模擬放牧采食對典型草原土壤中AM真菌分解枯落物的影響
2023-03-05 14:03:32趙心雨田麗華趙學春桂維陽
草地學報 2023年2期
趙心雨, 田麗華, 趙學春, 陳 超, 桂維陽
(貴州大學動物科學學院, 貴州 貴陽 550025)
枯落物是天然草原碳庫的主要組成部分之一,是草地生態系統物質和能量的重要來源[1-2]。枯落物經礦化作用分解后或被植物吸收利用,或以CO2的形式返回至大氣中,在維持草地生態系統碳平衡中起到關鍵作用[2-3]。
草地生態系統中的大部分碳庫位于土壤中[4]。典型草原中80%的植物可以和叢枝菌根真菌(Arbuscular mycorrhizal fungi,AMF)形成共生體,其中AMF菌絲是地下碳庫的重要載體之一[5]。大多數研究認為AMF并不具有腐生能力,因此AMF參與枯落物分解的過程主要通過影響其與土壤微生物之間的互作關系,改變土壤微生物對酶的分泌,調控枯落物的分解率[6]。有研究發現,AMF會通過分泌易分解有機質刺激土壤微生物活性,加快枯落物的分解[7]。Cheng等研究發現,土壤中的AMF被隔離后,枯落物的分解減少,土壤微生物的活性下降[8]。此外,AMF還會與土壤微生物競爭養分,進而刺激或抑制微生物的活性。Jannoura等發現,由于土壤中氮素的缺乏,豆科植物接種AMF后,微生物活性受抑制,導致枯落物—玉米(Zeamays)的分解率降低[9]。
水解酶和氧化酶是土壤微生物降解復雜有機化合物的重要物質,可以反映土壤生化過程的大小和趨勢[10]。纖維素是植物殘體進入土壤的碳水化合物的重要組分之一,是一種含碳量較高的化合物,結構相對簡單,分解較快[11]。纖維素二糖水解酶在參與纖維素的分解過程中可將纖維素分解為纖維二糖[12]。β-葡糖苷酶的主要作用是參與分解纖維素類多糖的水解過程,裂解β-葡糖苷和多聚糖形成葡萄糖,其作用產物是土壤微生物重要的營養源[13]。氧化酶中起主要作用的為多酚氧化酶,其可以促進腐殖質的形成,分解枯落物中的木質素,進一步將酚類物質氧化為醌[14]。有研究發現,AMF的生長會促進土壤酶的活性,例如β-葡糖苷酶、磷酸酶和脲酶等[15]。然而,較少有研究系統調查AMF是否通過調控土壤酶活性影響枯落物的分解。
植物功能群會影響土壤微生物的群落結構和功能[16]。不同植物功能群的地上凈初級生產力并不一致,這會改變植物對土壤微生物群落的碳分配,從而使微生物的活性受到干擾[17]。植物群落結構的改變會影響土壤的固碳能力,還會影響土壤微生物碳源(根系分泌物)的多樣性,植物與土壤微生物之間的相互關系以及土壤微環境的多樣性等[18-19]。并且,很多研究均指出不同植物功能群對AMF的依賴性表現差異較大,其中C4禾草相比于C3禾草及固氮雜草對AMF的依賴性更大,更易于形成共生體[20]。但植物功能群對AMF參與枯落物分解的影響卻鮮有報道。
植物的光合作用可以為地下部分輸送大量碳源[21]。因此,宿主植物光合作用的削弱,例如植物地上部分的刈割,會限制宿主植物為其共生的AMF的碳分配[22]。因此,刈割會通過調控宿主植物共生的AMF來影響土壤微生物群落,進而降低枯落物的分解率[23]。放牧是草地生態系統的主要土地利用方式之一,會對植物功能性其家畜采食作用中采食量的部分可以通過刈割處理代表,用于探究刈割對土壤微生物的影響,有助于我們預測草地生態系統中由放牧導致的土壤碳流轉的變化。放牧過程中家畜采食過程會改變地上植被的豐富度和多樣性,影響根際微生態環境[24-25]。
