親子依戀與兒童抑郁癥狀的關系:兒童對環境的生物敏感性的作用及父母差異*
2023-03-07 13:51:18徐健捷張一一林德堃車俐穎宋漫漫
心理學報 2023年3期
徐健捷 張一一 林德堃 車俐穎 宋漫漫 韓 卓
親子依戀與兒童抑郁癥狀的關系:兒童對環境的生物敏感性的作用及父母差異*
徐健捷 張一一 林德堃 車俐穎 宋漫漫 韓 卓
(北京師范大學心理學部, 北京 100875)
不良的親子依戀是導致兒童抑郁癥狀的風險因素, 但其對兒童的影響會因兒童對環境的生物敏感性不同而存在差異。為探究家庭中母子、父子依戀對兒童抑郁癥狀的影響及兒童生物敏感性(本文中采用迷走神經抑制作為指標)在其中的作用, 本研究結合行為任務、問卷報告、生理測量等多種研究手段, 招募150名學齡兒童(平均年齡8.64歲, 63名女孩)參與研究。結果表明:(1)學齡兒童的母子依戀水平高于父子依戀水平。(2)高水平的母子依戀與父子依戀會同等程度地降低兒童的抑郁癥狀。(3)兒童對環境的生物敏感性對母子依戀影響兒童抑郁癥狀路徑的調節作用顯著, 生理上對環境更敏感(高迷走神經抑制)的兒童更易得益于高母子依戀, 表現出較低的抑郁水平; 但同時, 這類兒童在母子依戀較低時也更易表現出較高的抑郁水平。(4)兒童的生物敏感性對父子依戀影響兒童抑郁癥狀路徑的調節作用不顯著, 高父子依戀對生物敏感性水平不同的兒童均存在有利影響。(5)在不同情境中測量的迷走神經抑制對親子依戀影響兒童抑郁癥狀的調節作用模式相似, 表現出跨情境一致的特點。本研究率先揭示兒童迷走神經抑制與親子依戀對兒童抑郁癥狀的聯合作用機制及父母角色差異。
親子依戀, 兒童抑郁癥狀, 生物敏感性, 父母差異
1 引言
抑郁是指人們在日常生活中較為持久的消極思維體驗(Teasdale, 1983), 在我國兒童中存在較高的流行率, 超過了10% (沈彥等, 2021)。若不進行有效干預, 會對兒童的學習生活、甚至青少年期和成年期的心理社會功能造成嚴重影響(Copeland et al., 2021; 姚文玉等, 2021)。家庭是兒童發展最重要的環境, 與父母關系疏離、未形成良好的依戀關系可能是兒童抑郁癥狀產生的一個重要原因(Shuang et al., 2022)。進一步地, 個體對環境的生物敏感性模型(Boyce & Ellis, 2005)指出, 兒童發展結果被環境影響的程度可能受其自身某些生理特質的影響:對于具有某些生理特質的兒童(例如, 在壓力情境下產生較強的迷走神經抑制), 他們可能在更多受益于積極環境的同時, 也更多被消極環境所損耗。相反, 對于不具有這些生理特質的兒童, 環境對其發展結果的影響有限。因此, 探究親子依戀與兒童抑郁癥狀間的關系時有必要考慮兒童自身對環境的生物敏感性。
改革開放以來, 我國經濟社會飛速發展, 家長的養育觀念、親子依戀關系及其對兒童的影響程度也在隨之改變(Chen et al., 2021)。例如, 相比于上世紀的家長, 當代中國母親與兒童互動時所表現出的親密和依戀程度有所下降(Chen et al., 2021); 而父親更少地扮演家庭中“權威”的角色, 與兒童有更多的親密互動(Li, 2020)。然而, 在當今家庭中, 兒童與父母的依戀程度及其對兒童抑郁癥狀的影響如何以及上述關系是否會因父母角色的不同而產生差異尚不清楚。因此, 本研究擬探討母子、父子依戀對不同生物敏感性兒童抑郁癥狀的影響, 以期揭示親子依戀影響兒童抑郁癥狀的特點及邊界條件, 從家庭研究的視角為兒童心理健康問題的預防和干預提供重要理論基礎和實證證據。
1.1 母子依戀、父子依戀與兒童抑郁癥狀
布朗芬布倫納的生物生態學模型(bioecological model)強調“近端過程” (proximal process; 個體在其最直接的生活環境中與周圍人和物的交互)是個體發展最有力的影響因素(Bronfenbrenner & Morris, 1998; Mer?on-Vargas et al., 2020)。家庭是兒童發展過程中最重要的場所(Eisenberg et al., 1998)。相應地, 在家庭系統中與父母之間的互動會對兒童發展產生至關重要的作用(Cox & Paley, 1997)。個體出生后與父母在不斷的互動中逐漸建立起一種深層持久的情感聯結, 形成了親子間的依戀(Ainsworth & Bowlby, 1991)。與影響兒童發展的其他家庭因素(如家長自身特質、父母教養方式等)相比, 兒童感知到的親子依戀是兒童發展更直接、更深層次的預測因素(趙鳳青等, 2022)。經典依戀理論指出, 與父母間的依戀會影響兒童對自己和他人進行感知和評價的內部工作模式:安全穩定的依戀關系會讓兒童認為自己是有價值的并且他人是可信任的; 然而, 若沒有建立安全穩定的依戀關系, 則會形成消極的內部工作模式, 產生對自我、他人的負性心理表征, 并進一步發展出一系列消極的情感、認知和行為方式, 導致抑郁相關癥狀(Ainsworth & Bowlby, 1991; 趙鳳青等, 2022)。大量研究表明, 較低的親子依戀水平是兒童抑郁癥狀產生和發展的風險因素(Spruit et al., 2020)。
依戀理論進一步指出, 兒童存在多重依戀的現象, 即可以與母親和父親建立不同特征的依戀關系(Ainsworth, 1989)。母子、父子依戀的特征雖然有所差異, 但也相互補充, 共同影響兒童發展(王曉蕾等, 2018)。雖然近年來隨著社會文化變遷, 不少父親逐漸承擔更多家庭責任, 但受傳統性別角色分工影響, 母親仍會為孩子提供更多生活上的照顧(Li, 2020; 侯芬等, 2018)。與之相應, 目前學界大部分研究者認為母子依戀的水平仍然高于父子依戀(Kwon & Elicker, 2012)。然而, 相對較低的依戀水平并不意味著父子依戀對兒童抑郁癥狀的影響會弱于母子依戀, 父親在與兒童的互動中多以進行游戲、設置挑戰的方式為兒童帶來快樂和自信, 對兒童的心理健康發展發揮著不可替代的作用(Amodia-Bidakowska et al., 2020)。然而, 至今學界關于母子、父子依戀對兒童發展的影響孰強孰弱仍存在爭議。主導性假說(Suess et al., 1992)認為, 母親是家庭活動的主導者和孩子的主要養育者, 母子間更多的互動意味著母子依戀會對兒童的心理發展結果起主導作用(Murphy et al., 2017)。特異性假說(Howes & Ritchie, 1999)則指出, 母子、父子依戀都會對兒童發展產生獨特的影響, 并不存在主次之分(Lunkenheimer et al., 2020)。此外, 關系親密度與角色義務的關系理論(McAdams & Bauer, 2004)認為, 母子依戀由于大多維持在較高水平, 兒童已經習以為常, 而父親養育孩子的角色義務少于母親, 使得兒童更加珍視與父親的情感聯結, 父子依戀對兒童的影響程度會高于母子依戀影響的程度(García et al., 2014)。然而, 由于以往有關親子依戀的研究對父子依戀關注不足, 上述彼此對立的假說并沒有足夠的實證證據支持。基于此, 提出本研究的第一個研究問題:母子依戀與父子依戀對兒童抑郁癥狀的影響程度有何差異?
