藥用植物黃芩的生物學研究進展及展望
2023-03-07 12:54:40鄭敏敏柳潔趙清
生物技術通報 2023年2期
鄭敏敏 柳潔 趙清
(1.上海辰山植物園 中國科學院分子植物科學卓越創新中心辰山科學研究中心 上海市資源植物功能基因組學重點實驗室,上海 201602;2.中國科學院大學,北京 100049)
黃芩(Scutellaria baicalensisGeorgi)為唇形科黃芩屬多年生草本植物,具披針或線性披針形葉,根莖肉質,花紫色或藍色(圖1),原產于中國、俄羅斯東部、蒙古、朝鮮、日本等地[1-2]。植物黃芩的干燥根即是傳統中藥里的“黃芩”,在中國具有悠久的藥用歷史,最早記載“黃芩”藥用的書籍是漢代的《神農本草經》,主治肺和肝臟的疾病[3]。東漢時期,張仲景的《傷寒雜病論》中也記載了含有“黃芩”的多個復方湯劑(表1),用于治療風寒、頭疼、發熱、腹瀉等疾病[4]。明代的醫學著作《本草綱目》中也記載“黃芩”具有清熱燥濕、瀉火解毒、消炎、止血安胎以及健脾胃等作用,可用于治療腹瀉、腹痛、出血、炎癥和呼吸道感染等疾病[5-6]。

圖1 黃芩植株、黃芩的根和黃芩藥材Fig. 1 S. baicalensis plant, S. baicalensis root and S. baicalensis as medicinal material
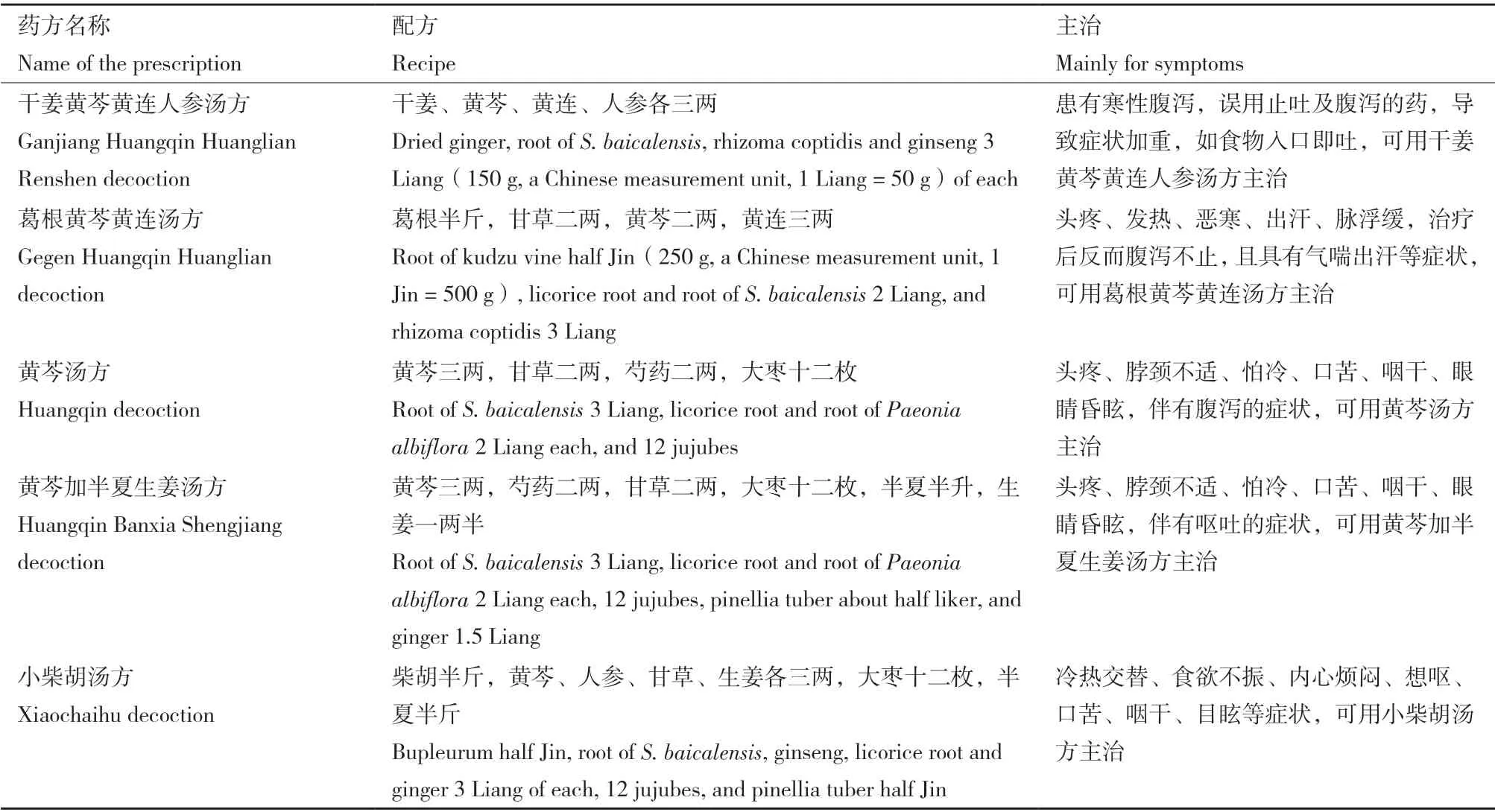
表1 《傷寒雜病論》里含有黃芩的復方湯劑Table 1 Compound decoctions in “Treatise on Febrile and Miscellaneous Diseases” containing S. baicalensis
1 黃芩的化學成分
隨著提取和分離技術的發展,迄今為止,已從黃芩植物中分離出132 種化合物,這些化合物大多從根中分離得到。根據結構劃分,化合物主要類型包括類黃酮化合物、苯乙醇苷、固醇類、萜類、酰胺以及酚類化合物,其中黃酮及其苷類化合物占絕大部分。132 種化合物中游離的類黃酮化合物有56種,包括42 種黃酮、3 種黃酮醇、9 種黃烷酮、1種查耳酮和1 種雙黃酮,其中黃芩素、漢黃芩素的含量最高(圖2-a,2-c);黃酮苷類有44 種,其中黃芩苷的含量最高,其次為漢黃芩苷(圖2-b,2-d);此外,還含有8 種苯乙醇苷、3 種固醇、1 種二萜、5 種酰胺、9 種酚類化合物以及6 種多糖[7-8]。
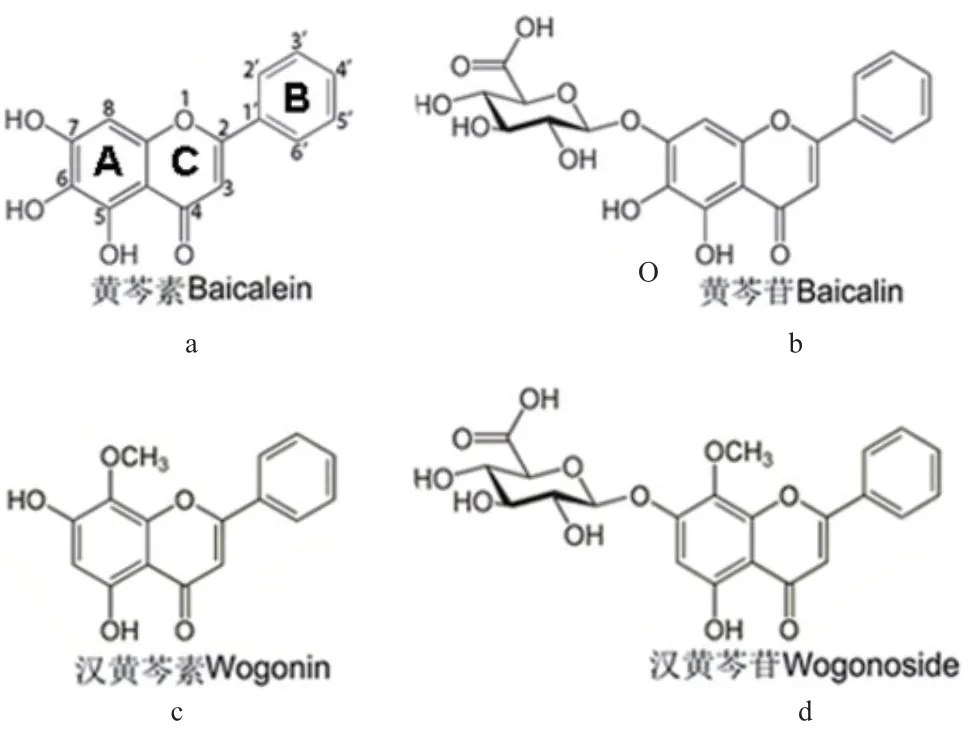
圖2 黃芩中主要活性黃酮的結構Fig. 2 Structures of main active flavones in S. baicalensis
黃芩中的黃酮化合物結構特殊,除常見的C-5和C-7 位被取代基取代以外,C-6 和C-8 位上也會被羥基取代,或C-8 位被甲氧基取代,這在其他植物中是很少見的。此外,黃芩植物中黃酮苷化合物的糖苷類型可分為O-葡萄糖苷、O-葡萄糖醛酸苷以及C-葡萄糖苷,大部分O-葡萄糖苷在7-OH 或2′-OH 上糖基化,作為糖基,葡萄糖醛酸苷不像葡萄糖苷普遍,但在黃芩中,至少存在17 種葡萄糖醛酸苷,且普遍在7-OH 上糖基化,少數在8-OH 和2′-OH 上[8]。
