野生與養殖魚類營養與品質差異研究進展
2023-02-26 06:02:22高欣石立冬任同軍韓雨哲
水產學雜志 2023年1期
關鍵詞:差異
高欣,石立冬,任同軍,韓雨哲,2
(1.大連海洋大學水產與生命學院,遼寧 大連 116023;2.大連海洋大學,遼寧省北方魚類應用生物學與增養殖重點實驗室,遼寧 大連 116023)
世界人口增長和人們對食用海產品有益健康的認識提高使全球魚類需求量一直在增加。魚類主要來源自自然資源和水產養殖[1]。然而,魚類總消費量的增加和不可持續的捕撈作業,單靠野生捕撈的魚類無法滿足日益增長的需求[2],水產養殖產業成為增加全球魚類供應的適宜手段[3],而養殖技術的多元化造成了不同養殖魚類的營養和品質區別。
魚肉主要化學成分(基礎營養成分、碳水化合物、維生素和礦物質等)的變化導致如風味、顏色、氣味、質地和表面外觀等屬性的變化,這些屬性決定了魚肌肉作為食物的可接受性[4]。與養殖群體相比,野生魚類的日糧組成受多種因素影響,包括生長動物的個體發育和所處地理位置。野生魚所捕食的食物可能有助于消化,形成特定的營養物質以及影響肉質的化合物的生物累積,但有毒重金屬的沉積量可能也較養殖群體高,例如汞、鎘和砷等[5,6]。養殖和野生群體除日糧組成、魚體健康和生長速度差異顯著外,其他因素也會影響魚肉營養和品質特性。例如,隨著動物年齡的增長、蛋白質合成活性降低、肌肉組織中游離氨基酸的含量增加,水體鹽度也會影響肌肉中游離氨基酸含量和口感[7]。目前國內對魚類營養和感官特性差異研究分析,主要集中在不同水體鹽度、飼料脂肪含量、養殖模式對其營養、質地和風味影響。然而針對不同品種的野生群體和養殖群體并沒有相關文獻綜述。本文比較了海、淡水中不同魚類養殖群體與野生群體的營養品質,為后續健康養殖技術升級及其飼料的研發提供基礎資料和理論依據。
1 野生和養殖魚類肌肉營養成分差異
1.1 魚類野生群體和養殖群體肌肉常規營養成分差異
魚肉作為一種蛋白質含量較高的食物,是人類重要的蛋白質來源。衡量魚類肌肉營養的指標主要包括蛋白質、脂肪、纖維、維生素及其他微量物質[4,5]。蛋白質是魚類肌肉中重要的營養成分,決定著肌肉的質量[6]。Haard(1992 年)根據溶解性不同,將肌肉中蛋白質分為三類:水溶性蛋白(主要是肌漿蛋白)、鹽溶性蛋白(主要是肌原纖維蛋白)和不溶性基質蛋白(主要是膠原蛋白)[7]。魚類肌肉中水溶性蛋白質含量較高,這可能與采食高能量的飼料和較少的運動有關,而較高的鹽溶性蛋白含量可導致肌肉失水的減少[8]。
表1 和表2 列舉了不同海、淡水魚類野生與養殖群體水分、粗脂肪和粗蛋白差異,發現野生群體粗脂肪含量均低于養殖群體,粗蛋白含量高于養殖群體。淡水野生群體水分含量高于養殖群體,而海水養殖和野生群體水分含量并無顯著差異。

表1 海水野生與養殖魚類基礎營養成分的差異(%)Tab.1 Differences in approximate nutrients between marine wild and cultured fishes(%)