1.2 兒童對環境的生物敏感性的調節作用
親子依戀與兒童抑郁癥狀在整體上存在著較強的關聯, 但并不意味著該關系適用于所有兒童。兒童發展的差別易感性模型(differential susceptibility model; Belsky, 1997)指出, 具有“易感性”特質的兒童表現出對父母養育的敏感性, 即在積極的養育環境(如安全的親子依戀)下, 易感性高的兒童會發展得更好, 但在消極養育環境下(如不安全的親子依戀), 易感性高的兒童會發展得更差, 更有可能產生抑郁等心理行為問題。進一步地, 對環境的生物敏感性模型(biological sensitivity to context model; Boyce & Ellis, 2005)聚焦兒童自身的神經生理反應差異, 從生物行為學的角度對差別易感性模型的作用模式進行闡述。具體來說, 該模型強調要通過測量兒童生理系統對外部事件的反應程度來作為兒童對環境的生物敏感性(以下簡稱“兒童生物敏感性”)指標, 生物敏感性水平高的兒童更易從積極養育環境中獲益但也更易在消極養育環境中受損。
迷走神經抑制(vagal suppression)是衡量兒童生物敏感性的一個常用指標, 是指在面對挑戰或壓力事件時兒童迷走神經活動水平的減弱程度, 可通過呼吸性竇性心律不齊(respiratory sinus arrhythmia, RSA)的變化來測量(Porges, 2007)。RSA是指在包含吸氣和呼氣的一個呼吸周期中特定頻段的心率變異程度(heart rate variability), 反映了迷走神經對心臟活動的影響(Porges, 2007)。在靜息狀態下, 迷走神經活動水平較高, 對心臟活動的抑制程度較強, 反映在呼氣時心跳速度明顯低于吸氣時的心率水平, 心率變異性較大, 即個體RSA水平較高, 這體現了個體維持內穩態的能力(Porges, 2007)。當遇到外部壓力事件時, 為了調動機體有效應對壓力, 迷走神經的活動水平會減弱(即迷走神經抑制), 對心率抑制作用降低, 使得個體的心跳加快, 心率變異性減小, RSA水平與靜息狀態相比會下降(Porges, 2007)。迷走神經活動水平在壓力或挑戰任務中相比于靜息狀態下降的程度(即迷走神經抑制程度)體現了個體迷走神經活動對外部事件的反應程度。迷走神經抑制程度越高意味著個體的生物敏感性水平越高(Cipriano et al., 2011)。
上述理論觀點得到了實證研究的支持。一項基于中國兒童的研究探討了壓力任務中迷走神經抑制程度作為生物敏感性指標對父母情緒教養影響兒童焦慮癥狀的作用(Han et al., 2020)。結果表明, 對于高迷走神經抑制的兒童(即高生物敏感性兒童),高支持型的父母情緒教養能夠降低兒童的焦慮癥狀水平; 但高支持型父母情緒教養對低迷走神經抑制兒童(即低生物敏感性兒童)的焦慮癥狀則不能起保護作用(Han et al., 2020)。此外, 國外兒童群體中也發現了類似的結果。例如, Cipriano等人(2011)發現, 在挑戰性任務中迷走神經抑制程度高的兒童對家庭暴力環境非常敏感, 隨著家庭暴力水平的提高, 高迷走神經抑制兒童的情緒問題也會隨之提升;與之相對, 家庭暴力對低迷走神經抑制兒童的情緒問題沒有明顯的影響。類似的, 研究者還發現, 與迷走神經抑制程度低的兒童相比, 高迷走神經抑制的兒童若處在消極的家庭環境中, 會表現出更少的學業投入(Obradovi? et al., 2010)和更多的行為問題(Tabachnick et al., 2021), 而在相對積極的環境中則會有較少的學業和行為問題。與之相對, 生理上對環境不敏感的兒童并不會受到家庭環境的過大影響(Obradovi? et al., 2010; Tabachnick et al., 2021)。
然而, 雖然前人研究較為詳細地探討了迷走神經抑制作為生物敏感性指標對家庭環境影響兒童發展的調節作用, 但其生物敏感性指標大多僅通過單一的壓力或挑戰事件來測量(Han et al., 2020; Tabachnick et al., 2021)。有研究雖然采用了不同任務測量兒童的迷走神經抑制程度, 但并沒有關注不同情境下迷走神經抑制程度對家庭環境與兒童發展結果關系的調節作用是否相似, 而是直接將不同任務情境中迷走神經抑制程度求平均值作為整體的生物敏感性指標(Cipriano et al., 2011; Obradovi? et al., 2010)。僅有的少量關注不同情境迷走神經抑制作用的文章并未將其看作生物敏感性的指標, 而是考查不同任務中迷走神經抑制程度的差異(Zeytinoglu et al., 2020)或其對兒童發展結果直接預測作用的異同(Qui?ones-Camacho & Davis, 2019)。
由此可見, 目前尚缺乏研究細致探討作為生物敏感性指標的迷走神經抑制對家庭環境與兒童發展間關系的影響究竟是跨情境一致的還是情境特異的。迷走神經抑制只是一種客觀發生的生理現象, 其具體的心理學意義需要結合任務情境進行解釋(Davis et al., 2020)。然而, 目前的敏感性模型并沒有考慮不同情境中的敏感性與環境交互對兒童發展的影響是否有差異, 因此檢驗迷走神經抑制是否跨情境一致能夠使我們更加清楚地了解迷走神經抑制與家庭環境交互影響兒童發展的作用機制, 推動兒童發展的環境敏感性模型的理論進展。值得注意的是, 雖然以往研究也發現迷走神經活動的基線水平可以調節家庭環境與兒童發展結果間的關系(Mezulis et al., 2015), 但本研究關注的核心問題是不同情境中迷走神經抑制的作用究竟是特異的還是一致的, 因此本研究中的兒童生物敏感性聚焦于迷走神經的抑制程度而非基線水平。基于以上論述, 提出本研究的第二個研究問題:不同任務情境中測得的兒童迷走神經抑制是否均會調節親子依戀(包括母子依戀和父子依戀)對兒童抑郁癥狀的影響, 遵循對環境的生物敏感性模型的作用模式?
1.3 研究概覽
綜上, 雖然以往研究探討了親子依戀對兒童抑郁癥狀的影響, 但并沒有系統考查上述影響是否會因父母角色而產生差異, 也缺乏從實證研究角度對目前多個相互矛盾的理論假說進行檢驗。此外, 前人研究針對兒童生物敏感性在家庭環境影響兒童發展中的作用是跨情境一致還是情境特異的探討不足, 亟需實證研究來回答這一重要科學問題。鑒于此, 本研究結合問卷報告、行為任務和生理測量, 系統探究不同情境(社會壓力任務、消極情緒任務)中兒童對環境的生物敏感性(即迷走神經抑制)在母子父子依戀影響兒童抑郁癥狀中的作用模式及其中的父母角色差異。根據前人研究結論和相關理論, 擬提出如下假設:母子依戀與父子依戀均可以獨立地影響兒童抑郁癥狀, 且該影響會受到不同情境中兒童生物敏感性的調節, 即生物敏感性高的兒童既更易受到高親子依戀的積極影響, 也更易受到低親子依戀的消極影響。
2 研究方法
2.1 被試