前人對AMF參與枯落物的分解主要在溫室和盆栽試驗中進行,AMF和土壤微生物之間的互作受控制試驗的條件限制,難以展現其在野外原位環境中的真實性[26]。本試驗采用網袋法,使用兩種不同孔徑的尼龍網(隔離植物根系但允許AMF菌絲通過、同時隔離植物根系和AMF菌絲),并通過測量枯落物的分解率、土壤酶活性,以闡明在典型草原中,放牧的采食作用(植物功能群和刈割處理)對AMF參與枯落物分解過程的影響。
本研究提出3個假設:(1)不同植物功能群處理中枯落物的分解率有差異;(2)AMF會通過影響土壤酶活性改變枯落物的分解;(3)刈割會抑制枯落物分解。
1 材料與方法
1.1 研究區概況
研究區位于內蒙古多倫縣中科院植物所十三里灘恢復草地生態系統試驗示范基地(42°02′N ~ 116°17′E、海拔1 324 m)。氣候為溫帶內陸型季風氣候,年降水量為385.5 mm,年均溫為2.1℃。研究區土壤為栗鈣土,肥力低,結構疏松。植被類型為溫帶典型草原,主要植被:大針茅(Stipakryroii)、羊草(Leymuschinensis)、糙隱子草(Cleistogenessquarrosa)、冷蒿(Artemisiafrigida)等。
1.2 試驗設計
本研究采用隨機區組試驗設計,選取研究區地勢開闊、平坦且植被均勻的位置設立試驗。選擇植物功能群和刈割兩種利用方式模擬放牧中的采食作用(采食種類和采食量),其中植物功能群處理:C3禾草、C4禾草、雜類草和對照(CK),共4水平。刈割處理:刈割和不刈割,共兩水平,五個區組,共40個小區。小區面積:1.5 m × 1.5 m樣方,間隔:2 m。
每種植物功能群處理僅保留本功能群植物,其余植物地上部分全部使用剪刀剪去,使地下部分凋萎死亡但不破壞土壤結構。試驗于2015年5月1日開始,每隔兩周剔除一次,10月1日結束。刈割為每月一次(5—10月),留茬5 cm,屬于中度放牧強度。
1.3 測定項目及方法
1.3.1枯落物分解袋 2014年10月中旬,在研究區內用剪刀齊地面采集當地優勢物種羊草。挑選植株長度在15~20 cm,4~5片葉子的羊草立枯體,以保證材料質量的均勻度,切碎成1 cm左右的小段備用。
枯落物分解袋使用兩種規格尼龍網制成,分別為0.45 μm(完全隔離AMF菌絲和植物根系)和25 μm(僅隔離植物根系允許AMF菌絲通過)。分解袋長10 cm、直徑 6.5 cm(見圖1),袋內裝原位土150 g和羊草立枯物2 g,每種孔徑各小區均有3個重復,混合均勻后回填,填埋時間為每年5月1號,每年10月1號取樣。
枯落物分解袋取出后,將袋內的剩余枯落物小心取出,經清洗后烘干稱重。袋內的土壤用于測定土壤酶活和AMF菌絲密度。
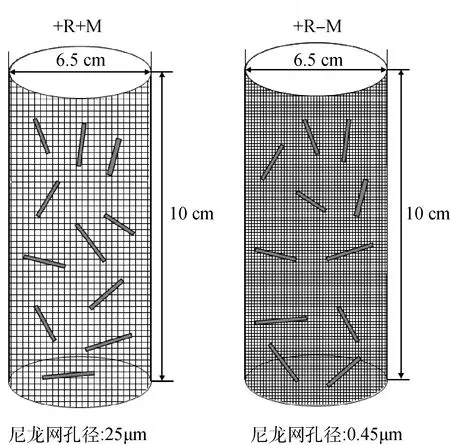
圖1 凋落物分解袋示意圖Fig.1 Litter decomposition bag