近幾十年來,大量臨床和藥理研究表明,黃酮化合物是黃芩的主要生物活性物質,尤其是根部的
2 黃芩中的藥用活性物質及藥理研究
黃芩素、漢黃芩素及其苷類,具有抗腫瘤、抗病毒、抗炎、抗氧化、護肝及神經保護等藥理活性[6]。
2.1 抗腫瘤活性
黃芩素、漢黃芩素及其苷類具有良好的抗腫瘤活性,對乳腺癌、結直腸癌、肝癌、肺癌等多種癌癥都具有抑制效果,其主要機制是通過阻斷癌細胞的增殖,抑制癌細胞轉移、激活癌細胞衰老凋亡或者誘導機體自噬[9-12]。腫瘤的發生、擴散和對治療產生的抗性與抗凋亡蛋白 Bcl-2、Bcl-x(L)、Bcl-w和 Mcl-1 的過度表達密切相關。Polier 等[13]將可抑制Bcl-2、Bcl-x(L)和 Bcl-w 的抑制劑ABT-263 與漢黃芩素、黃芩素、白楊素等活性黃酮共同處理 T細胞白血病異種移植小鼠,發現這些活性黃酮可以通過下調 Mcl-1 來增強ABT-263 誘導細胞凋亡表達,并且漢黃芩素對正常增殖的T 細胞和血小板沒有毒性。黃芩素能通過劑量依賴性調節甲狀腺癌細胞的NF-kB 信號改變Bax/Bcl-2 比率,介導細胞凋亡蛋白濃度增加導致癌細胞發生自噬,還可以降低有絲分裂蛋白Cyclin B1 的濃度抑制甲狀腺癌細胞的有絲分裂細胞周期,從而抑制癌細胞增殖[14]。漢黃芩素主要通過信號通路如絲氨酸-蘇氨酸激酶 Akt 和AMP活化蛋白激酶(AMPK)通路、p53 依賴性/非依賴性細胞凋亡、端粒酶活性的抑制、轉錄因子(NF-κB和AP)的抑制、活性氧(ROS)以及ER 應激介導的細胞凋亡和自噬來誘導癌細胞的凋亡和抑制癌細胞增殖[15-16]。此外,黃芩素和漢黃芩素與成熟的化療藥物如水飛薊素、順鉑、10-羥基喜樹堿(HCPT)、多西他賽(多烯紫杉醇)聯合使用,可以明顯提高抗癌效果,還能降低癌細胞的耐藥性以及對正常細胞的毒性[17-22]。黃芩素還可以與靶向治療乳腺癌的溶瘤麻疹病毒(oncolytic measles virus, MV)聯用,先利用MV 感染乳腺癌細胞,再聯合黃芩素治療,可以提高誘導癌細胞的凋亡的比率[23]。
2.2 抗病毒活性
黃芩素、漢黃芩素及其苷類能夠有效地抑制病毒的復制,具有抗病毒活性。新型冠狀病毒(SARSCoV-2)于2019年末開始在全球肆虐,在尋找有效藥物治療新型冠狀病毒病的過程中,發現中醫藥中的“三藥三方”,即金花清感顆粒、連花清瘟顆粒和膠囊、血必凈注射液、清肺排毒湯、化濕敗毒方、宣肺敗毒方在此次疫情中控制COVID-19 的臨床療效顯著[24-25]。吉米麗汗·司馬依等[26]研究表明,金花清感顆粒的山柰酚、黃芩素和千層紙黃素A 等活性成分可能通過與ACE2 結合作用于PTGS2、BCL2和CASP3 等靶點調節多條信號通路,從而發揮對COVID-19 的治療作用。Xia 等[27]分析了連花清瘟膠囊中10 種活性成分,構建了由65 個靶基因組成的與COVID -19 相關的靶基因集,篩選出了9 個中心基因,將參與肺損傷、肺纖維化和病毒感染的最重要基因Akt1與連花清瘟膠囊6 種活性成分(β-胡蘿卜素、山柰酚、木犀草素、柚皮素、槲皮素和漢黃芩素)進行分子對接,發現6 種活性成分均可進入Akt1 的活性口袋,連花清瘟膠囊主要通過調節免疫反應、細胞凋亡和抵御病毒感染等機制發揮作用。