堿脅迫下內生真菌對宿主布頓大麥顯微結構的影響
2023-03-05 13:38:12戶萌菲劉小珍陳水紅
草地學報 2023年2期
龍 鳳, 戶萌菲, 陳 勝, 劉小珍, 陳水紅
(塔里木大學生命科學與技術學院/塔里木盆地生物資源保護利用兵團重點實驗室, 新疆 阿拉爾 843300)
目前,大多數關于作物脅迫的研究主要集中在鹽脅迫上,鹽堿混合脅迫研究相對較少。然而,在自然界中,高濃度的鹽和高pH值經常同時發生,它們的協同作用可能比單獨使用任何一種脅迫的影響對植物生長和發育更有害[1]。植物受到堿脅迫時,通常會改變自身根莖葉形態結構和生理過程去適應環境。植物主要通過表皮的增厚、葉片肉質加厚、葉片縮小和改變濃密短柔毛的覆蓋等變化來增強對環境的適應能力[2]。有研究表明,能夠在高濃度堿脅迫下生長的耐鹽植物,是由長期的進化過程中通過改變特定的形態組織結構來適應逆境,降低鹽離子帶來的傷害[3]。了解禾草植物如何改變形態結構來應對堿脅迫帶來的傷害,有助于為后期在飼用牧草和鹽堿地的改良方面提高理論基礎。
禾草內生真菌是指在宿主禾草體內度過部分或者全部生命周期,但不引起宿主植物任何癥狀的一類真菌[4]。目前,關于禾草內生真菌的研究主要集中在子囊菌門(Ascomycota)麥角菌科(Clavicipitaceae)的香柱菌(Epichloё)屬[5]。在鹽堿脅迫下,禾草內生真菌的侵染能夠提高宿主植物的對外界環境的耐受性和適應性[6-8]。布頓大麥(Hordeumbogdanii)是一種耐鹽植物,它具有較強的生態適應性以及競爭優勢,在作為飼用牧草和荒漠及鹽堿地改良草種等方面具有廣闊的應用價值[9]。目前關于布頓大麥-內生真菌共生體的研究主要集中在生長發育[10-13]、生理生化[12,14]、營養價值[15]等方面,但該共生體在非生物脅迫下細胞顯微結構的響應研究較少。因此,為了探究內生真菌侵染對宿主布頓大麥在堿脅迫條件下的顯微結構是否有影響,本研究運用石蠟切片技術觀察內生真菌侵染對布頓大麥根、莖、葉在不同堿處理下各個顯微結構的變化,為內生真菌提高宿主布頓大麥耐堿性的生理機制提供形態學基礎。
1 材料與方法
1.1 試驗材料