1.3.2AMF菌絲密度測定方法 AMF菌絲密度的測定使用真空泵微孔抽濾法[27]。取5.000 g風干土壤于1 L燒杯中,加入1 L去離子水和攪拌子,置于磁力攪拌器上攪拌5 min以使菌絲充分脫落,從而形成土壤懸濁液;將此懸濁液靜置至無明顯漩渦后過雙層篩,上篩為20目,下篩為400目,過濾土壤懸濁液;再向燒杯中加水1 L后重復過濾一次,保證燒杯中所有土壤全部轉移至篩中;上篩中的雜質丟棄,下篩中的土用約100 mL水沖洗至攪拌機中,攪拌10 s,停止5 s,再攪拌20 s;攪拌后的懸濁液全部轉移至250 mL三角瓶中,用去離子水定容至250 mL,置于220 r·min-1搖床中劇烈搖晃后,靜置1 min;使用移液器在同一液面下1 cm處吸取5 mL懸濁液,轉移至放有0.45 μm濾膜的抽濾裝置的過濾杯中,進行真空抽濾;抽濾后將濾膜轉移至載玻片,滴數滴0.05%曲利苯藍溶液染色,蓋上蓋玻片后靜置20 h;重復“劇烈搖晃”和“抽濾”4次,即每個土壤樣品制4片濾膜。使用“100 ×”倍數下用顯微鏡(CX41,Olympus,Japan)觀察并計數。使用網格交叉法估算菌絲長度,隨機觀察圓形濾膜上25個視野,每個視野的交叉點數為一根菌絲與十字交叉格橫豎方向上的全部點數。
菌絲密度計算公式:
菌絲長度(m)=11/14×總交叉點數×網格單元格長度×濾膜上樣塊面積/網格面積
菌絲密度(m·g-1)=菌絲長度/所稱土的質量
1.3.3土壤酶活性測定方法 試劑配制:
(a)醋酸鈉緩沖液:配制50 mmol·L-1的醋酸鹽溶液(將6.804 g三水合乙酸鈉溶解在800 mL去離子水中),再使用12 mol·L-1的鹽酸將緩沖液調節pH至5.0后保存在4 ℃冰箱,需在8天內使用完畢。
(b)甲基傘型酮(MUB)及酶底物(表1):配制200 μmol·L-1的底物和10 μmol·L-1MUB,提前一天配制,放置在4oC冰箱保存,3天內使用完畢。
測量方法:
(1)稱取1.00 g鮮土于250 mL錐形瓶中,加入125 mL的醋酸鈉緩沖液,使用磁力攪拌器(攪拌子長度為3.5 cm)使土壤成均一懸濁液,后置于往復式搖床中震蕩1 h(200 r·min-1);
(2)用八通道移液器吸取200 μL醋酸鈉緩沖液于黑色96微孔板上,分別是空白對照,標準熒光,底物對照,依次加入50 μL緩沖液、10 μmol·L-1-MUB和200 μmol·L的底物;
(3)用八通道移液器吸取200 μL土壤懸濁液于黑色96微孔板上,每一個土壤樣品設置8個重復,并設置樣品對照和標準對照;吸取50 μL的200 μmol·L-1的底物,加到96孔板對應的樣品池里,樣品對照加50 μL緩沖液,標準對照加50 μL 10 μmol·L-1的MUB。
(4)點樣結束后,立即20oC暗光培養2 h,培養結束后添加10 μL 1 mol·L-1的氫氧化鈉溶液終止反應;
(5)使用多功能酶標儀(SpectraMax i3x,Molecular Devices,CA,USA)在365 nm激發光,450 nm透射光下測定熒光值(DeForest,2009),土壤酶活性計算過程如下:
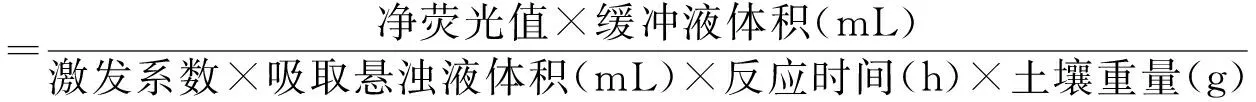


表1 土壤酶種類及其縮寫和酶作用底物Table 1 Enzymes measured in soil samples,and their substrates and abbreviation
1.4 數據統計與分析
采用SAS 8.0(SAS Institute,USA)對各指標進行三因素方差分析(Three-way ANOVA)。結果顯示刈割對各指標的影響均不顯著,故將刈割處理合并為重復(附表1),再使用雙因素方差分析(Two-way ANOVA)進行檢驗,多重比較方法使用Duncan檢驗。使用一元線性回歸分析(Linear regression)AMF菌絲密度與枯落物分解率的相關性。