孫遜等[28]和劉暢等[29]基于網絡藥理學和分子對接法,對化濕敗毒方的主要活性成分進行篩選,構建成分-疾病靶點網絡,并與新型冠狀病毒的3CL 水解酶(3CLpro, Mpro)、血管緊張素轉化酶Ⅱ(ACE2)進行分子對接,結果表明與Mpro 結合最好的是黃芩素。Zhao 等[30]研究表明清肺排毒湯中4 種活性物質(黃芩苷、甘草酸、橙皮苷和金絲桃苷)和細胞的7 個靶點(AKT1、TNF-α、IL6、PTGS2、HMOX1、IL10 和 TP53)是與清肺排毒湯療效相關的關鍵分子。此外,對于其他一些病毒如艾滋病病毒(HIV)[31]、呼吸道合胞病毒(RSV)[32]、能引起新生兒畸形的寨卡病毒(ZIKV)[33]、登革熱病毒[34-35]、導致失能性關節炎的基孔肯雅病毒(CHIKV)[36]等病毒,黃芩素和黃芩苷也表現出了良好的抗病毒活性。
2.3 抗炎
炎癥可能引起細胞因子風暴,導致器官的損傷和衰竭[37]。研究表明,黃芩主要活性成分黃芩素、漢黃芩素及其苷類具有抗氧化和抗炎的功效,已經在多種疾病中得到驗證,包括肝炎[38]、心血管疾病[39]、肺部疾病[40]、過敏性氣道炎[41]、神經炎癥及神經退行性疾病[42-43]以及腸道疾病[44]。黃芩素、漢黃芩素及其苷類能夠直接作用于淋巴細胞、巨噬細胞、肥大細胞、樹突狀細胞、單核細胞、中性粒細胞等免疫細胞,抑制其產生炎癥細胞因子 IL-1β、IL-6、IL-8 和 TNF-α,以及其他炎癥介質,如一氧化氮(NO)、前列腺素(PGs)、白三烯(LTs)和活性氧(ROS)作用于免疫細胞,抑制其產生炎癥因子及其他炎癥介質。分子機制主要是通過toll 樣受體的下調、Nrf2 和 PPAR 信號通路的激活以及核硫氧還蛋白系統和炎癥相關通路(如 MAPK、Akt、NFκB 和 JAK-STAT)的抑制[37,45]。Huang 等[38]研究發現,黃芩苷能通過上調 TUG1 的表達,從而抑制p38MAPK 和 JNK 通路,減輕由脂多糖(LPS)刺激的正常肝臟L-02 和THLE2 細胞凋亡和炎癥反應。
3 黃芩的生物技術研究
黃芩中的黃酮化合物藥用價值高,因此解析這些活性黃酮的生物合成途徑并闡明其調控機制,將為黃芩的育種或用合成生物學技術異源生產這些活性物質提供理論指導。
3.1 黃芩的組織培養
目前,黃芩的組織培養研究主要集中在愈傷組織的誘導、愈傷組織的分化與再生、細胞懸浮培養以及多倍體誘導[46]。如劉建霞等[47]以黃芩幼苗的莖段為材料,研究了不同的激素濃度配比對黃芩外植體誘導愈傷組織、再生芽和根的影響,發現黃芩莖段在MS+6-BA 1.0 mg/L+NAA 0.5 mg/L 的培養基中愈傷組織的誘導效果最佳;在MS+6-BA 0.5 mg/L+NAA 0.5 mg/L 的培養基中黃芩莖段的再生芽的分化效果最好;在1/2 MS+NAA 0.4 mg/L 的培養基中,黃芩莖段再生根的分化效果最好[47]。由于懸浮培養細胞具有次生代謝產物含量高和抗氧化活性較強的特點,韓淑蘭[48]建立了黃芩細胞懸浮培養體系,并對黃芩苷等次生代謝產物含量進行測定,發現黃芩苷等次生代謝產物在懸浮培養20 d 時獲得了最高的產量78.31 mg/g 干重。李富雄等[49]利用秋水仙素誘導了黃芩的同源四倍體。于輝等[50]測定了四倍體黃芩和二倍體黃芩中的黃芩苷和漢黃芩苷的含量,結果表明四倍體黃芩中黃芩苷和漢黃芩苷含量顯著高于二倍體,由二倍體的12.83%、3.35%增為20.69%、6.23%,分別提高了61.26%和85.97%。