有研究表明,養殖魚肉的水分含量較野生群體低,而脂肪較高[24,25]。Haard[7](1992)年提出水分含量影響魚肉品質,即水分含量越低,營養水平越高。Lopparelli 等[26](2004 年)也感官評定了不同脂肪水平的歐洲舌齒鱸(Dicentrarchus labrax)[27]的肌肉,發現高脂組的多汁性和嫩度顯著高于低脂組,人工養殖魚類的基本營養水平可能更高,肉質更多汁鮮嫩。這可能是人工飼料更接近魚類的營養需求和飲食習慣,而一些天然水體生長的魚類的日糧組成主要為軟體動物(腹足類和貝類),也可能會遭到饑餓,能量攝取不如養殖魚類。溫度、光照和生長階段等因素也影響肌肉組成和脂肪沉積。例如,持續光照會導致脂肪含量減少[28];夏末秋初脂肪儲備增加,而冬季過后脂肪儲備減少[29];生殖繁殖期的野生洄游魚類脂肪含量會較高[30]。因此要考慮野生和養殖魚類肌肉營養成分與含量的差異,還要考慮個體因素、餌料組成成分、生長階段、水溫和生存環境等因素。
1.2 野生和養殖魚類肌肉脂肪酸含量差異
n-3 多不飽和脂肪酸(PUFA)對人體有重要的生理功能,尤其是二十碳五烯酸(EPA)和二十二碳六烯酸(DHA),可以調節炎癥和免疫過程以及預防心血管疾病等[31],是衡量魚肉質量的重要指標。EPA和DHA 對魚類生長發育、繁殖性能和免疫視覺靈敏度等有重要作用[32,33],幼魚生長中對DHA 的需求量較高[34-36]。但過量的EPA 和DHA 會抑制水產動物的生長。如星斑川鰈(Starry Flounder)[37]和半滑舌鰨(Cynoglossus semilaevis)[32]幼魚隨著飼料DHA/EPA 值的升高,增重率、飼料轉化率等呈先升高后降低的趨勢。因此只有適宜比例的EPA 和DHA 才可以提高魚類幼體的生長率和存活率。
魚類需要的PUFA 有n-6 和n-3 兩種類型,主要包括18:2(n-6)、18:3(n-3)、EPA 和DHA。大多數魚類無法直接合成或僅能由特定前體物形成PUFA,其中18:2(n-6)和18:3(n-3)所有脊椎動物都不能直接合成,而養殖魚類可以通過飼料獲得[38]。n-3 PUFA 還具有抗炎癥、降血脂、抗氧化和改善胰島素抵抗的作用,而n-6 PUFA 具有促炎的作用。n-3/n-6 PUFA 與II 型糖尿病、冠心病、肥胖癥等疾病有關[39]。譚青等[40]研究了不同比率n-3/n-6 PUFA對大菱鲆(Scophthalmus maximus)幼魚生長性能及飼料利用、體組成和消化酶活性的影響,發現n-3/n-6 PUFA 水平較低時,對大菱鲆幼魚有降脂作用,同時機體對蛋白質的消化吸收能力下降。
表3 列舉了不同野生與養殖海水魚類脂肪酸含量的差異,發現養殖群體的n-3 PUFA 普遍高于野生群體,但野生大黃魚(Pseudosciaena crocea)[41]較養殖群體高68.75%;養殖群體中的EPA/DHA、EPA+DHA 和n-3/n-6 PUFA 均較野生群體高。表4列舉了不同淡水野生與養殖魚類脂肪酸含量差異,發現野生群體n-3 PUFA 和n-3/n-6 PUFA 普遍較養殖群體高,而養殖群體EPA/DHA 和EPA+DHA高于野生群體。