2 結果與分析
2.1 枯落物分解率和AMF菌絲密度
兩年間,植物功能對枯落物分解率有顯著影響,且變化趨勢相似(圖2,2016:P<0.001;2017:P<0.001)。雜類草和CK顯著高于C3禾草處理,C4禾草處理最低。兩年中,AMF均促進了枯落物的分解(圖2a,2b,P<0.001)。
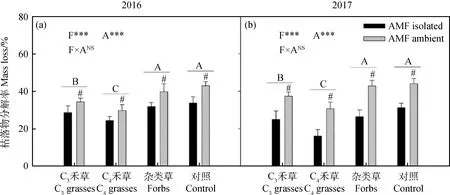
圖2 2016年(a)和2017年(b)植物功能群處理(F)和AMF處理(A)對枯落物分解率的影響Fig.2 The effect of AMF Treatment (A) on mass loss of different plant functional groups (F) in 2016 (a) and 2017 (b)注:不同字母表示植物功能處理中差異顯著。#表示在植物功能群處理內差異顯著。*=P<0.05;**=P<0.01;***=P<0.001;NS=P>0.05Note:Different letters indicate significant differences among the plant functional groups. #that a significant difference between the treatments of AMF isolated or not within one plant functional group. *P<0.05;**P<0.01;***P<0.001;NSP>0.05. The same as below
兩年間,植物功能群對AMF菌絲密度有顯著影響,但趨勢并不一致(圖3,2016:P<0.01;2017:P<0.01)。2016年,雜類草和CK較高,C3和C4禾草處理較低;2017年,雜類草顯著高于C3和C4禾草處理。2017年,AMF菌絲密度在各功能群中均呈顯著差異,說明尼龍網在所有處理中都顯著阻隔了AMF菌絲的生長(圖3,P<0.05)。
2.2 土壤酶活性
兩年間,β-葡糖苷酶在不同植物功能群中呈顯著差異(圖4a,4d:P<0.001)。2016年,雜類草和CK處理較高,C3和C4禾草處理較低;2017年,C4禾草顯著低于其余三種處理。2016年,AMF僅對C3禾草中β-葡糖苷酶活性有顯著影響(圖4a,P<0.05)。
2017年中,纖維素二糖水解酶在不同植物功能群間呈顯著差異,雜類草高于C3禾草和CK處理,C4禾草處理呈最低值(圖4e,P<0.05)。
兩年間,多酚氧化酶在不同植物功能群中呈顯著差異(圖4c,4f,2016:P<0.001;2017:P<0.001)。2016年,雜類草和CK處理較高,顯著高于C3和C4禾草處理;2017年,多酚氧化酶活性在雜類草和CK處理中最高,C3禾草較低,最低為C4禾草處理。2016年,AMF顯著提高除C3禾草外其余處理中的多酚氧化酶活性(圖4c,P<0.05);2017年,AMF顯著提升了所有處理的多酚氧化酶活性(圖4f,P<0.05)。
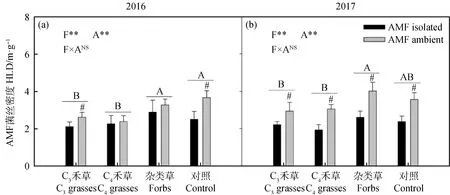