作者研究團隊也成功建立了黃芩再生體系,將新鮮健康葉片滅菌后置于配比為MS + 3% sucrose+ 2.0 mg/L 6-BA + 0.2 mg/L IAA 的愈傷誘導培養基中,暗培養5 周,可觀察到有愈傷組織長出;將得到的愈傷組織置于配比為MS + 3% sucrose + 0.1 mg/L 6-BA+0.5 mg/L PP333 的不定芽誘導培養基中,光照時間為16 h/24 h,光培養60 d 后,可以得到不定芽;將不定芽從愈傷組織上切下,置于配比為MS + 2%sucrose + 0.17 g/L K2HPO4+ 0.5 mg/L PP333 的生根培養基中,光照時間16 h/24 h,光培養30 d 后,可以長出白色的根,得到完整的黃芩再生植株(圖3)。

圖3 黃芩的再生植株Fig. 3 Regenerated plants of S. baicalensis
3.2 黃芩的毛狀根培養系統
由于發根農桿菌的Ri 質粒誘導的毛狀根具有生長迅速、激素自養、生長條件簡單、次生代謝產物含量高且穩定、分化程度高和不易變異等特點,毛狀根培養系統在藥用植物代謝途徑的解析、生產高價值的次生代謝產物以及生產重組蛋白等方面得到廣泛應用[51-52]。黃芩中,利用毛狀根系統進行代謝工程的相關研究已有許多報道,如通過利用毛狀根系統進行基因工程改造,將次生代謝產物合成相關基因或調控的轉錄因子引入毛狀根來調節次生代謝產物的產生。Kim 等[53]使用發根農桿菌介導的毛狀根系統分別過表達肉桂酸4-羥化酶(cinnamate 4-hydroxylase,C4H)和4-香豆酰輔酶A連接酶(4-coumaroyl: CoA-ligase,4CL)基因,通過HPLC 檢測黃芩苷、黃芩素和漢黃芩素含量發現,與對照組相比,過表達SbC4H的毛狀根中,黃芩苷的含量從39.9 mg/g 增加到44.1 mg/g 干重,增加了1.2-1.4 倍,黃芩素和漢黃芩素分別從27.1 mg/g 增加到62.2 mg/g,4.7 mg/g 增加到10.8 mg/g 干重,分別增加了1.4-3.2 倍和1.6-3.4 倍;在過表達Sb4CL的毛狀根中黃芩素是對照組的2.4 倍,漢黃芩素是對照組1.5-2.5 倍。Park 等[54]通過毛狀根體系建立了黃芩的查耳酮異構酶(chalcone isomerase,CHI)基因過表達和SbCHIRNAi 沉默的轉基因毛狀根系,通過HPLC 對黃芩素、黃芩苷和漢黃芩素的含量進行測定,結果表明,SbCHI過表達毛狀根的黃芩素、黃芩苷和漢黃芩素分別是對照組的1.8-2.5 倍、2.4-3.4倍和1.2-3 倍,SbCHIRNAi 毛狀根系的黃芩素、黃芩苷和漢黃芩素相比對照組分別減少了13%-80%、4.6%-50%和22%-85%。Park 等[55]通過在毛狀根培養物中過表達玉米的轉錄因子Lc(ZmLc)和擬南芥的轉錄因子PAP1(AtPAP1)可以增強黃芩苷、黃芩素和漢黃芩素的含量。