實驗室前期處理得到的溫宿縣E+與E-布頓大麥種子[15],并鑒定種子中帶有內生真菌屬于Epichloё屬[16]后,將種植在塔里木大學動科試驗站的試驗田(80°76′E,41°58′N,海拔1 514 m)繁種子,收獲種子作為試驗的材料。分別挑取E+,E-飽滿的種子進行清水萌發,長出子葉后,移栽入4 L水培盒中,每盆19孔,每孔1個苗。每盆連接一個充氧泵,每天定時進行換氣,每3 d更換一次1/2 Hoagland營養液。溫室的晝/夜溫度為(28±2)℃/(15±3)℃,日照時長約為12 h。待布頓大麥分蘗3~5株時,用李春杰等[17]的莖髓苯胺藍鏡檢法檢測布頓大麥是否含有內生真菌,不帶菌的為E-,帶菌為E+。
1.2 堿脅迫處理
混合堿(Na2CO3∶NaHCO3=1∶1)處理,濃度梯度為50 mmol·L-1,100 mmol·L-1,每個處理重復4次,為了減少堿沖擊效應,每12 h遞增50 mmol·L-1堿(將前一次的堿溶液換成增加后的溶液),直到所有處理達到預定濃度,未加混合堿的處理作為試驗的對照,每7 d更換一次對應濃度的混合堿溶液,至21 d取樣,進行后期指標測定。
1.3 顯微切片制作與觀察
將各堿處理E+和E-布頓大麥,分別根(分生區)、莖(根頸)、葉(中央)剪成約1 cm小段,放裝有70% FAA(70%酒精∶冰醋酸∶甲醛=90∶5∶5)青霉素小瓶中,然后用注射器抽取瓶中空氣使樣品沉入瓶底固定24 h。參照黃倩[18]的石蠟切片制作方法,固定完成后將材料取出進行脫水、透明、浸蠟、包埋、切片(厚度為10 μm)、粘片、脫蠟、染色(番紅-固綠雙重染色)、封片(加拿大樹膠,solarbio公司CAS:8007-47-4),最后用研究級正置生物顯微鏡(日本,尼康Nikon Eclipse Ci-L)觀察,利用NIS-Elements軟件進行拍照并測量根的維管束面積、表皮厚度、內皮層厚度和導管直徑,測量莖的維管束面積、氣腔直徑和表皮厚度,測量葉片的維管束面積、泡狀細胞面積、氣孔直徑和導管直徑。每個樣品測量3個樣本,每個樣本測量8個視野,以平均值表示。
1.4 統計分析
采用SPSS統計軟件(IBM SPSS Statistics 26)進行數據分析,圖中數據為平均值±標準誤差。采用雙因素方差分析方法檢測堿處理和內生真菌對布頓大麥根莖葉顯微結構的影響,采用單因素ANOVA方差分析法檢測不同堿濃度處理對E+和E-布頓大麥根莖葉顯微結構的影響。采用獨立樣本T檢驗來檢測同一堿濃度下E+和E-根莖葉顯微結構的影響。統計顯著性差異定義在95%置信水平。
2 結果與分析
2.1 堿脅迫對布頓大麥植株根的顯微結構影響