圖3 2016年(a)和2017年(b)植物功能群處理(F)和AMF處理(A)對AMF菌絲密度(HLD)的影響Fig.3 The effect of AMF treatment (A) on extra-radical mycorrhizal hyphal length (HLD) among different plant functional groups (F) in 2016 (a) and 2017 (b)注:不同字母表示植物功能處理中差異顯著。#表示在植物功能群處理內差異顯著。*P<0.05;**P<0.01;***P<0.001;NSP>0.05Note: Different letters indicate significant differences in plant functional treatment. #indicates that there is significant difference in plant functional group treatment. * P<0.05;**P<0.01;***P<0.001;NS P>0.05
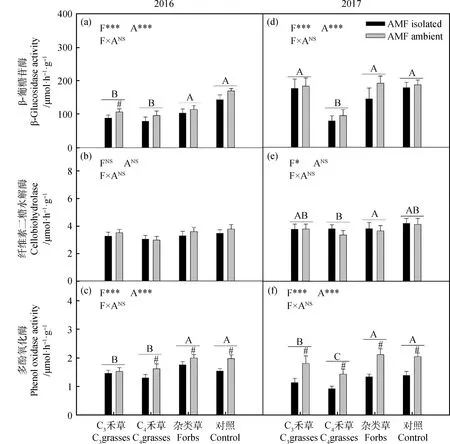
圖4 2016年(a)和2017年(b)植物功能群處理(F)和AMF處理(A)對土壤酶活的影響Fig.4 The effect of AMF treatment (A) on enzymes activity among different plant functional groups (F) in 2016 (a) and 2017 (b)注:不同字母表示植物功能處理中差異顯著。#表示在植物功能群處理內差異顯著。*P<0.05;**P<0.01;***P<0.001;NSP>0.05Note: Different letters indicate significant differences in plant functional treatment. #indicates that there is significant difference in plant functional group treatment. * P<0.05; ** P<0.01; ***P<0.001; NS P>0.05
2.3 AMF菌絲密度與枯落物分解率的關系
兩年間,AMF菌絲密度與枯落物分解率間均有顯著的線性正相關關系(圖5a,5b,2016年:P=0.033 1;2017年:P=0.006 6)。
3 討論
本試驗使用網袋法,通過適宜孔徑的尼龍網可以成功隔離AMF菌絲,顯著降低了0.45 μm網袋內的AMF菌絲密度,通過比較枯落物分解率、土壤水解酶活性和氧化酶活性,在天然草地探究了植物功能群和刈割對AMF參與枯落物分解的影響,明確AMF促進枯落物分解的可能原因。試驗結果發現,2016和2017兩年中AMF根外菌絲的生長均會加速枯落物的的分解,且不同功能群處理之間的差異顯著,雜類草處理中的枯落物分解率顯著高于C3禾草處理,C4禾草處理中枯落物分解率最小。枯落物的分解對刈割的響應不顯著。使用線性回歸分析發現,兩年間AMF菌絲密度和枯落物分解率之間均有顯著的正相關關系。本研究說明AMF會通過調控土壤酶活促進枯落物的分解,且植物功能群和刈割處理不會影響AMF對枯落物的分解作用。
