 開始,水蕹菜組和羅莎生菜組NO3--N 濃度沒(méi)有出現(xiàn)大幅度波動(dòng),說(shuō)明當(dāng)養(yǎng)殖系統(tǒng)進(jìn)入穩(wěn)定運(yùn)行階段后,蔬菜根系能夠以一定速度吸收水體中的NO3--N,且伴隨著水體DO 的消耗,根系表面噬氫菌屬、紅細(xì)菌屬等反硝化細(xì)菌對(duì)NO3--N的反硝化作用增強(qiáng),能夠?qū)⑵滢D(zhuǎn)化為N2排出養(yǎng)殖系統(tǒng)[20]。
自第7 d 開始對(duì)照組NO3--N 濃度一直呈上升趨勢(shì),與NO2--N 變化趨勢(shì)相似(圖5)。該結(jié)果說(shuō)明,沒(méi)有蔬菜的吸收作用,水體中氮鹽缺少有效的輸出途徑,長(zhǎng)時(shí)間累積會(huì)造成水體富營(yíng)養(yǎng)化,危及魚體生命。
3.4 水蕹菜和羅莎生菜對(duì)養(yǎng)殖水體凈化效果比較
試驗(yàn)第42 d,水蕹菜組NO2--N 和NO3--N 濃度均顯著低于羅莎生菜組(P<0.05),TN 濃度雖差異不顯著(P>0.05),但同樣低于羅莎生菜組(圖6),說(shuō)明水蕹菜吸收水體無(wú)機(jī)氮鹽,減少有害氮鹽的效果優(yōu)于羅莎生菜。韓璐瑤等[14]從吸收動(dòng)力學(xué)角度分析了水蕹菜和生菜根系對(duì)氮、磷營(yíng)養(yǎng)鹽的吸收能力,認(rèn)為水蕹菜擁有最大的吸收速率Imax和最小親和力常數(shù)Km,水蕹菜適宜任意濃度的水體凈化,而生菜更適合低氮、磷水平的水體凈化。戴習(xí)林等[15]在對(duì)蝦養(yǎng)殖水體的凈化研究中發(fā)現(xiàn),蕹菜平均日吸收氮量顯著大于生菜。
蔬菜對(duì)NO3--N 的吸收能力增強(qiáng),必然會(huì)加快水體中硝化反應(yīng)速率,大量消耗水體中DO。由DO數(shù)據(jù)可以看出,整個(gè)試驗(yàn)周期內(nèi)水蕹菜組DO 濃度整體偏低,除第42 d,水蕹菜組DO 濃度顯著低于羅莎生菜組(P<0.05)。由DO 數(shù)據(jù)可以進(jìn)一步驗(yàn)證,本試驗(yàn)條件下水蕹菜吸收氮鹽的效率高于羅莎生菜。
3.5 結(jié)論
本研究認(rèn)為,浮床覆蓋率為35%的麥穗魚養(yǎng)殖系統(tǒng)中,水蕹菜和羅莎生菜均能夠通過(guò)發(fā)達(dá)的根系形成穩(wěn)定的硝化環(huán)境,提高水體中的硝化效率,再加上植物根系對(duì)營(yíng)養(yǎng)鹽的同化作用,能夠有效去除水體中的有害氮鹽,保證水質(zhì)指標(biāo)穩(wěn)定,有利于麥穗魚的生長(zhǎng)。本試驗(yàn)條件下,水蕹菜的綜合凈水效果優(yōu)于羅莎生菜。
魚菜共生系統(tǒng)與單一養(yǎng)殖系統(tǒng)相比,增加了更多的人為控制環(huán)節(jié),系統(tǒng)更加復(fù)雜。因此,蔬菜在魚菜共生系統(tǒng)中對(duì)水質(zhì)的凈化研究尚有大量工作要做。
猜你喜歡
工業(yè)設(shè)計(jì)(2022年8期)2022-09-09 07:43:20
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測(cè)繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(shù)(2019年12期)2019-12-25 03:06:46
制造技術(shù)與機(jī)床(2019年10期)2019-10-26 02:47:06
中國(guó)洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
鐵道通信信號(hào)(2018年5期)2018-06-28 03:06:24
家庭影院技術(shù)(2017年9期)2017-09-26 03:41:45
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(shù)(2016年6期)2016-04-20 06:21:32