由圖1可知,布頓大麥根部結構主要由維管束、內皮層和表皮組成。堿脅迫對布頓大麥根的細胞結構影響較為明顯。隨著堿濃度的升高,根部細胞外皮層從50 mmol·L-1開始出現皺縮和破裂,到100 mmol·L-1處根部細胞外表皮層皺縮加深。另外,E+植株根中的導管數明顯高于E-植株。由表1可知在0 mmol·L-1時維管束面積E+顯著高于E-(P<0.05);堿處理后布頓大麥的維管束面積在縮小,在50 mmol·L-1處E+與E-的維管束面積差異不顯著,但在100 mmol·L-1處E+的維管束面積顯著高于E-(P<0.05)。堿脅迫導致對根的導管直徑增加,E+的導管直徑長于E-。堿脅迫對根的表皮厚度的影響,在無堿處理下表皮厚度E+與E-無顯著差異,但在堿脅迫下E+的表皮厚度顯著高于E-;同時,布頓大麥內皮層隨著堿濃度的升高植株根內皮層在不斷加厚,在50 mmol·L-1處E+顯著高于E-的(P<0.05)。雙因素方差分析后,發現內生真菌和堿處理顯著影響維管束面積、導管直徑、表皮厚度和內皮層厚度;除了導管直徑以外,上述指標的內生真菌與堿處理兩者之間存在交互作用(P<0.05)。
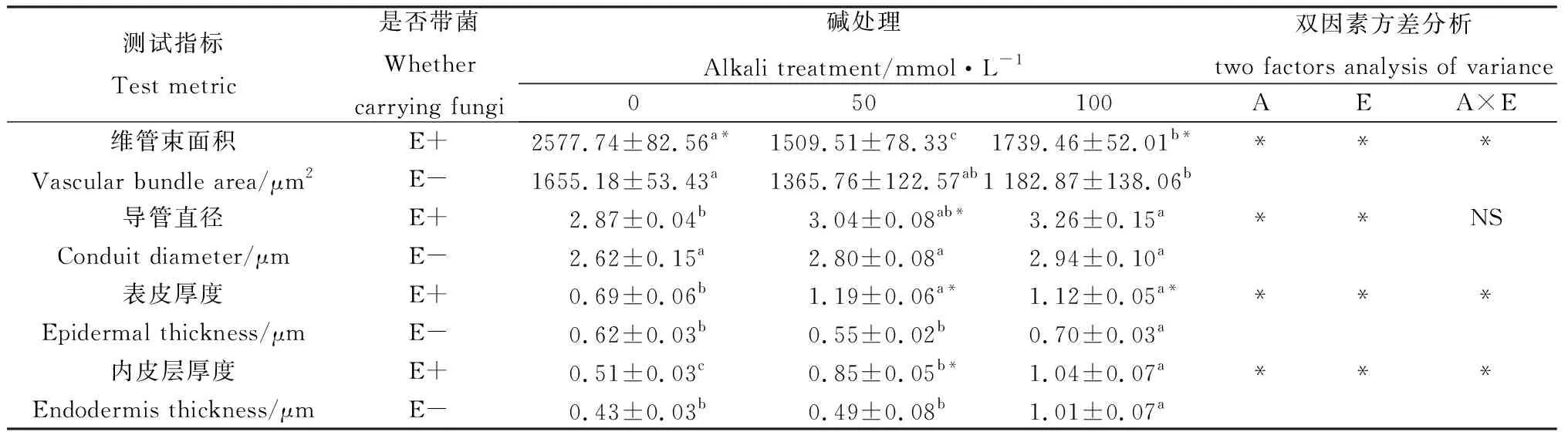
表1 堿脅迫下內生真菌對布頓大麥根結構的影響Table 1 Effects of endophytic fungi on root structure of Hordeum bogdanii under alkali stress
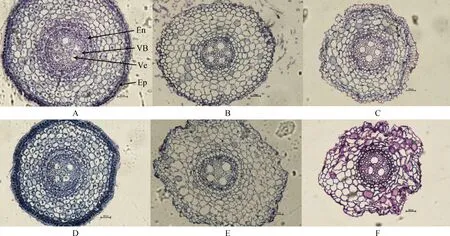
圖1 不同堿濃度對E+和E-布頓大麥根的橫切顯微結構的影響Fig.1 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii roots注:A,B,C分別為E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分別為E- 0 mmol·L-1,E-50 mmol·L-1,E- 100 mmol·L-1;En,內皮層;VB,維管束;Ve,導管;EP,表皮;標尺為200 μm,放大倍數為20×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+100 mmol·L-1 respectively;D. E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;En,endothelial layer;VB,vascular bundle;Ve,catheter;Ep,epidermis;Scale is 200 μm,the magnification is 20×10