Leifheit等發現,AMF通過改變土壤微環境中的養分和水分條件可以直接影響土壤中的微生物(分解者)活性,或通過為微生物提供或改變碳源間接影響微生物活性,進而影響土壤中的有機質分解[28]。與Armada等試驗結果類似,本試驗結果發現AMF的生長會促進土壤中β-葡糖苷酶的活性[15]。Toljander等研究得出,AMF根外菌絲的分泌物中包含有低分子量糖類,可被β-葡糖苷酶快速分解,為土壤中的其他微生物提供碳源,刺激其活性加快枯落物分解[29]。另外,AMF會顯著增加土壤中的多酚氧化酶,說明AMF的生長會促進土壤中其他微生物上調對多酚氧化酶的分泌。天然草地的羊草中,木質素含量相比與纖維素含量較少,但卻是枯落物中最難以分解的復合物。木質素的分解過程中起主要作用的是多酚氧化酶,此酶主要由土壤中的白腐菌分泌,很多研究發現,當土壤中氮素缺乏時,會抑制土壤中其他微生物與白腐菌之間的競爭作用,從而使得白腐菌活性上升[30]。AMF會吸收土壤中的氮素并將其轉運給宿主植物,所以AMF的生長會加劇土壤微環境中氮素的匱乏,進而刺激土壤微生物(白腐菌等)的活性,以分泌更多的多酚氧化酶,促進枯落物中木質纖維素的分解。但AMF的出現對土壤中纖維素二糖水解酶活性無顯著作用,說明AMF對枯落物中纖維素的分解效果不明顯。土壤中大量的細菌和真菌均可以分泌纖維素二糖水解酶,故其活性的控制因子復雜較難以被單一因素影響;并且AMF獲取碳源的主要方式是跟與其共生的宿主植物進行養分交換,并不需要從土壤中直接獲取碳源用于自身生長[31]。所以枯落物纖維素的分解率并不受AMF控制,但可能會是其他環境因子。
植物功能群處理之間對枯落物分解率的影響有顯著差異,主要是因為不同植物功能群對碳分配的策略不同[32]。C4禾草處理中物種多樣性較低,僅糙隱子草(Cleistogenessquarrosa)一種,且這種植物株型矮小生物量低。2016和2017年中,C4禾草的地上和地下生物量均較其他功能群植物更少,導致與植物共生的AMF的菌絲密度較小,對土壤酶活的激發程度低。且C4禾草處理中的蓋度相比其他功能群植物更少,導致地表裸露面積較大,會接受更多的太陽輻射,使土壤水分蒸發效率上升,抑制土壤微生物的活性,進而減少了對枯落物的分解。而雜類草和CK(多樣性較高)處理中,土壤酶活性顯著高于其他植物功能群處理。前人已有研究得出,植物多樣性的增會促進土壤微生物生物量的升高,因為植物多樣性高的群落會為土壤微生物提供更為豐富的碳源[33]。且2016和2017兩年中雜類草和CK處理中地下生物量較高,表明其會為地下部分提供更多的碳,增加微生物的數量,促進土壤酶活性,加快枯落物的分解。
本研究發現,刈割對枯落物的分解無影響。Ren等研究發現,放牧過程中僅7%的土壤微生物變異與放牧過程本身有關,而89%的變異更可能來自于其他因素,說明微生物群落變化與土地管理方式引起的環境變化關系可能更密切,而不是放牧中的采食過程[34]。適度刈割會激發土壤微生物的活性[35]。但是,這也取決于時間尺度[36],本研究僅有三年跨度,刈割對枯落物分解的影響需要更長時間跨度才會顯示。
4 結論
通過植物功能群和刈割處理模擬放牧過程中家畜采食作用,探究此過程中AMF對枯落物分解的影響。結果發現AMF主要通過提升土壤β-葡糖苷酶和多酚氧化酶活性,加速枯落物的降解。不同植物功能群分解枯落物的能力不同,其中雜類草處理分解枯落物的能力更強,而C4禾草處理中的枯落物分解率最低。AMF和植物功能群處理無顯著交互作用,表明植物群落組成的變化并不會改變AMF對枯落物的分解作用。刈割對枯落物的分解在短時間內無顯著影響。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