毛狀根系統還可以用于表征新基因、新轉錄調控因子的功能以及解析未知的代謝途徑。2016、2018 和2019年,Zhao 等[56-58]找到了可能參與根特異黃酮(黃芩素、漢黃芩素)合成途徑中相關酶的候選基因,通過農桿菌介導的轉化分別得到這些基因的RNAi 沉默的轉基因毛狀根,結果表明根特異黃酮的含量會隨著這些基因表達量的減少而下降,驗證了II 型黃酮合成酶(flavone synthase II, FNSII-2)、肉桂酰輔酶A 連接酶(cinnamate CoA ligase, CLL-7)、黃酮6 位羥化酶(flavone 6-hydroxylase,F6H)、黃酮8位羥化酶(flavone 8-hydroxylase,F8H)和8 位甲氧基轉移酶(8-O-methyl transferase,PFOMT5)的確參與根特異黃酮合成途徑中4′-脫氧黃酮的合成,解析了黃芩素、漢黃芩素的合成途徑。
作者研究團隊曾比較過不同組織來源的外植體對發根誘導效率的影響。用發根農桿菌A4 去侵染黃芩不同的外植體(子葉、葉片和下胚軸)(圖4-a-c),結果發現只有子葉和葉片能夠產生毛狀根(圖4-d 和e),且葉片誘導而來的毛狀根比子葉誘導而來的毛狀根粗壯,更適合后續毛狀根研究。此外,作者研究團隊還發現將紅色熒光蛋白基因(red fluorescent protein gene,dsRed)轉入黃芩毛狀根,用激發光照射后,紅色熒光十分明顯(圖4-f 和g),非常適合作為黃芩發狀根中的報告基因,用于篩選轉基因陽性毛狀根。

圖4 不同黃芩外植體誘導的毛狀根及轉入紅色熒光蛋白的毛狀根Fig. 4 Hairy roots induced by different S. baicalensis explants and hairy roots transferred with dsRed(red fluorescent protein gene)
4 黃芩的組學研究
黃芩是一種非常重要的藥用植物,然而與其相關的基因組、轉錄組信息較少,限制了對其進行生物學研究。2016年,黃芩的花和毛狀根的轉錄組已有發表[57],相關的轉錄組數據已上傳NCBI 的SRA(Sequence Read Archive)數據庫(https://www.ncbi.nlm.nih.gov/bioproject/PRJNA300475)。隨著三代測序技術發展,利用Illumina 和PacBio 測序策略和HIC 技術,黃芩染色體水平的高質量全基因組序列已有報道[56],數據已上傳至國家基因組科學數據中 心(National Genomics Data Center)(https://bigd.big.ac.cn/gwh/Assembly/10400/show)。這是唇形科中第一個完成染色體水平全基因組測序的植物,研究者還從黃芩的花、花蕾、葉、莖、根以及JA 處理的根中提取樣品進行了RNA-seq(https://www.ncbi.nlm.nih.gov/sra/SRX4549232),輔助基因組注釋,共注釋了28 930 個基因,在闡明黃芩的黃芩素和漢黃芩素的合成途徑以及相關酶基因的發現及基因組進化方面取得了很大的進展。此外,還有研究報道了黃芩和半枝蓮的染色體水平基因組(https://ngdc.cncb.ac.cn/gsa/browse/CRA001730),并證實了半枝蓮和黃芩的分化時間為13.28 Mya,通過比較基因組發現二者染色體端粒比例相似,半枝蓮的染色體的重排和擴張可能是近期長末端重復序列的插入導致,為黃芩和半枝蓮的育種、進化、化學多樣性和基因組編輯研究提供重要見解[59]。黃芩的代謝組信息也已有報道[60]。