2.2 堿脅迫對布頓大麥植株莖的顯微結構影響
不同濃度堿脅迫對E+和E-布頓大麥莖的顯微結構的影響見圖2和表2。由表2可知,堿脅迫對布頓大麥E+和E-維管束面積無顯著影響,并且E+與E-在維管束面積上無顯著差異。堿脅迫縮小了E+植株的氣腔直徑,對E-無顯著影響;在0 mmol·L-1和100 mmol·L-1堿脅迫發現E+的氣腔直徑顯著長于E-(P<0.05)。隨著堿濃度的增加莖的表皮厚度在逐漸增加。由表2雙因素方差分析可知,堿脅迫和內生真菌顯著影響表皮厚度,但兩者之間不存在交互作用。
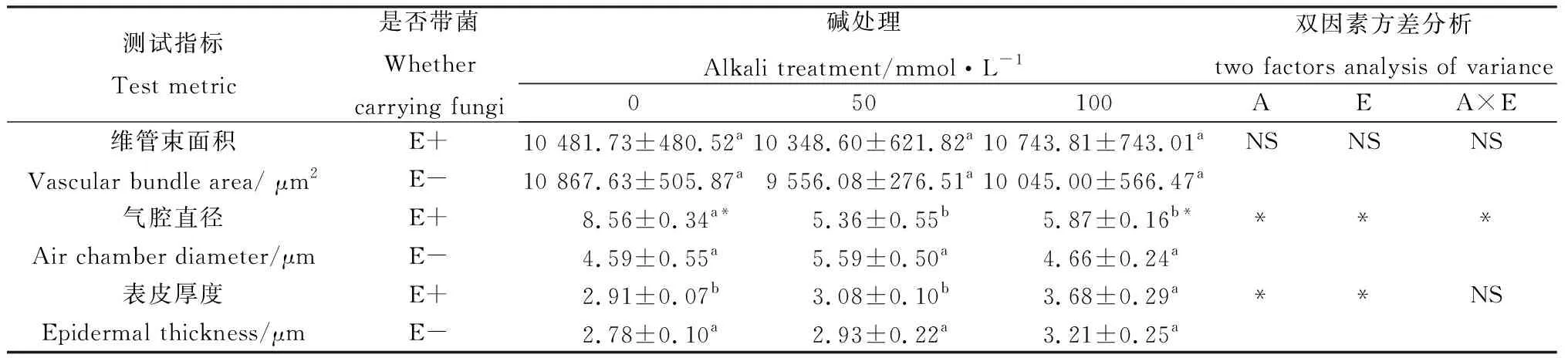
表2 堿脅迫下內生真菌對布頓大麥莖結構的影響Table 2 Effects of stem structure of Hordeum bogdanii by endophytic fungi under alkali stress
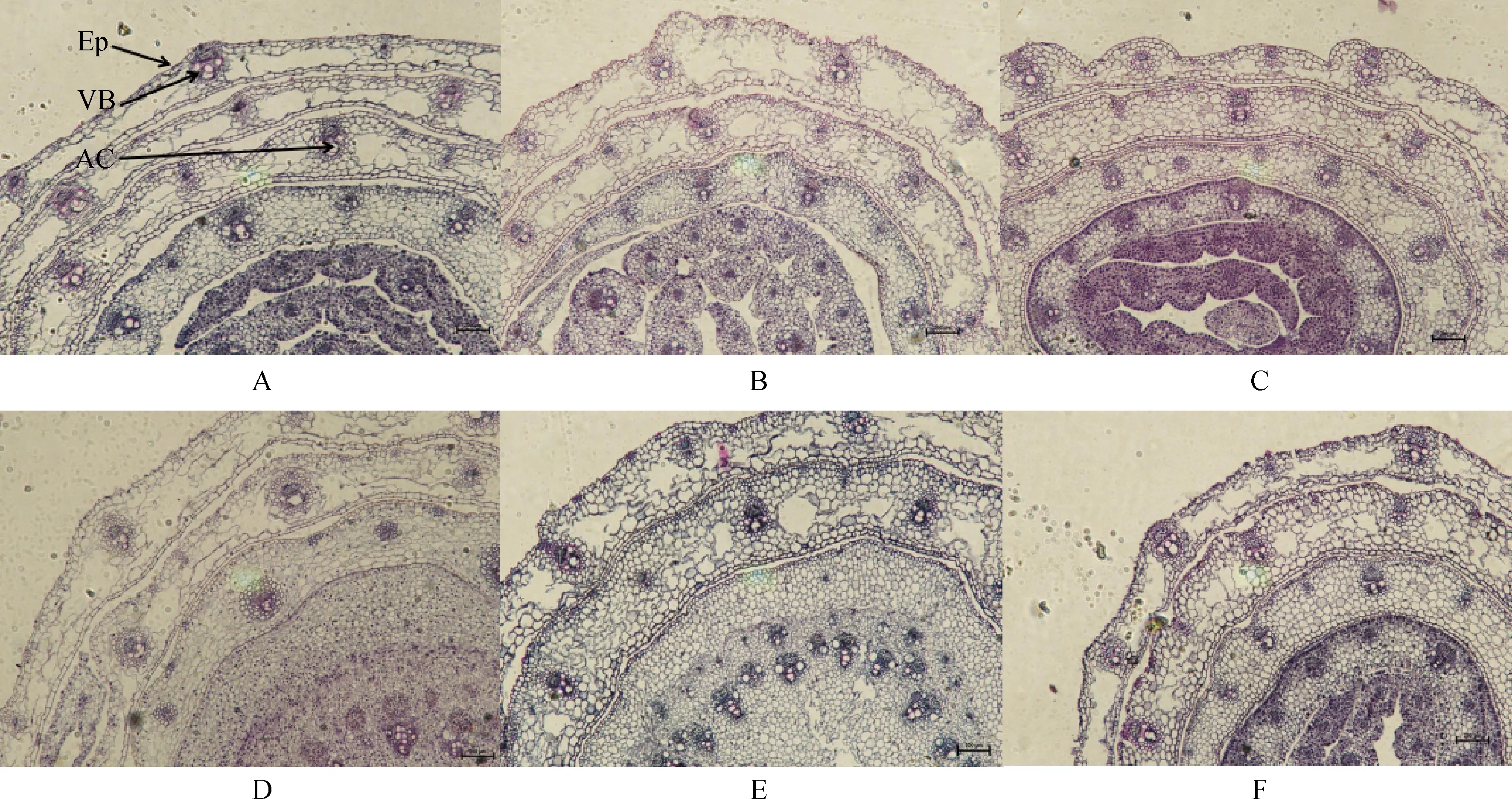
圖2 不同堿濃度對E+和E-布頓大麥莖的橫切顯微結構的影響Fig.2 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii stems注:A,B,C分別為E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分別為E- 0 mmol·L-1,E- 50 mmol·L-1,E- 100 mmol·L-1;En,內皮層;VB,維管束;AC,氣腔;EP,表皮;標尺為200 μm,放大倍數為10×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+ 100 mmol·L-1 respectively;D,E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;En,endothelial layer;VB,vascular bundle;AC,air chamber;Ep,epidermis;Scale is 200 μm,the magnification is 10 × 10