表4 淡水種類的野生群體與人工養殖魚類脂肪酸含量差異(%)Table 4 Differences in fatty acid contents between freshwater wild and cultured fish(%)
造成脂肪酸差異的原因可能為:淡水魚類和鮭鱒魚類具有將PUFA 合成高度不飽和脂肪酸的能力,而海水魚類的仔稚魚自身無法直接合成n-3 PUFA 或合成能力很弱,只能從無節幼體和輪蟲中獲取[45]。李松等[46]對比了不同養殖模式和餌料組成的大黃魚脂肪酸含量發現,多通框網箱養殖魚中PUFA 含量優于筏式小網箱養殖魚,配合飼料組魚的單不飽和脂肪酸(MUFA)優于冰鮮魚餌料組;可能在季節性和性別間表現出相當程度的差異,表明年齡組成與血脂和體脂的正相關性[47]。
1.3 野生和養殖魚類肌肉氨基酸含量差異
蛋白質的品質主要取決于氨基酸的種類組成和含量,特別是必需氨基酸(EAA)的供應[48]。必需氨基酸缺乏會造成魚體生長緩慢、飼料利用率降低等。用必需氨基酸(∑EAA)、總氨基酸(∑TAA)和必需氨基酸指數(EAAI)等指標評價野生和養殖魚類氨基酸含量的差異。
表5 和表6 分別列舉了不同海、淡水野生與養殖魚類∑EAA、∑TAA、EAAI 含量差異,發現野生海、淡水魚類∑EAA、∑TAA 和EAAI 含量普遍高于養殖魚類。
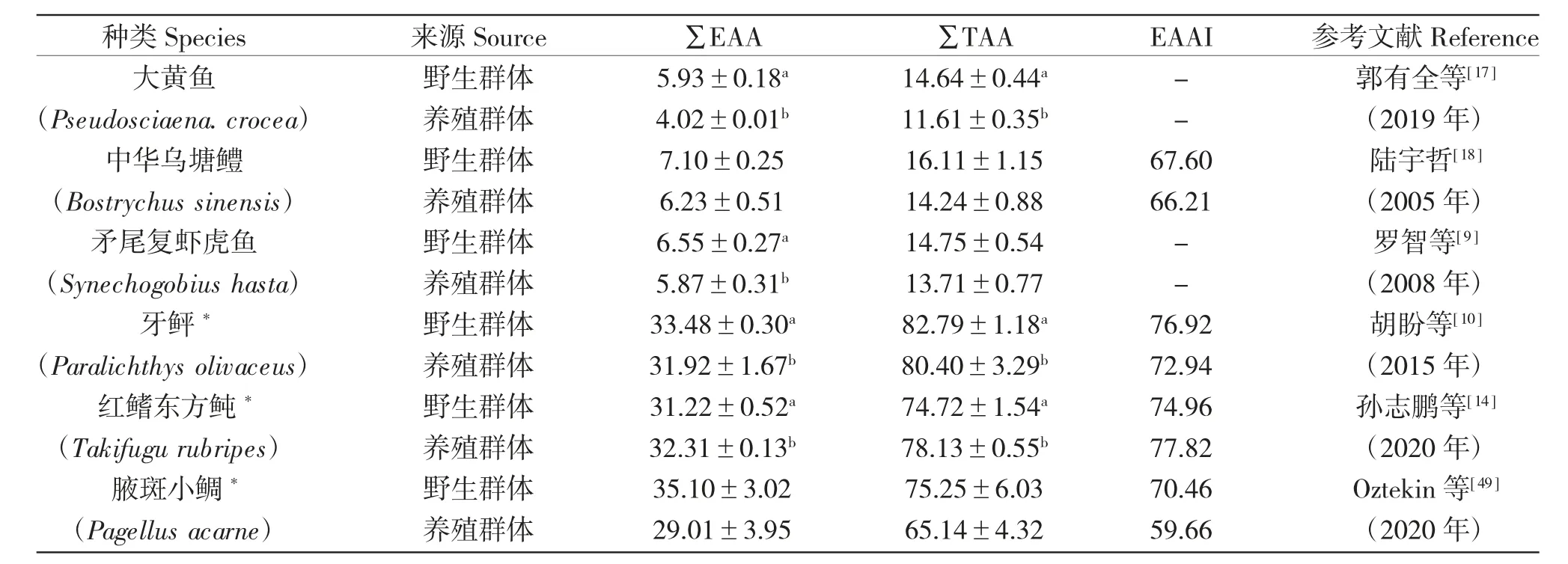
表5 海水野生與養殖魚類∑EAA、∑TAA、EAAI 含量的差異(%)Tab.5 Differences in contents of ∑EAA,and ∑TAA and EAAI between marine wild and cultured fish(%)

表6 淡水種類的野生群體與人工養殖魚類∑EAA、∑TAA、EAAI 差異(%)Tab.6 Differences in contents of ∑EAA,and ∑TAA and EAAI between freshwater wild and cultured fish(%)
造成野生與養殖魚類氨基酸差異的原因可能是:一是食物組成差異,養殖魚類所需氨基酸來源主要是飼料中的植物蛋白源,而野生群體氨基酸主要來源天然餌料。不同蛋白源的氨基酸消化率差異很大,也可能影響養殖魚的體組成和營養品質;二是受生長環境和生長階段影響,野生魚類可能長期處在饑餓狀態,游離組氨酸含量食物充足的養殖魚少和鵝肌肽含量更多;三是性成熟期間,肌肉中游離氨基酸含量減少,性腺中氨基酸含量增加[51]。
1.4 野生和養殖魚類肌肉礦物質元素差異