5 黃芩代謝生物學
5.1 黃芩中黃酮合成途徑的解析
研究表明,黃芩中存在地上部分和地下部分兩條黃酮合成途徑。地上部分(莖、葉和花)主要合成野黃芩素和野黃芩苷,為經典的黃酮合成途徑,普遍存在于高等植物中;地下部分(根)主要合成黃芩素、漢黃芩素、黃芩苷和漢黃芩苷,地下部分的黃酮合成途徑目前只在黃芩中發現,是黃芩特有的,也稱根特異黃酮(root-specific flavones, RSF)合成途徑[56-58](圖5)。

圖5 黃芩的地上地下兩條黃酮合成途徑Fig. 5 Two flavonoid biosynthetic synthesis pathways of S. baicalensis in aerial and root parts
經典黃酮途徑從莽草酸途徑生成的苯丙氨酸開始,苯丙氨酸在苯丙氨酸解氨酶(phenylalanine ammonia lyase, PAL)催化下形成肉桂酸(cinnamic acid)[61];肉桂酸在肉桂酸羥化酶(C4H)作用下生成對香豆酸(p-coumaric acid)[62];對香豆酸經過4-香豆酰輔酶A 連接酶(4CL)催化生成香豆酰輔酶A(p-coumaroyl CoA)[63];香豆酰輔酶A 和丙二酰輔酶 A 在查耳酮合成酶(chalcone synthase, CHS)作用下生成柚皮素查耳酮(naringenin chalcone);柚皮素查耳酮在查耳酮異構酶(CHI)催化下生成柚皮素(naringenin)[64];柚皮素在黃酮合成酶(flavone synthase II, SbFNSII-1)的催化下生成芹菜素;芹菜素在黃酮6 位羥化酶(SbF6H)的作用下生成野黃芩素[57-58,65]。
根特異黃酮合成途徑同樣起始于苯丙氨酸,在SbPAL 作用下生成肉桂酸。隨后,肉桂酸在肉桂酸輔酶A 連接酶(SbCLL-7)催化下形成肉桂酰輔酶 A(cinnamoyl CoA);肉桂酰輔酶A 在松屬素查耳酮合成酶(pinocembrin-chalcone synthase, SbCHS-2)作用下生成松屬素查耳酮(pinocembrin chalcone);松屬素查耳酮在 CHI 的作用下產生松屬素(pinocembrin);松屬素在黃酮合成酶(SbFNS II-2)催化下生成白楊素(chrysin);白楊素在黃酮 6 位羥化酶或黃酮 8 位羥化酶(SbF8H)作用下分別生成黃芩素(baicalein)或去甲漢黃芩素(norwogonin);去甲漢黃芩素最終在8 位氧甲基轉移酶(SbOMT)作用下生成漢黃芩素[56-58]。黃芩素和漢黃芩素在黃酮類7-O-葡萄糖醛酸基轉移酶(UBGAT)的催化下生成黃芩苷和漢黃芩苷[66]。
5.2 黃芩中黃酮合成途徑的調控