本項目通過社區宣傳的方式, 招募學齡兒童及其家庭自愿參與研究, 共招募到150名兒童(= 8.64歲,= 1.67歲) 參與, 其中女生63名(占42.0%), 男生87名(占58.0%)。兒童由一位家長(= 39.22歲,= 4.07歲) 陪同來到實驗室, 其中母親121名(占80.7%), 父親29名(19.3%)。家長受教育程度從高中(占6.0%)到研究生以上(33.3%)。被試的家庭月收入從4000~4999元(0.7%)到20000元以上(40.9%)。
2.2 實驗流程
兒童和家長來到實驗室后, 實驗人員會首先向其介紹實驗目的和整體流程, 如果兒童和家長都同意參與實驗, 會進一步簽署知情同意書。之后, 兒童放松地坐在椅子上熟悉實驗環境。與此同時, 實驗人員會在兒童的雙側肋骨下方和右側鎖骨下方貼上粘貼3枚氯化銀一次性電極貼, 用來測量心電(electrocardiogram, ECG)。呼吸綁帶環繞在被試的胸腔上, 用來測量兒童的呼吸。待連接好儀器且被試適應完畢后, 依次進行靜息任務、社會壓力任務和消極情緒任務。所有任務中兒童的表現會被全程錄像, 兒童的生理活動會被全程記錄。任務結束后, 兒童會在實驗人員的指導下單獨填寫一系列問卷。在填寫問卷過程中, 主試會向兒童逐條朗讀各個條目, 并且澄清任何兒童提出來的疑問, 直到確保兒童清楚了解每個問題并做出相應回答。全部實驗流程結束后每個家庭會收到180元的被試費, 兒童會得到一個小禮物。研究得到了北京師范大學倫理委員會批準。
2.2.1 靜息任務
在靜息任務中, 兒童被要求安靜地坐在椅子上休息, 可以閉上眼睛, 放松身體, 盡量不要有大的動作。參考以往測量迷走神經抑制的相關研究(Tabachnick et al., 2021), 整個靜息任務持續2分鐘。
2.2.2 社會壓力任務
兒童會被告知需要參加一個演講任務, 演講的主題是“我最喜歡的科目”。主試進一步告訴兒童, 整個過程會全程錄像, 演講結束后會將錄像交給演講方面的專家進行分析和打分, 專家會仔細研究演講的內容、兒童的眼神和肢體動作等。為了進一步增加兒童的壓力程度, 兒童在演講的時候主試在一旁假裝傾聽記錄, 不給出任何表情和言語反饋。研究表明, 該范式能夠引發中國兒童較強的壓力和較大程度的迷走神經抑制(Han et al., 2020)。整個社會壓力任務持續4分鐘。
2.2.3 消極情緒任務
兒童被要求觀看一段視頻, 視頻的內容是一名小朋友在牙科診所治療的場景, 目的是引發兒童的負性情緒。該范式已被證明能夠引發兒童較強的消極情緒和較大程度的迷走神經抑制(Fortunato et al., 2013)。該任務持續2分半。
2.3 研究工具
2.3.1 母子依戀與父子依戀
采用Armsden和Greenberg (1987)編制, 金燦燦等(2010)修訂的父母與同伴依戀問卷(Inventory of Parent and Peer Attachment, IPPA)測量母子依戀與父子依戀。該量表共15個題目, 分為3個維度:信任、溝通和疏離, 每個維度各有5個題目。問卷采用5點計分, 從1 (完全不符合)到5 (完全符合)。兒童分別評價與母親(如“我會把自己的問題和煩惱告訴媽媽”)和父親(如“我會把自己的問題和煩惱告訴爸爸”)的依戀水平。將疏離維度的條目反向計分后, 所有條目得分求和分別得到母子依戀和父子依戀的總分, 得分越高, 表示母子或父子依戀程度越高。母子依戀與父子依戀量表的內部一致性信度良好, Cronbach’s α分別為0.76和0.81。采用Mplus 8.3對量表結構進行驗證性因素分析, 結果顯示, 母子依戀(χ2= 128.11,= 87, χ2/= 1.47, CFI = 0.90, RMSEA = 0.06, SRMR = 0.06)與父子依戀(χ2= 130.35,= 87, χ2/= 1.50, CFI = 0.92, RMSEA = 0.06, SRMR = 0.06)數據的模型擬合良好, 結構效度較為理想。
2.3.2 兒童抑郁癥狀
采用Radloff (1977)編制、陳祉妍等(2009)翻譯的流調中心抑郁量表(Center for Epidemiologic Studies Depression Scale, CES-D)測量兒童的抑郁癥狀水平。該量表共20個題目, 分為4個維度:軀體癥狀與活動遲緩(6題, 如“我覺得太累了, 不能做事情了”), 人際(2題, 如“我覺得人們不喜歡我”), 抑郁情緒(8題, 如“我覺得恐懼”)和積極情緒(4題, 如“我覺得開心”)。問卷采用4點計分, 從0 (沒有)到3 (總是)。兒童根據條目內容描述評定一周內該癥狀出現的頻率。將積極情緒維度的條目反向計分后, 所有條目得分求和, 得分越高, 表示抑郁癥狀水平越高。量表的內部一致性信度良好, Cronbach’s α為0.79。采用Mplus 8.3對量表結構進行驗證性因素分析, 結果顯示, 模型擬合良好:χ2= 218.86,= 158, χ2/= 1.39, CFI = 0.91, RMSEA = 0.05, SRMR = 0.06), 表明結構效度較為理想。本研究中兒童抑郁癥狀的得分在0~37分之間。根據劃界分標準(汪向東等, 1999), 71.7%的兒童無抑郁癥狀(15分及以下), 9.7%的兒童可能有抑郁癥狀(16~19分), 18.6%的兒童存在抑郁癥狀(20分及以上)。
2.4 生理測量
采用16導生理記錄儀(Biopac MP150)采集兒童在任務中的心電和呼吸數據, 采樣率為1000 Hz。對心電進行0.5到35 Hz的初步濾波后, 將其導入MindWare HRV 3.1.1進行計算。根據心率變異性的頻域計算指南, 提取在呼吸頻率范圍內心電信號的R波時間序列, 所有原始心電數據均由研究助理進行逐一檢查, 手動添加未識別的R波峰值以及剔除軟件誤識別的R波峰值。之后, 采用時段為30秒的漢明窗(Hamming window)函數, 通過快速傅里葉變換(fast fourier transform; FFT)將每一時段心動周期(heart period)的時間序列轉換為功率譜(power spectrum)。進一步地, 將兒童的高頻帶通濾波設定為研究者普遍采用的0.24~1.04 Hz (Cipriano et al., 2011; Fracasso et al., 1994; Han et al., 2020), 再將其進行自然對數轉換, 得到每個時段(30秒)的RSA值(單位為ln[ms2])。兒童在2分鐘的靜息任務中有4個RSA片段, 在4分鐘的社會壓力任務中有8個RSA片段, 在2分半的消極情緒任務中有5個RSA片段。
2.5 兒童生物敏感性(迷走神經抑制)
在獲得每個任務多個30秒RSA片段后, 將每個任務中的所有RSA片段進行平均, 分別得到靜息任務、社會壓力任務和消極情緒任務中RSA活動的平均水平。之后, 采用計算迷走神經抑制程度的經典方式(Qui?ones-Camacho & Davis, 2019), 將靜息任務下的RSA水平減去社會壓力任務或消極情緒任務中的RSA水平, 分別得到社會壓力任務和消極情緒任務中的RSA下降水平, 差值越大表明該任務相比基線RSA下降程度越大, 迷走神經抑制程度越高, 即兒童生物敏感性越高。