礦物質是所有生物生長、骨骼礦化、繁殖和能量代謝所必需[52]。而礦物元素作為魚肉中的重要組成成分,主要集中在魚脊椎、鱗和肌肉中,是人類礦物質重要的來源[53]。魚類生活史和攝食是影響其組織礦物質組成的重要因素。鈣、磷、鉀、錳、鈉、氯化物和硫等15 種微量元素,大部分已在魚類組織中檢測到[54,55],而礦物質來源不同,也會影響礦物質的生物利用度[56]。在自然條件下,魚類從生長環境和由多種生物組成的天然食物中吸收和保留大部分元素,而在養殖條件下,魚類的礦物質營養幾乎完全依賴所投喂的飼料。淡水野生中華鱘(A.sinensis)和大麻哈魚(Oncorhynchus.Keta)中鈣、鎂和鋅等含量較養殖群體高,野生大麻哈魚[57]鋅含量是養殖群體的16 倍,在中華鱘(A.sinensis 養殖群體中未檢測到鐵和硒。養殖的烏鱧(Channa argus[58]、黃顙魚(P.fulvidraco)、齊爾白鮭(Coregous nasus)[59]和翹嘴紅鲌(Ergthroculter ilishaeformis)[60]中鈣、磷和錳等含量較野生群體高,尤其是齊爾白鮭的鈣含量是野生群體的6 倍,烏鱧中錳含量是野生群體的39 倍。
海水野生群體礦物質含量普遍較養殖群體高,例如野生黑雕(Sparus macrocephalus)和黃姑魚(Nibea albiflora)[61]含硒量分別較養殖群體高43.47%和51.72%,尤其野生半滑舌鰨含硒量是養殖群體的6 倍。但也有研究得出相反結論。如養殖牙鲆(P.olivaceus)較野生群體礦物含量高,其中硒含量較野生群體高16.12%,錳含量比野生群體高1 倍。
通過不同種類的淡、海水野生和養殖魚類的礦物質含量對比表明,淡水野生魚類鎂和鋅含量較養殖群體高,養殖魚類磷和錳含量較野生群體高;海水野生魚類礦物質普遍較養殖群體高,只有牙鲆相反,原因可能是為了滿足市場需求,其生長特性在養殖產業不斷發展、優化,所以養殖魚類品質優于野生群體;海、淡水野生魚類含硒量都較豐富。硒作為谷胱甘肽過氧化物酶的組成部分,具有免受自由基氧化傷害、抗衰老和增強創傷組織再生能力等,對人體健康極為重要[62]。水質變化也影響魚類肌肉組織中礦物質的濃度,例如,海水養殖的二倍體虹鱒(Oncorhynchus mykiss)肌肉組織比淡水養殖的虹鱒具有更好的礦物質來源[63]。
2 野生和養殖魚類肌肉質構特性差異
一般來說,人們更傾向于肉質彈性好、咀嚼性高的魚肉。表7 和表8 分別列出了海、淡水野生魚類與養殖魚類質構差異,表明不論海水還是淡水種類的野生群體魚類肌肉的內聚性、彈性、咀嚼性和硬度均較養殖群體高。
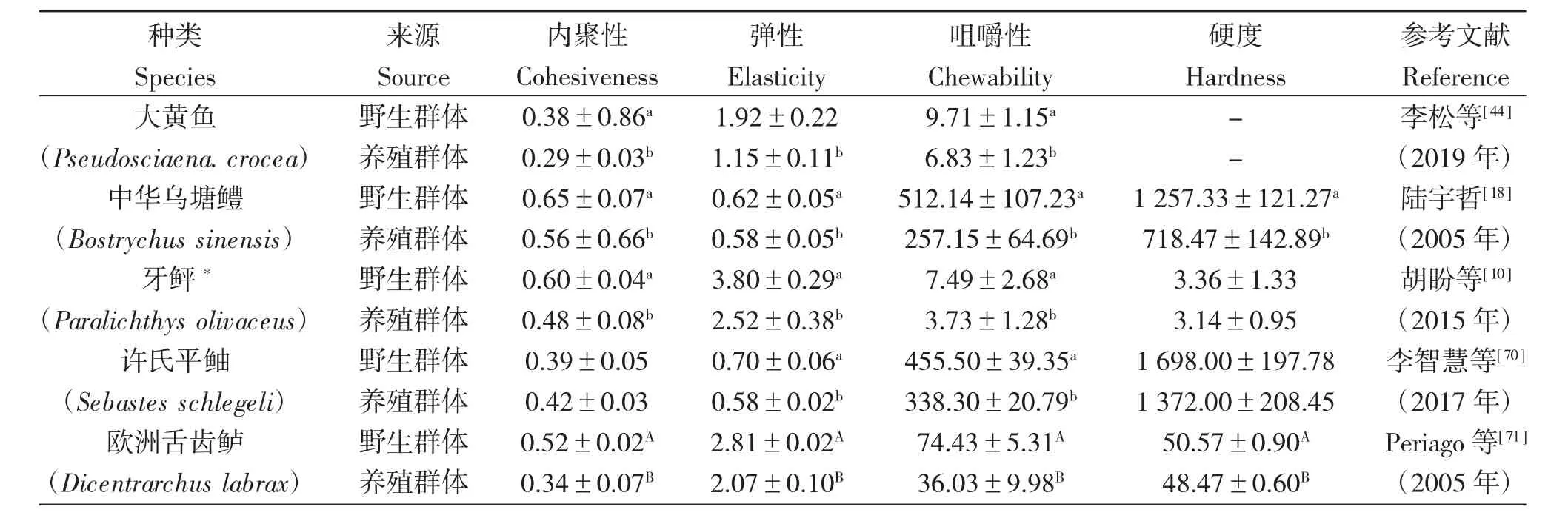
表7 海水種類的野生群體與人工養殖魚類質構測定差異Tab.7 Differences in texture between marine wild and cultured fishes

表8 淡水種類的野生群體與人工養殖魚類質構測定差異Tab.8 Differences in texture between freshwater wild and cultured fishes