代謝途徑不僅受內部基因的調控,還受外部因素影響。環境因子(光質、溫度和水分)和誘導子(生物誘導子和非生物誘導子)同樣能調控黃芩中活性黃酮的積累[65]。有研究表明,光照和黑暗兩個條件下對黃芩懸浮細胞進行培養發現,光照顯著促進了黃芩苷和黃芩素的積累,可能是通過誘導PAL、黃芩素-7-O-葡萄糖醛酸基轉移酶(UBGAT)基因的表達,或通過內源激素進行調控[67-68]。不同光源(紅色、藍色、白色)對黃芩幼苗的根、莖和葉進行照射后發現,白光處理的黃芩幼苗根部的黃芩苷、黃芩素和漢黃芩素含量最高[69]。另有研究表明,黑暗條件有利于黃芩發狀根的生長和黃芩苷的積累[70]。不同溫度培養對黃芩愈傷組織中黃芩苷含量也有顯著影響,在一定范圍內黃芩苷的含量隨著溫度增加而增加,20℃培養時黃芩苷含量最高,高溫35℃或較低溫度(低于15℃)都對黃芩苷的合成產生了一定的抑制,可能是影響了黃芩苷合成途徑中關鍵酶的活性[71]。此外,短期、輕中度干旱脅迫能夠不同程度增強PAL、C4H、4CL 和CHS 酶活性,促進黃芩苷積累,長期、重度干旱脅迫會抑制這4 種酶的活性,導致黃芩苷的合成受到抑制[72]。
誘導子常被用來促進次生代謝產物的積累,如曲霉和米曲霉等真菌誘導子能提高黃芩毛狀根中黃芩苷的含量[73]。茉莉酸甲酯(methyl jasmonate,MeJA)能夠通過上調黃酮合成途徑相關基因從而促進黃芩懸浮細胞、愈傷組織和發根中黃芩苷的積累[57,62,74-75]。在黃芩愈傷組織培養基中加入一定濃度的水楊酸會抑制愈傷組織生長,但能促進黃芩苷的積累[76]。添加外源植物激素如6-芐氨基嘌呤(N-(phenylmethyl)-9H-purin-6-amine, 6-BA)和萘乙酸(1-naphthylacetic acid, NAA)培養黃芩愈傷組織,也能夠提高黃芩苷含量。稀土元素如鑭(La),能夠以茉莉酸(jasmonic acid, JA)為信號分子誘導黃芩幼苗積累黃芩苷[77]。高濃度砷抑制黃芩植物黃芩苷和漢黃芩苷的生成,但促進了黃芩素、漢黃芩素和千層紙素A 的生成[78]。
5.3 黃芩中黃酮合成途徑的進化
進化分析表明,黃芩的根特異黃酮合成途徑是從地上部分的經典黃酮合成途徑進化而來,該途徑進化在黃芩屬和鼠尾草屬(Salvia)的分化后形成(<32.7 Mya),該途徑中只有SbCHI基因僅有一個基因座,且在地上部分和地下部分黃酮合成途徑中功能相同,其他關鍵酶基因分別經歷點突變(SbCLL-7)、串聯增殖以及新功能化(SbCHS-2、SbFNSII-2、SbF8H和SbPFOMT)、片段復制(SbFNSII-2)和反轉錄復制(SbF6H)進化而來[79]。
PAL 催化苯丙氨酸脫氨基生成反式肉桂酸和氨,是連接初級代謝和次級代謝之間的關鍵酶之一[61]。Xu 等[62]使用 cDNA 末端快速擴增(RACE)技術從黃芩中克隆了3 個編碼苯丙氨酸解氨酶亞型(SbPAL1、SbPAL2和SbPAL3)的全長 cDNA。黃芩和半枝蓮的比較基因組分析顯示,黃芩中的PAL基因數(5 個)與半枝蓮的PAL(4 個)相比要多,說明這些基因的特異性擴增可能是在黃芩和半枝蓮物種形成后通過串聯復制發生的(<13.28 Mya)[59]。
CLL 是4CL 的異構體。基于根RNA-seq 數據庫,Zhao 等[56]從黃芩中鑒定并克隆了5 個CLL基因的全長cDNA(SbCLL-1、SbCLL-5、SbCLL-6、SbCLL-7,SbCLL-8);純化的重組蛋白SbCLL-1 和SbCLL-5 可以催化肉桂酸、4-香豆酸和咖啡酸與CoA 反應,而SbCLL-7 只能以肉桂酸為底物添加CoA;比較基因組分析表明,SbCLL-1和SbCLL-7在唇形科其他物種(丹參、一串紅和芝麻)中具有同源性,但丹參、一串紅和芝麻中的CLL-7對肉桂酸、4-香豆酸或咖啡酸沒有活性,表明SbCLL-7對肉桂酸的特異性活性在黃芩近緣種的同源基因中沒有共享,說明黃芩中的CLL基因的點突變可能發生在黃芩、丹參和一串紅分化后(<32.7 Mya)。