2.6 統計分析
采用SPSS 25.0進行描述統計、相關分析, 并通過配對樣本檢驗比較母子依戀與父子依戀的水平是否存在差異。采用Mplus 8.3通過路徑分析的方式進行模型檢驗。首先, 檢驗母子依戀和父子依戀對兒童抑郁癥狀的直接效應, 采用卡方差異檢驗比較母子依戀與父子依戀對兒童抑郁癥狀影響的強弱。進一步地, 分別檢驗社會壓力任務和消極情緒任務中兒童生物敏感性對父子、母子依戀影響兒童抑郁癥狀的調節作用, 并通過卡方差異檢驗比較兒童生物敏感性調節母子、父子依戀對兒童抑郁癥狀影響的強弱。若上述兩任務測得的兒童生物敏感性相關較高, 且對親子依戀影響兒童抑郁的調節作用模式相似, 則將兩任務中的兒童生物敏感性指標構建潛變量, 提取具有跨情境一致性的兒童生物敏感性指標。進一步采用潛調節結構方程法檢驗調節效應。缺失數據采用極大信息似然法(full information maximum likelihood estimation, FIML)處理, 該方法可以基于所有可得的信息(每個變量所匯報的最大被試量)對模型進行計算, 最大程度減小缺失數據帶來的估計偏差(Schlomer et al., 2010)。
若調節作用顯著, 根據敏感性領域研究者的建議(Roisman et al., 2012), 本研究進一步采用顯著性區間檢驗法(analysis of region of significance)檢驗調節作用是否符合對環境的生物敏感性模型, 采用如下3個判定標準:(1)確定環境變量(X, 親子依戀)的顯著性區間(region of significance, RoS):在環境變量 ± 2范圍內, 敏感性指標(Z, 即兒童生物敏感性)與因變量(Y, 即兒童抑郁癥狀)在交叉點兩側均存在顯著相關的區域, 即在親子依戀過低或過高時, 不同迷走神經抑制程度的兒童在抑郁癥狀水平上存在顯著差異。(2)交互作用比例(proportion of interaction, PoI):在自變量 ± 2范圍內, 交互效應圖中交叉點左右兩側直線間的面積比介于0.4到0.6之間。(3)受影響比例(proportion affected, PA):樣本中自變量得分大于交叉點對應分數的被試比例在16%到84%之間。若兒童生物敏感性對親子依戀影響兒童抑郁癥狀的調節作用同時滿足上述3個假設, 則表明調節作用的結果符合對環境的生物敏感性模型的標準。
3 結果
3.1 描述統計
剔除不在正負3個標準差范圍內的數據后, 對變量進行描述統計和相關分析(見表1)。相關分析結果表明, 母子依戀與父子依戀呈正相關(= 0.49), 社會壓力任務與消極情緒任務中的迷走神經抑制呈正相關(= 0.72), 靜息迷走神經活動水平與社會壓力任務(= 0.39)和消極情緒任務(= 0.44)中的迷走神經抑制呈正相關, 兒童抑郁癥狀與母子依戀(=?0.43)和父子依戀(= ?0.35)呈負相關。其他變量之間不存在顯著相關。此外, 除兒童年齡與兒童抑郁癥狀水平呈現負相關趨勢(= ?0.16,= 0.081)外, 其余控制變量與兒童抑郁癥狀相關均不顯著(s > 0.615)。
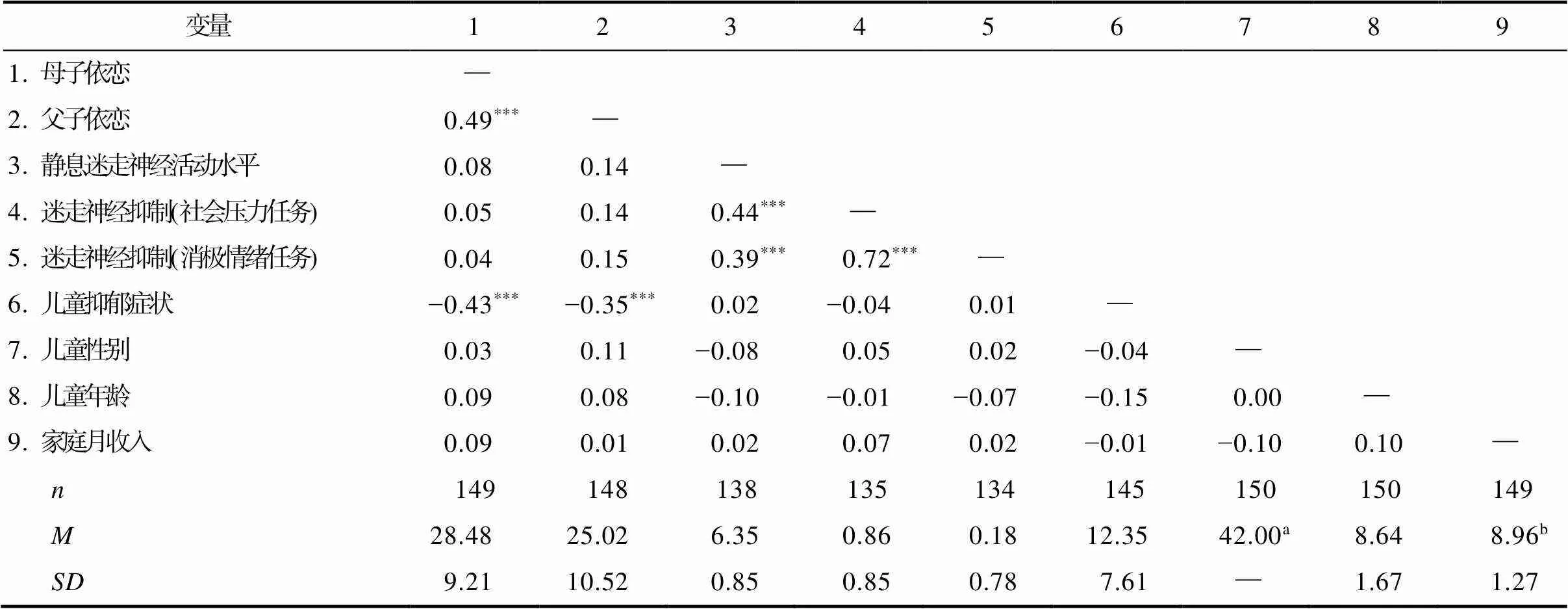
表1 本研究主要研究變量均值、標準差與相關關系表
注:靜息迷走神經活動水平為靜息狀態下兒童RSA的平均水平; 迷走神經抑制為相比于靜息狀態下, 兒童在任務中RSA水平的下降程度。兒童性別, 兒童年齡和家庭月收入為本研究的控制變量。
a女生所占的百分比。b家庭月收入從1 = “1000~1999元”到10 =“高于20000元”。
***< 0.001。
3.2 母子依戀、父子依戀水平差異
配對樣本檢驗表明, 兒童報告的母子依戀水平高于父子依戀水平(Δ= 3.55,(146) = 4.28,< 0.001, Cohen’s= 0.35)。
3.3 母子依戀、父子依戀對兒童抑郁癥狀的影響
由于控制變量中只有兒童年齡與兒童抑郁癥狀水平呈現負相關趨勢(= ?0.15,= 0.081), 因此后續分析僅將兒童年齡作為控制變量納入模型。將母子依戀與父子依戀同時納入路徑分析模型, 結果表明, 母子依戀負向影響兒童抑郁癥狀(= ?0.28,= 0.07, β = ?0.34,< 0.001), 父子依戀也存在負向影響兒童抑郁癥狀的趨勢(= ?0.12,= 0.07, β = ?0.17,= 0.058)。卡方差異檢驗結果表明, 將母子依戀、父子依戀影響兒童抑郁癥狀的路徑系數限制相等后, 模型擬合并沒有顯著變化(Δχ2= 1.66, Δ= 1,= 0.198), 表明母子依戀和父子依戀對兒童抑郁癥狀的影響程度沒有顯著差異, 均能負向影響兒童抑郁癥狀(= ?0.19,= 0.03,< 0.001)。
3.4 不同情境中迷走神經抑制程度差異
配對樣本檢驗表明, 相比于靜息狀態的迷走神經活動水平(= 6.35,= 0.85), 兒童在社會壓力任務(= 5.47,= 0.89)和消極情緒任務(= 6.18,= 0.90)中的迷走神經活動水平顯著降低, 即發生了迷走神經抑制(Δ社會壓力任務= 0.86,(134) = 11.76,< 0.001, Cohen’s= 1.00; Δ消極情緒任務= 0.16,(133) = 2.42,= 0.017, Cohen’s= 0.19)。
3.5 不同情境中兒童生物敏感性的調節作用
相關結果表明, 兒童在社會壓力任務和消極情緒任務中的兒童生物敏感性指標存在高相關(= 0.72,< 0.001)。采用路徑分析進行調節作用檢驗表明, 上述兩任務中兒童生物敏感性對母子、父子依戀影響兒童抑郁癥狀的調節作用模式也高度相似。具體來說, 在社會壓力任務中, 兒童生物敏感性調節母子依戀對兒童抑郁癥狀的影響(= ?0.31,= 0.09, β = ?0.29,< 0.001), 但不能調節父子依戀對兒童抑郁癥狀的影響(= 0.05,= 0.07, β = ?0.06,= 0.438)。進一步采用簡單斜率檢驗, 分析兒童生物敏感性調節母子依戀對兒童抑郁癥狀影響的模式。結果表明, 對于生物敏感性水平高(+1)的兒童, 其抑郁癥狀水平隨著母子依戀的升高而降低(= ?0.53,= 0.10, β = ?0.64,< 0.001), 而對于生物敏感性水平低(?1)的兒童, 其抑郁癥狀水平不受母子依戀變化的影響(= ?0.01,= 0.11, β = ?0.01,= 0.935)。顯著性區間檢驗表明, 社會壓力任務中兒童生物敏感性對母子依戀影響兒童抑郁癥狀的調節作用同時符合對環境的生物敏感性模型的3個判定標準(在母子依戀取值±2范圍內, 兒童生物敏感性與兒童抑郁癥狀呈現出不同方向的相關性, PoI = 0.42, PA = 0.53)。