2.3 堿脅迫對布頓大麥植株葉的顯微結構影響
從圖3葉片的顯微結構圖可知,布頓大麥的上下表皮細胞大小不規則,上下表皮都存在氣孔,泡狀細胞只存在于上表皮。隨著堿濃度的升高,布頓大麥葉的維管束面積逐漸降低,泡狀細胞的細胞面積增大,同一堿濃度處理下E+的維管束面積和泡狀細胞面積高于E-,并且維管束面積達到顯著水平(P<0.05)。堿脅迫縮小了E-的導管直徑,但對E+的導管直徑無顯著影響,在100 mmol·L-1堿濃度下E+的導管直徑顯著大于E-(P<0.05)。在葉片氣孔直徑中發現50 mmol·L-1堿濃度處理下E+的顯著大于E-(P<0.05)。通過雙因素方差分析發現堿脅迫顯著影響布頓大麥葉的維管束面積、泡狀細胞面積、導管直徑以及氣孔直徑;內生真菌顯著影響葉片的維管束面積和氣孔直徑(P<0.05),但內生真菌和堿脅迫不存在交互作用(表3)。
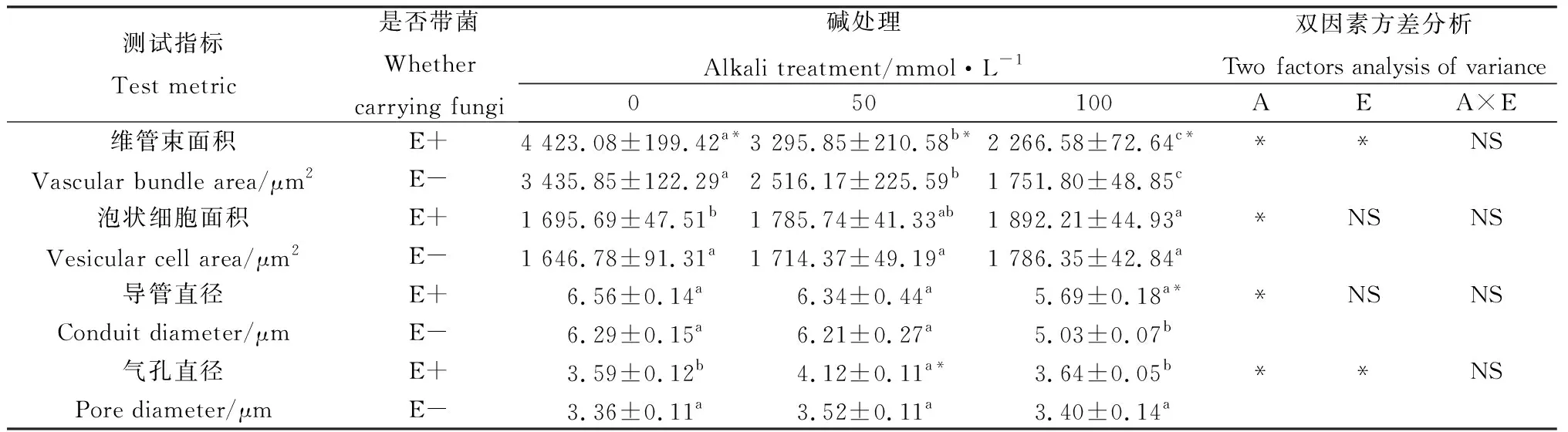
表3 堿脅迫下內生真菌對布頓大麥葉結構的影響Table 3 Effects of leaf structure of Hordeum bogdanii by endophytic fungi under alkali stress
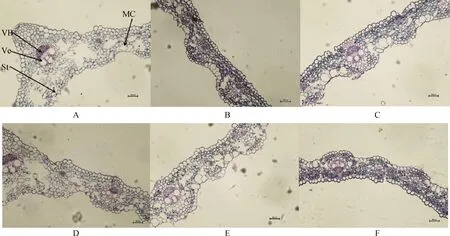
圖3 不同堿濃度對E+和E-布頓大麥葉的橫切顯微結構的影響Fig.3 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii leaves注:A,B,C分別為E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分別為E- 0 mmol·L-1,E-50 mmol·L-1,E- 100 mmol·L-1;VB,維管束,Ve,導管;St,氣孔;MC,泡狀細胞;標尺為200 μm,放大倍數為20×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+ 100 mmol·L-1 respectively;D,E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;VB,vascular bundle;Ve,catheter;St,air hole;MC,vesicular cells;Scale is 200 μm,the magnification is 20×10