質構特性的比較表明:大多數海、淡水野生魚類的彈性、咀嚼性高于養殖群體。消費者往往更喜歡彈性和咀嚼性高的肉質[64]。肌肉硬度差異的原因主要是野生和養殖魚的攝食、運動等差異所致。肌纖維的數量、大小和分布也影響魚肉質地。較小肌纖維的魚肉硬度更高[65],其中肌肉中脂肪含量和膠原蛋白的穩定性也可能為重要的影響因素,魚肉的肌理與肌肉脂肪含量呈負相關[66];膠原蛋白是細胞外基質的主要組成部分,其數量和交聯與魚肉的紋理呈正相關[67]。養殖者為追求利益最大化,會投喂促生長的配合飼料,然而肉質的硬度與增長速度有關,快速生長會降低魚的肉硬度[68],這往往會使魚肉的口感大幅度下降。魚肉質地也受pH 和溫度的影響,會使蛋白質變性或水解,導致魚肉組織軟化[ 69 ]。
3 野生和養殖魚類形態差異
魚體的形態變化與魚類的攝食、放養密度、棲息地和捕食風險等因素相關[76]。研究表明,眼徑大小的變化可能與水透明度和光照條件有關[77];頭部或口形態的差異可能與喂養生態環境或食物大小和位置組成有關[78];魚的體型通常與水流速有關。體型纖細延長的魚通常胸鰭相對較長[79]。與養殖個體相比,野生黑鯛(S.macrocophalus)體延長、體型更偏紡錘型、腹部較小且背鰭更銳利[6];養殖大黃魚眼徑大、體高,野生大黃魚的吻長和眼后頭長則大于養殖大黃魚[80]。野生群體在天然餌料中會沉積特定的色素,而在養殖魚類難以從餌料獲得足夠的相關色素沉積,天然體色消褪[81]。野生真鯛(Pagrosomus major)較養殖群體體窄、體型緊湊、體色更鮮艷、鱗片有光澤[82]。綜上所述,養殖群體普遍較野生群體肥滿度高、體寬、腹部偏圓、體色暗淡。
4 展望與未來
衡量魚體肉質的指標及其影響因子很多,不能僅通過感官評價和營養價值而論,還需考慮到品種、地域、水源以及對應的水產品安全性的差異。野生魚需要考慮苗種來源、環境狀況;養殖魚需要了解其養殖方式、飼養環境、藥物殘留以及飼料管控,因此野生群體并不一定比養殖群體安全性更高。大多數規模化養殖魚類的色澤、口味和質地等不如野生群體,這也是消費者主觀認為野生魚類品質更高的主要原因之一。因此,管控好養殖環境、選育優良種質和減少病害的發生,是改善養殖魚類感官品質的重要途徑。如何通過營養調控手段改善魚體營養組成、魚體肉質風味及魚肉組織學結構,也是魚類營養學的一個重要而復雜的研究課題。
有關野生與養殖魚類的營養與品質等需進一步研究:(1)野生與養殖魚代謝調控和轉錄組的檢測,如不同養殖密度的魚類可能經歷各種應激,導致魚體代謝、免疫系統和魚肉品質產生不利影響;(2)野生與養殖魚類食用安全性及健康風險評估等,如野生與養殖魚類組織中重金屬分布特征及安全性評價,尤其是肉食性魚類重金屬累積較高;(3)野生與養殖紅鰭東方鲀的河豚毒素組織分布、含量及營養價值比較研究。對野生與養殖群體的生理生化水平研究,以期了解不同養殖模式的生存狀態,可通過營造仿生態環境的技術,改善養殖群體遺傳多樣性下降的現狀。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