同樣地, 在消極情緒任務中的兒童生物敏感性也能夠調節母子依戀對兒童抑郁癥狀的影響(= ?0.23,= 0.10, β = ?0.22,= 0.022), 但不能調節父子依戀對兒童抑郁癥狀的影響(= 0.06,= 0.08, β = 0.06,= 0.187)。進一步采用簡單斜率檢驗, 分析兒童生物敏感性調節母子依戀對兒童抑郁癥狀影響的模式。結果表明, 對于生物敏感性水平高(+1)的兒童, 其抑郁癥狀水平隨著母子依戀的升高而降低(= ?0.42,= 0.10, β = ?0.51,< 0.001), 而對于生物敏感性水平低(?1)的兒童, 其抑郁癥狀水平不受母子依戀水平變化的影響(= ?0.07,= 0.12, β = ?0.08,= 0.579)。顯著性區間檢驗表明, 消極情緒任務中兒童生物敏感性對母子依戀影響兒童抑郁癥狀的調節作用同時符合對環境的生物敏感性模型的3個判定標準(在母子依戀取值±2范圍內, 兒童生物敏感性與兒童抑郁癥狀呈現出不同方向的相關性, PoI = 0.44, PA = 0.53)。
上述結果表明, 兒童在社會壓力任務與消極情緒任務中的生物敏感性水平及其對母子、父子依戀影響兒童抑郁癥狀的調節作用模式高度相似, 表明兒童生物敏感性在本研究的兩個實驗任務中具有跨情境的一致性。后續分析將會對社會壓力任務與消極情緒任務中的兒童生物敏感性指標合成潛變量, 表示跨情境的兒童生物敏感性, 并進一步考察其對母子、父子依戀影響兒童抑郁癥狀的調節作用。
3.6 跨情境兒童生物敏感性的調節作用
跨情境兒童生物敏感性調節母子、父子關系對兒童抑郁癥狀影響的模型結果如圖1所示。為了使模型圖清晰易懂, 控制變量(兒童年齡)對兒童抑郁癥狀的影響以及變量間的相關未在圖中展示(如母子依戀與父子依戀之間的相關)。結果表明, 跨情境兒童生物敏感性調節母子依戀對兒童抑郁癥狀的影響(= ?0.27,= 0.07, β = ?0.34,< 0.001), 但不能調節父子依戀對兒童抑郁癥狀的影響(= 0.04,= 0.05, β = 0.06,= 0.441), 且兩調節效應強度差異顯著(Δχ2= 12.07, Δ= 1,< 0.001)。對跨情境兒童生物敏感性對母子依戀影響兒童抑郁癥狀的調節作用進行簡單斜率檢驗(如圖2), 結果表明, 跨情境兒童生物敏感性水平高(+1)的兒童, 其抑郁癥狀水平隨著母子依戀的升高而降低(= ?0.51,= 0.09, β = ?0.62,< 0.001), 而對于跨情境兒童生物敏感性水平低(?1)的兒童, 其抑郁癥狀水平不受母子依戀水平的變化的影響(= 0.03,= 0.12, β = 0.03,= 0.806)。
顯著性區間檢驗法的結果表明, 與生物敏感性低的兒童相比, 當母子依戀水平低(小于?0.43)時, 生物敏感性高的兒童報告了更高水平的抑郁癥狀; 當母子依戀水平高(大于+0.49)時, 生物敏感性高的兒童報告了更低水平的抑郁癥狀。此外, 通過計算可得, 該調節模型的PoI為0.45, PA為0.53。上述指標均表明本研究中跨情境兒童生物敏感性對母子依戀影響兒童抑郁癥狀的調節作用符合對環境的生物敏感性模型的標準。
4 討論
不良的親子依戀會損害兒童心理社會功能的發展, 增加兒童罹患抑郁等心理行為問題的風險(Ainsworth & Bowlby, 1991)。對環境的生物敏感性模型(Boyce & Ellis, 2005)指出, 親子依戀水平可能會對不同敏感性兒童造成不同的影響。考慮到我國傳統父母養育角色的差異(如“男主外, 女主內”)以及近年來教養模式的改變(如父親投入增加), 本研究探討了當代中國學齡兒童家庭中親子依戀對兒童抑郁癥狀的影響, 尤其關注上述影響是否會因兒童對環境的生物敏感性水平和父母角色不同而存在差異。結果表明, 雖然父子依戀水平整體上低于母子依戀水平, 但父子依戀與母子依戀對兒童抑郁癥狀的影響程度不存在顯著差異。此外, 與生理上對環境不敏感的兒童相比, 生物敏感性高(表現為迷走神經抑制程度高)的兒童更容易受到高母子依戀的積極影響或低母子依戀的消極影響; 而父子依戀對不同生物敏感性兒童抑郁癥狀的影響程度不存在顯著差異, 高水平的父子依戀均能降低兒童的抑郁癥狀水平。
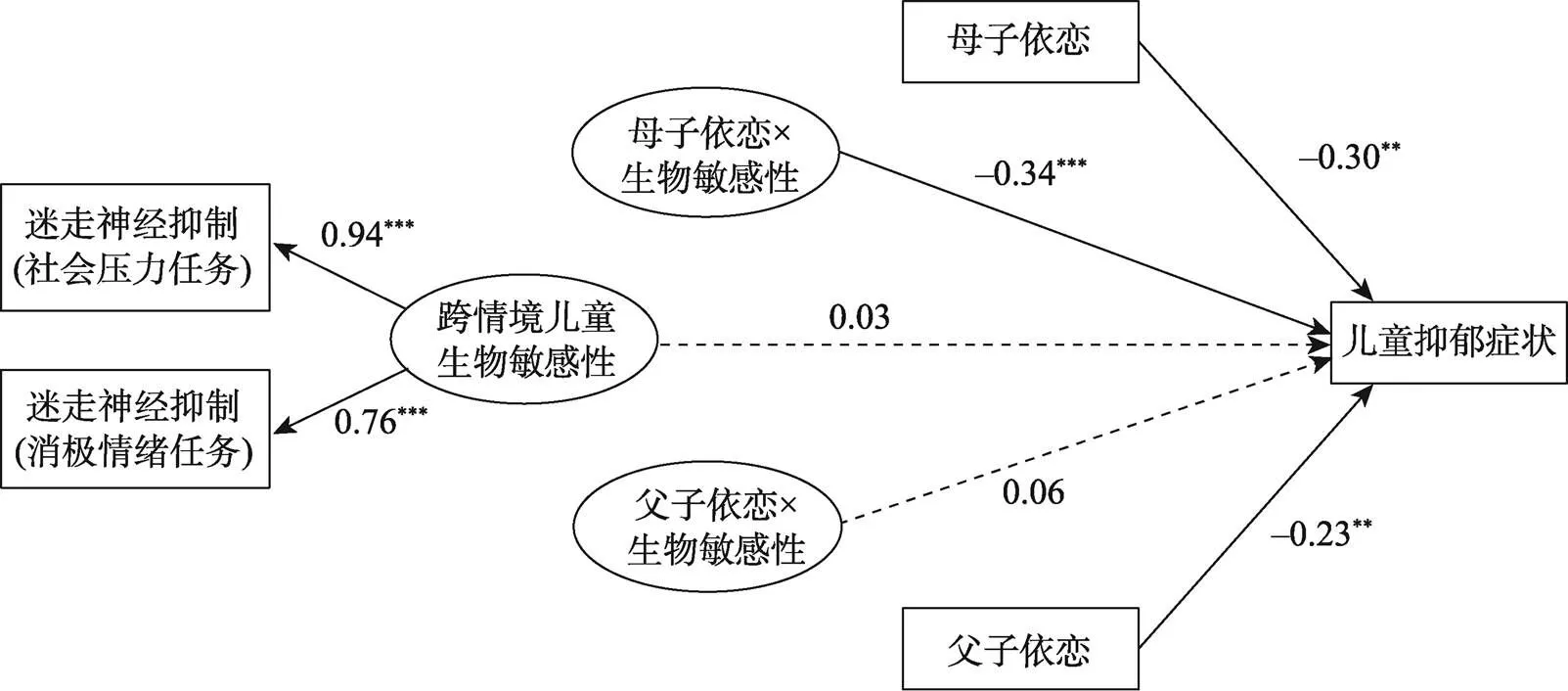
圖1 跨情境兒童生物敏感性調節母子、父子依戀對兒童抑郁癥狀影響的模型圖(圖中數字為標準化系數)
注:*< 0.05,**< 0.01,***< 0.001。
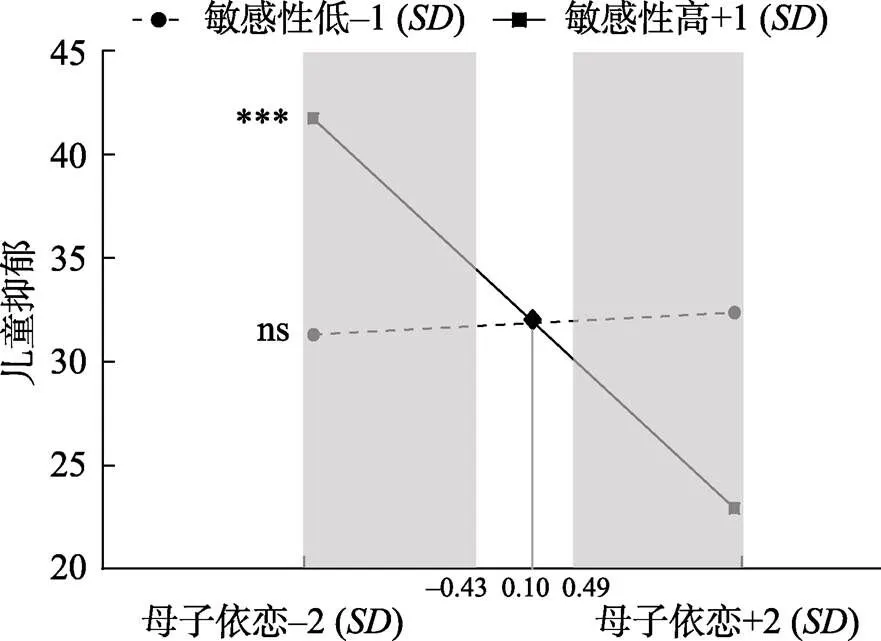
圖2 跨情境兒童生物敏感性對母子依戀影響兒童抑郁的調節作用示意圖
注:陰影部分代表顯著性區域(RoS)。*< 0.05,**< 0.01,***< 0.001。
4.1 母子依戀、父子依戀對兒童抑郁癥狀的影響
本研究發現, 母子依戀水平高于父子依戀水平。該研究結果支持了傳統家庭角色分工的觀點, 即母親在家庭中主要負責撫養后代、照顧子女, 而父親主要負責提供社會經濟資源, 相比于母親較少地參與到兒童的教養活動之中, 與兒童形成的依戀水平相對較低(侯芬等, 2018)。以往實證研究也發現, 相比于跟父親的互動, 兒童在與母親的互動的過程中會表現出更多的參與度和更積極的情緒(Kwon & Elicker, 2012)。同樣地, 在與兒童的互動中, 相比于父親, 母親也會有更高的參與程度和更多的積極教養行為(Kwon & Elicker, 2012)。
回歸分析和卡方差異比較的結果表明, 高水平的母子依戀與父子依戀均與兒童抑郁癥狀呈負相關, 且母子依戀與父子依戀影響兒童抑郁癥狀的強度不存在顯著差異。該結果為依戀理論中的多重依戀假說(Ainsworth, 1989)和親子依戀影響兒童發展的特異性假說(Howes & Ritchie, 1999)提供了實證支持, 即兒童與父親和母親的依戀水平都會對其心理健康產生重要影響。這說明作為兒童的重要撫養者之一, 父親用其獨特的方式與孩子進行互動, 對兒童的情緒和社會功能起著不可替代的作用(Amodia- Bidakowska et al., 2020)。近年來, 隨著我國社會文化的發展, 越來越多的父親參與到對孩子的直接養育之中(Li, 2020)。相比于本世紀初, 父親每天與孩子相處的時間、對撫養孩子的投入度均有所增加, 很可能對兒童的健康發展有著越來越大的影響(Li, 2020)。
4.2 兒童生物敏感性的調節作用
本研究結果表明, 迷走神經抑制程度可以作為兒童生物敏感性的指標調節母子依戀對兒童抑郁癥狀的影響。具體來說, 相比于對環境不敏感(迷走神經抑制程度低)的兒童, 高敏感(迷走神經抑制程度高)的兒童更容易受到母子依戀的積極或消極的影響。當母子依戀水平較高時, 高敏感的兒童會更多地從這種積極關系中獲益, 抑郁癥狀更少; 而當母子依戀水平較低時, 高敏感的兒童在消極關系中遭受更多損害, 抑郁癥狀更多。該結果符合對環境的生物敏感性模型(Boyce & Ellis, 2005)的觀點, 豐富了從家庭視角對該模型的實證探索(Cipriano et al., 2011; Han et al., 2020; Tabachnick et al., 2021)。進一步地, 在社會壓力任務和消極情緒任務兩個性質不同的情境中所測量的迷走神經抑制對母子依戀影響兒童抑郁癥狀的作用存在高度的相似性, 表明迷走神經抑制作為生物敏感性指標的作用很可能是跨情境一致的。這為回答生物敏感性作用是“情境特異”還是“跨情境一致”之爭提供了實證證據。