3 討論
植物的形態結構總是與環境相適應。逆境環境影響植物生長,并可引起植物形態結構的相應變化[19]。維管束是主要輸水組織,堿脅迫導致根的維管束橫截面面積減小,韌皮部和木質部的傳導能力明顯減弱[20]。趙海新等[21]的研究也證實了堿脅迫可促使植物根系維管束數目減少、木質化程度加強、導管數量增加等方式來提高根的運輸能力。此外,陳泰祥[22]研究發現堿脅迫對野大麥的生長影響強于鹽脅迫;內生真菌侵染減緩了宿主野大麥(Hordeumbrevisubulatum)在鹽脅迫下根和莖的皮層厚度的降低趨勢,增強了根的內皮層厚度的增高趨勢;同時內生真菌也減緩了葉片厚度、維管束的降低趨勢[23]。本研究與陳泰祥的研究結果相似,堿脅迫導致布頓大麥根維管束橫截面積減小,木質化程度加深,導管直徑變大,這些結構的變化可能是布頓大麥為了適應堿脅迫帶來的損壞做出的響應。同時,植物在受到外界環境的刺激下,植物會增加表皮厚度和內皮層厚度來防止體內水分過分的蒸騰,保持水分,并起著機械支持作用[24]。內生真菌的存在對植物顯微結構的改變有關,真菌侵染與葉片木質部、葉片韌皮部、莖維管束、莖木質部導管和根間木質部面積顯著相關。在NaCl脅迫下,Epichlo?內生真菌侵染對野大麥的葉脈、莖表皮、根皮層和根內皮層厚度增加有關[25]。本研究發現,Epichlo?內生真菌提高布頓大麥根的維管束面積、增加導管直徑、表皮厚度和內皮層厚度,說明堿脅迫條件下,Epichlo?內生真菌的侵染對宿主細胞起到一定的保護能力,提高了宿主植物對水分和無機鹽的吸收能力,并減少水分的散失。
植物通過木質部將根部吸收的水分和礦物質往上運輸到地上部各營養器官,地上部光合作用的產物則通過韌皮部向下運輸至地下部,并將葉片產生的有機物質運送到根內或暫存莖內[26]。據報道,堿脅迫導致草木犀(Melilotusofficinalis)莖稈側面的維管束大量減少和上、下表皮厚度增加[20]。本研究對E+和E-植株的莖在不同濃度堿脅迫下的顯微結構進行了觀察,發現莖中維管束與髓部薄壁細胞都發達。在堿脅迫下,莖通過表皮厚度、維管組織髓結構的改變來適應環境的變化。莖的表皮厚度隨著堿濃度增大逐漸加厚,說明植物通過增加表皮厚度減少機體水分的散失。同時,本研究發現在0 mmol·L-1和100 mmol·L-1E+植株的氣腔直徑顯著高于E-,因氣腔主要功能是有利于器官中的細胞呼吸時的氣體交換,所以我們推測Epichlo?內生真菌對宿主植物在堿脅迫時細胞呼吸有促進作用。