但與假設不符的是, 不同情境中所測量的兒童迷走神經抑制程度均不能調節父子依戀對兒童抑郁癥狀的影響, 即父親對兒童抑郁的作用似乎并不受兒童對環境的生物敏感性的影響。雖然以往研究沒有直接檢驗兒童生物敏感性對母子、父子互動影響兒童發展的差異作用, 但有其他研究表明, 相比于父子關系, 母子關系對兒童發展的作用程度更易受到兒童自身特質的影響。例如, 母親管教對兒童學業成績和社交能力的影響會因兒童互依型自我構念水平的不同而存在差異, 但上述調節效應并不存在于父親管教對兒童學業和社交能力的影響中(Lan et al., 2019)。此外, 消極的母子互動關系(如母子沖突、母親拒絕)對孩子抑郁相關癥狀的影響程度會受到孩子的年齡、是否為獨生子女等自身特質的調節(肖雪等, 2017; Ramírez-Uclés et al., 2018), 但上述研究并沒有發現消極的父子互動關系對孩子抑郁癥狀的影響會因孩子自身特質不同而發生變化。值得指出的是, 在本研究中, 父子依戀對兒童抑郁癥狀的直接效應顯著。因此, 從另一個角度看, 不顯著的調節作用意味著在本研究中相比于母子依戀, 父子依戀對不同敏感性兒童抑郁癥狀的影響可能更為一致, 即不論兒童是否具備敏感的生理特質, 與父親形成安全穩定的依戀均能夠一定程度降低其抑郁癥狀水平。未來研究可以采用更加廣泛和有代表性的樣本, 進一步檢驗本研究結果的可重復性。
4.3 研究意義
在理論層面, 本研究檢驗了不同情境中兒童生物敏感性對親子依戀影響兒童抑郁癥狀的作用及其父母角色差異, 推動了該領域對兒童生物敏感性測量方式的完善與敏感性理論的構建。以往考慮迷走神經抑制作為生物敏感性指標的文章主要通過單一的實驗范式對其進行測量, 其結果存在較大的偶然性(Tabachnick et al., 2021)。僅有的幾篇采用不同情境測量迷走神經抑制的文章大多未經詳細檢驗, 直接將不同任務中的迷走神經抑制合成總分, 這容易忽視不同情境中迷走神經抑制的獨特作用(Cipriano et al., 2011; Obradovi? et al., 2010)。本研究考慮到上述研究的局限性, 首先對比不同情境中迷走神經抑制程度的相似性; 再比較不同情境中迷走神經抑制程度對母子依戀影響兒童抑郁是否均存在調節作用; 如果調節作用均存在, 進而比較其模式是否一致(如是否都符合敏感性模型的條件)。基于以上標準綜合判斷迷走神經抑制是否可以作為兒童生物敏感性的指標, 以及其作用是跨情境一致的還是跨情境特異的, 創新性地提出了檢驗不同情境中兒童生物敏感性作用的新標準。進一步地, 基于該標準, 本研究發現兒童生物敏感性對親子依戀影響兒童抑郁的作用很可能是跨情境一致的且存在父母差異, 提出對環境的生物敏感性模型的構建需要進一步考慮敏感性指標測量的情境以及與兒童互動的角色(如母親和父親)。
在實踐層面, 本研究表明家庭中高水平的父子依戀與母子依戀均能降低兒童抑郁癥狀的水平, 且父子依戀對兒童抑郁癥狀的作用并未受到兒童生物敏感性的影響。再考慮到目前父子依戀程度仍低于母子依戀程度, 處于相對較低的水平, 心理學工作者有必要呼吁全社會加強對父親教養投入的重視, 提升父子依戀水平, 進一步降低兒童產生心理健康問題的風險。此外, 兒童敏感性對母子依戀影響兒童抑郁癥狀的調節作用表明母親對兒童發展的影響可能更為多樣化, 即母親可能更需要根據兒童自身的特點來調整自己對待孩子的方式, 以最大程度地促進孩子的健康發展。
4.4 研究不足與未來展望
本研究在理論和實踐層面均有著較大貢獻, 但仍存在以下幾點不足。第一, 本研究的兒童發展結果僅考慮兒童抑郁癥狀, 沒有同時考慮兒童不同方面的發展指標。有研究表明, 母親與父親對兒童不同方面的發展結果有著不同的影響。例如, 母子依戀更可能影響兒童的抑郁等情緒問題, 而父子依戀與兒童的攻擊性等外化問題關聯更為密切(Stover et al., 2016; 肖雪等, 2017)。未來研究應綜合考慮兒童學業、社會、心理等各方面的發展指標, 系統探究母子、父子依戀對兒童各方面發展的影響的異同。
第二, 本研究采用問卷報告法測量兒童的親子依戀和抑郁癥狀水平, 可能存在回憶偏差等問卷報告的局限性。有研究基于真實情境中的親子互動任務, 通過觀察編碼等形式測量兒童與家長的互動特點及兒童的消極情緒反應, 探究母子、父子互動特點對兒童發展的影響(Lunkenheimer et al., 2020)。未來研究可基于父?母?子三人真實互動, 結合多種技術手段, 同時觀察到母子、父子亞系統之間的依戀程度和教養方式, 以及父母如何協同教養, 從家庭系統的視角更為全面地探討父親、母親對兒童發展的即時和長期影響及其差異。
第三, 本研究雖然基于兩個性質不同的壓力情境測量兒童的生物敏感性, 但并沒有考慮兒童在中性或積極情境中迷走神經反應能否作為生物敏感性指標影響外界環境對兒童發展的作用。多重迷走神經理論(the polyvagal theory; Porges, 2007)指出, 個體在面對壓力事件時會發生迷走神經抑制, 但個體在面對壓力程度較低的社會參與情境中迷走神經活動不僅不會抑制反而會增強, 這會幫助個體更好地投入到與他人的社會互動中。在放松的社會參與情境中, 若發生較高程度的迷走神經抑制, 反而會意味著個體存在較大程度的壓力, 沒有很好地投入到互動中。在這種情境下, 迷走神經抑制水平很可能并不是個體敏感性的指標, 迷走神經抑制對親子依戀影響兒童發展的作用也不一定會符合對環境的生物敏感性模型的標準。與之相對, 積極互動情境中過高的迷走神經抑制可能反映出個體的生物脆弱性(biological vulnerability), 使得其與親子依戀共同影響兒童發展的模式呈現出“素質?壓力”模型的形式。因此, 未來研究應系統測量兒童在積極、中性和消極情境中的迷走神經活動, 檢驗其是否能作為生物敏感性的指標, 以更為全面地解答生物敏感性作用的“跨情境一致”和“情境特異”之爭。
第四, 本研究參考以往測量不同情境下迷走神經抑制的研究(Cipriano et al., 2011; Obradovi? et al., 2010), 未在不同被試間將社會壓力和消極情緒這兩個挑戰任務的順序進行平衡。未來研究需要考慮不同任務順序是否會對迷走神經抑制程度及其作用產生影響, 更加嚴謹地探討迷走神經抑制在不同情境中的作用特點。
5 結論
本研究探討了母子、父子依戀對兒童抑郁癥狀的影響及差異, 并進一步檢驗不同情境中兒童生物敏感性(即迷走神經抑制)在其中的調節作用。研究發現, 母子、父子依戀整體上對兒童抑郁癥狀的影響程度相當, 但存在不同的作用特點:母子依戀對高敏感兒童的影響程度更大, 而父子依戀對不同敏感性的兒童均存在影響。本研究從跨情境和父母差異視角豐富了對環境的生物敏感性模型, 并揭示了母子、父子依戀對兒童抑郁癥狀發展的獨特且重要的作用。
Ainsworth, M. D. S. (1989). Attachments beyond infancy.,(4), 709?716.
Ainsworth, M. S., & Bowlby, J. (1991). An ethological approach to personality development.,(4), 333?341.
Amodia-Bidakowska, A., Laverty, C., & Ramchandani, P. G. (2020). Father-child play: A systematic review of its frequency, characteristics and potential impact on children’s development.,, 100924.
Armsden, G. C., & Greenberg, M. T. (1987). The inventory of parent and peer attachment: Individual differences and their relationship to psychological well-being in adolescence.,(5), 427?454.
Belsky, J. (1997). Attachment, mating, and parenting: An evolutionary interpretation.,(4), 361?381.
Boyce, W. T., & Ellis, B. J. (2005). Biological sensitivity to context: I. An evolutionary?developmental theory of the origins and functions of stress reactivity.,(2), 271?301.
Bronfenbrenner, U., & Morris, P. A. (1998). The ecology of developmental processes. In W. Damon (Series Ed.) & R. M. Lerner (Vol. Ed.),(5th ed., pp. 993?1028). Wiley.
Chen, X., Chen, X., Zhao, S., Way, N., Yoshikawa, H., Zhang, G., ... Li, D. (2021). Autonomy-and connectedness-oriented behaviors of toddlers and mothers at different historical times in urban China.,(8), 1254?1260.
Chen, Z. Y., Yang, X. D., & Li, X. Y. (2009). Psychometric features of CES-D in Chinese adolescents.,(4), 443?445.
[陳祉妍, 楊小冬, 李新影. (2009). 流調中心抑郁量表在我國青少年中的試用.,(4), 443?445.]
Cipriano, E. A., Skowron, E. A., & Gatzke-Kopp, L. M. (2011). Preschool children’s cardiac reactivity moderates relations between exposure to family violence and emotional adjustment.,(3), 205?215.
Copeland, W. E., Alaie, I., Jonsson, U., & Shanahan, L. (2021). Associations of childhood and adolescent depression with adult psychiatric and functional outcomes.,(5), 604?611.
Cox, M. J., & Paley, B. (1997). Families as systems.(1), 243?267.
Davis, E. L., Brooker, R. J., & Kahle, S. (2020). Considering context in the developmental psychobiology of self-regulation.,(4), 423?435.
Eisenberg, N., Cumberland, A., & Spinrad, T. L. (1998). Parental socialization of emotion.,(4), 241? 273.
Fortunato, C. K., Gatzke-Kopp, L. M., & Ram, N. (2013). Associations between respiratory sinus arrhythmia reactivity and internalizing and externalizing symptoms are emotion specific.,(2), 238?251.
Fracasso, M. P., Porges, S. W., Lamb, M. E., & Rosenberg, A. A. (1994). Cardiac activity in infancy: Reliability and stabilityof individual differences.,(3), 277?284.
García, J. I. R., Manongdo, J. A., & Ozechowski, T. J. (2014). Depression symptoms among Mexican American youth: Paternal parenting in the context of maternal parenting, economic stress, and youth gender.,(1), 27?36.
Han, Z. R., Zhang, X., Davis, M., & Suveg, C. (2020). The role of children’s neurophysiological functioning in the links between emotion-parenting behaviors and child anxiety symptoms: A biological sensitivity to context framework.,(2), 618?635.
Hou, F., Wu, X. C., Zou, S. Q., Liu, Y, & Huang, B. B. (2018). The association between parental involvement and adolescent’s prosocial behavior: The mediating role of parent-child attachment.,(4), 417?425.
[侯芬, 伍新春, 鄒盛奇, 劉暢, 黃彬彬. (2018). 父母教養投入對青少年親社會行為的影響: 親子依戀的中介作用.,(4), 417?425.]
Howes, C., & Ritchie, S. (1999). Attachment organizations in children with difficult life circumstances.,(2), 251?268.
Jin, C. C., Zou, H., Zeng, R., & Dou, D. H. (2010). The trait of attachment and the effect of attachment on social adjustment of middle school students: Parents intimacy as a moderator.,(6), 577?583.
[金燦燦, 鄒泓, 曾榮, 竇東徽. (2010). 中學生親子依戀的特點及其對社會適應的影響: 父母親密的調節作用.,(6), 577?583.]
Kwon, K. A., & Elicker, J. G. (2012). The role of mothers' and fathers' parental control and coparenting in toddlers’ compliance.,(5), 748?765.
Lan, X., Scrimin, S., & Moscardino, U. (2019). Perceived parentalguan and school adjustment among Chinese early adolescents:The moderating role of interdependent self-construal.,, 18?27.
Li, X. (2020). Fathers’ involvement in Chinese societies: Increasing presence, uneven progress.,(3), 150?156.
Lunkenheimer, E., Hamby, C. M., Lobo, F. M., Cole, P. M., & Olson, S. L. (2020). The role of dynamic, dyadic parent- child processes in parental socialization of emotion.,(3), 566?577.
McAdams, D. P., & Bauer, J. J. (2004). Gratitude in modern life: Its manifestations and development. In R. A. Emmons & M. E. McCullough (Eds.),(pp. 81?99). Oxford University Press.
Mer?on-Vargas, E. A., Lima, R. F. F., Rosa, E. M., & Tudge, J. (2020). Processing proximal processes: What Bronfenbrenner meant, what he didn't mean, and what he should have meant.,(3), 321?334.
Mezulis, A. H., Crystal, S. I., Ahles, J. J., & Crowell, S. E. (2015). Examining biological vulnerability in environmental context: Parenting moderates effects of low resting respiratory sinus arrhythmia on adolescent depressive symptoms.,(8), 974?983.
Murphy, S. E., Boyd‐Soisson, E., Jacobvitz, D. B., & Hazen, N. L. (2017). Dyadic and triadic family interactions as simultaneous predictors of children's externalizing behaviors.,(2), 346?359.
Obradovi?, J., Bush, N. R., Stamperdahl, J., Adler, N. E., & Boyce, W. T. (2010). Biological sensitivity to context: The interactive effects of stress reactivity and family adversity on socioemotional behavior and school readiness.,(1), 270?289.
Porges, S. W. (2007). The polyvagal perspective.,(2), 116?143.
Qui?ones-Camacho, L. E., & Davis, E. L. (2019). Parasympathetic regulation in cognitive and emotional challenge contexts differentially predicts specific aspects of children’s emotionalfunctioning.,(2), 275?289.
Radloff, L. S. (1977). The CES-D scale: A self-report depression scale for research in the general population.,(3), 385?401.