葉是植物進行光合作用和蒸騰作用的主要器官,因結構特征的變化最能體現植物對環境的適應[27],所以研究最多的器官是葉[28-29]。葉脈在水分和養分的運輸中發揮著重要的生理作用[30]。在堿脅迫下草木犀小葉的上、下表皮細胞壁增厚,減少了葉片非氣孔路徑的水分流失以及在200 mmol·L-1堿濃度處理后,木質部導管直徑減少了47 %,增加了蒸騰阻力[20]。ARAFA等[31]發現,較高水平的鹽脅迫(3 000和6 000 ppm NaCl)降低了高粱木質部和韌皮部組織厚度、葉片厚度、維管束尺寸以及后生木質部導管直徑。本研究通過觀察堿脅迫下E+和E-植株葉片顯微結構的形態變化,發現隨著堿濃度的增加葉片維管束面積變小,導致水分、無機鹽和有機物質的輸送能力減弱,使得葉肉細胞得不到充足的養分,所以在顯微結構上表現出葉肉細胞體積變小。泡狀細胞與葉的卷曲和伸開的作用有關,在水分吸收和保持細胞水勢中起一定的作用,本研究發現堿脅迫使泡狀細胞下陷較深,面積變大,使得在缺水時快速將葉片卷曲,減少水分的散失,這與鹽脅迫下小花堿茅(P.tenuiflora)葉片泡狀細胞的研究結果一致[32],但E+植物較E-植株泡狀細胞下陷更深,面積變大,表明堿脅迫下Epichloё內生真菌能有效的防止布頓大麥葉肉細胞內水分的丟失。氣孔直徑的變化可以看出E+的先升后降,但堿處理后E+的氣孔直徑始終大于0 mmol·L-1時的氣孔直徑;E-無顯著影響,說明Epichloё內生真菌具有促進布頓大麥氣體交換能力。
4 結論
在堿脅迫下,內生真菌的侵染影響了布頓大麥的顯微結構。堿脅迫下Epichloё內生真菌的侵染提高了根的維管束面積、增加了導管直徑、表皮厚度和內皮層厚度,提高了植物對水分和無機鹽的吸收能力,增加對細胞的保護和減緩水分的散失;Epichloё內生真菌增加了莖中氣腔直徑、表皮厚度和促進維管組織中髓的發育;同時,葉的泡狀細胞面積和氣孔直徑變大,使得葉片的失水能力減弱;從而有效的防止布頓大麥葉肉細胞內水分的丟失,促進氣體交換能力。本研究從形態結構方面證明了堿脅迫下Epichloё內生真菌提高了宿主布頓大麥的耐受性。