Ramírez-Uclés, I., González-Calderón, M. J., del Barrio-Gándara, V., & Carrasco, M. á. (2018). Perceived parental acceptance- rejection and children’s psychological adjustment: The moderating effects of sex and age.,(4), 1336?1348.
Roisman, G., Newman, D., Fraley, R., Haltigan, J., Groh, A., & Haydon, K. (2012). Distinguishing differential susceptibility from diathesis?stress: Recommendations for evaluating interaction effects.(2), 389?409.
Schlomer, G. L., Bauman, S., & Card, N. A. (2010). Best practices for missing data management in counseling psychology.(1), 1?10.
Shen, Y., Zheng, J., Shi, Z. M., Sun, M., Luo, H. Z., Cui, Y., ... Ji, H. X. (2021). Investigation of anxiety and depression in children and adolescents in Chongqing.,(6), 991?993.
[沈彥, 鄭嬌, 史戰明, 孫蒙, 羅華中, 崔宇, ... 吉航西. (2021). 重慶市兒童青少年焦慮抑郁發生情況調查.,(6), 991?993.]
Shuang, M., Yiqing, W., Ling, J., Guanzhen, O., Jing, G., Zhiyong, Q., & Xiaohua, W. (2022). Relationship between parent-child attachment and depression among migrant children and left-behind children in China.,, 1?8.
Spruit, A., Goos, L., Weenink, N., Rodenburg, R., Niemeyer, H., Stams, G. J., & Colonnesi, C. (2020). The relation between attachment and depression in children and adolescents: A multilevel meta-analysis.,(1), 54?69.
Stover, C. S., Zhou, Y., Kiselica, A., Leve, L. D., Neiderhiser, J. M., Shaw, D. S., Natsuaki, M. N., Scaramella, L. V., & Reiss, D. (2016). Marital hostility, hostile parenting, and child aggression: Associations from toddlerhood to school age.,(3), 235?242.
Suess, G. J., Grossmann, K. E., & Sroufe, L. A. (1992). Effects of infant attachment to mother and father on quality of adaptation in preschool: From dyadic to individual organisation of self.,(1), 43?65.
Tabachnick, A. R., Moore, C., Raby, K. L., Goldstein, A., Zajac, L., & Dozier, M. (2021). Respiratory sinus arrhythmia as a moderator of early maltreatment effects on later externalizing problems.,(3), 821?831.
Teasdale, J. D. (1983). Negative thinking in depression: Cause, effect, or reciprocal relationship?,(1), 3?25.
Wang, X. D., Wang, X. L., & Ma, H. (1999).. Beijing, China: Chinese Mental Health Journal Press.
[汪向東, 王希林, 馬弘. (1999).. 北京: 中國心理衛生雜志社.]
Wang, X. L., Chen, L. H., Bu, Y., & Lin, D. H. (2018). The effects of paternal-and maternal-attachment on children’ s cortisol reactivity to stress.,(1), 10?20.
[王曉蕾, 陳麗華, 卜鈺, 林丹華. (2018). 父子依戀、母子依戀與兒童應激下皮質醇反應的關系.,(1), 10?20.]
Xiao, X., Liu L. S., Xu, L. Y., & Li, Y. F. (2017). Marital conflict, parent-child relationship and adolescents' depression in Chinese only and non-only child family.,(4), 468?476.
[肖雪, 劉麗莎, 徐良苑, 李燕芳. (2017). 父母沖突, 親子關系與青少年抑郁的關系: 獨生與非獨生的調節作用.,(4), 468?476.]
Yao, W. Y., Zhang, W., Liu, Y., Zhang, S. M., Ding, X. C., & Xu, G. M. (2021). Developmental trajectories of depression and academic achievement in children: Based on parallel latent growth modeling.,(2), 223?229.
[姚文玉, 張雯, 劉影, 張思敏, 丁雪辰, 徐剛敏. (2021). 兒童抑郁水平與學業成績的發展軌跡: 基于平行潛變量增長模型.,(2), 223?229.]
Zeytinoglu, S., Calkins, S. D., & Leerkes, E. M. (2020). Autonomic nervous system functioning in early childhood: Responses to cognitive and negatively valenced emotional challenges.,(5), 657?673.
Zhao, F. Q., Cheng, B. B., Li, Y. X., Liu, Z. B., Liu, B., & Zhang, B. S. (2022). Father coparenting and adolescent depression in nuclear families: The mediating roles of father-child attachment and mother-child attachment.,(1), 109?117.
[趙鳳青, 程貝貝, 李奕萱, 劉志斌, 劉兵, 張寶山. (2022). 核心家庭父親協同教養對青少年抑郁的影響: 父子依戀和母子依戀的中介作用.,(1), 109?117.]
Parent-child attachment and children’s depressive symptoms: The role of children’s biological sensitivity and parental gender differences
XU Jianjie, ZHANG Yiyi, LAM Tak Kwan, CHE Liying, SONG Manman, HAN Zhuo
(Faculty of Psychology, Beijing Normal University, Beijing 100875, China)
Secure and stable parent-child attachment is beneficial for children’s mental health, whereas maladaptive parent-child attachment may lead to children’s maladjustment such as depressive symptoms. Due to rapid sociocultural development in China, traditional patriarchal parenting patterns with fathers as breadwinners and mothers as homemakers have gradually diminished. Instead, Chinese fathers today spend an increasing amount of involvement in their children’s development. However, little to no research has examined the unique effects of father-child and mother-child attachment on children’s developmental outcomes in contemporary China. More importantly, the Biological Sensitivity to Context Theory (BSCT) suggests that the association between parent- child attachment and child developmental outcomes may vary among children with different levels of biological sensitivity. According to this theory, children who are biologically sensitive are more susceptible to adaptive or maladaptive parent-child relationships. Taken together, the present study aimed to examine how children’s depressive symptoms were affected by mother-child and father-child attachments and whether this effect was moderated by the children’s biological sensitivity. The potential father vs. mother difference was also examined.
150 school-aged children (63 girls and 87 boys,age= 8.64 years) participated in the current study. Children reported their depressive symptoms as well as their perceived parent-child attachment with mothers and fathers, respectively. Children’s biological sensitivity (i.e., vagal suppression) was assessed by the decrease of respiratory sinus arrhythmia (RSA) between the resting phase and the task phases (i.e., a social stress task and a negative emotion provoking task) through the Biopac MP150 systems. Descriptive statistics and bivariate correlations were analyzed via SPSS 25.0, and moderation models were conducted via Mplus 8.3.
Our results are highlighted by five major points: (1) The level of mother-child attachment was higher than that of father-child attachment. (2) Both mother-child attachment and father-child attachment were uniquely and negatively associated with children's depressive symptoms, and the strengths of the aforementioned paths were equivalent. (3) Children's biological sensitivity (vagal suppression) measured in the social stress task and the negative emotion provoking task had similar moderating effects on the relationship between parent-child attachment and children’s depressive symptoms, indicating the cross-context consistency of the roles of biological sensitivity. (4) Children’s cross-context biological sensitivity moderated the relationship between mother-child attachment and children’s depressive symptoms. Specifically, highly sensitive children (vs. non-sensitive children) were more likely to benefit from secure mother-child attachment but were also more likely to be harmed by insecure mother-child attachment. (5) Children's cross-context biological sensitivity did not moderate the relationship between father-child attachment and children’s depressive symptoms, such that higher father-child attachment was consistently associated with lower children’s depressive symptoms, regardless of children’s levels of biological sensitivity.
Based on attachment theory and the BSCT, the present study indicates that children’s attachment with their mothers or fathers are uniquely associated with children's depressive symptoms, and that mother-child attachment jointly interacted with children’s biological sensitivity to influence children’s depressive symptoms. As a theoretical application, our study innovatively suggests that future studies should consider the context in which an indicator of biological sensitivity is assessed as well as parental roles (father vs. mother) when testing the BSCT in family studies. As a practical application, our findings indicate the potential different roles of father-child attachment and mother-child attachment in protecting children from suffering depressive symptoms, providing empirical evidence to support the development of family-based prevention and intervention projects aimed at alleviating children’s psychopathological problems.
parent-child attachment, children’s depressive symptoms, biological sensitivity to context, parental gender
2021-09-26
*北京市社會科學基金重點項目(21DTR030)資助。
韓卓, E-mail: rachhan@bnu.edu.cn
B844
猜你喜歡
福建中學數學(2023年5期)2024-01-25 17:41:36
中學生數理化·中考版(2022年10期)2022-11-10 09:37:46
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
護士進修雜志(2017年3期)2017-02-14 07:19:35
小學生作文(中高年級適用)(2016年3期)2016-11-11 06:30:23
雜文選刊(2016年7期)2016-08-02 08:39:56
小天使·一年級語數英綜合(2016年6期)2016-05-14 12:21:05