Zhao 等[57]還從黃芩中鑒定并克隆了兩個CHS基因的全長cDNA(SbCHS-1和SbCHS-2)。SbCHS-1在花中特異性表達,可能與經典黃酮和花青素的生物合成有關;SbCHS-2在根中高度表達,編碼一種催化松屬素查耳酮生成的酶。基于基因組分析,SbCHS-2的基因組區域發生了串聯復制事件,導致SbCHS-2產生了5 個相鄰的基因拷貝(SbCHS2.1-SbCHS2.6)。SbCHS-1和SbCHS-2的新功能化是隨著鼠尾草屬和黃芩屬(~32.7 Mya)的分化而發生的,原始CHS基因在黃芩中復制,一個拷貝(SbCHS-1)移向假3 號染色體,另一個拷貝(SbCHS-2L)保留在假9 號染色體(約19 Mya)上,并在12 Mya 左右復制(SbCHS2.3)。然后通過串聯復制擴增,產生其他SbCHS-2基因,并且可能在最后一個Mya內新功能化[56]。總之,黃芩的SbCHS-2經歷了幾輪擴增,在物種分化之前出現的重復(SbCHS2L和SbCHS2.3),以及在物種水平分化后出現串聯復制(CHS-2.1、CHS-2.2、CHS-2.4、CHS-2.5和CHS-2.6)。比較基因組表明,在半枝蓮中,CHS-2的復制程度沒有黃芩中那么大[59],這證實了CHS-2的大部分復制是近期內發生的[56],可能是黃芩作為中藥被人為篩選后引起的。
黃芩中,兩種不同類型的FNS 酶(FNSI 和FNSII)通過引入C2 和C3 位的雙鍵來實現黃烷酮到黃酮的轉化。FNSI 屬于酮戊二酸依賴的雙加氧酶(2-oxoglutarate-dependent dioxygenase, 2OGD)超家族,最早在傘形科植物中發現[80],FNSII 是一種廣泛分布于被子植物中的細胞色素P450(CYP450)單加氧酶[81]。一般來說,FNSI 和FNSII 催化具有4′-OH基團黃烷酮類,如柚皮素、圣草酚和甘草素轉化為相應的黃酮[82]。Zhao 等[57]首次報道了黃芩中具有新功能的FNSII(SbFNSII-2),該FNSII 是專門合成4′-脫氧黃酮所必需的,此外,黃芩的地上部分還存在FNSII 亞型(SbFNSII-1),能將柚皮素轉化為芹菜素。基因組分析表明,有兩個SbFNSII-1位點是唇形科分化后(<42.7 Mya)發生片段復制的一部分,然而,黃芩基因組中僅存在一個SbFNSII-2位點,表明SbFNSII-2是由SbFNSII-1串聯復制產生,并且是在鼠尾草屬和黃芩屬分化(<32.7 Mya)之后發生的新功能化[56]。黃芩和半枝蓮比較基因組分析表明,在黃芩與半枝蓮共線區域可以檢測到FNSII-2的一個直系同源基因,以及FNSII-1和FNSII-2的串聯復制[59]。這表明黃芩屬中FNS-II復制早于物種分化,黃芩屬中4 ′-脫氧黃酮化合物的進化十分保守[79]。
黃芩中的兩個CYP450 家族成員負責黃酮化合物6 位和8 位的羥基化(分別是SbF6H 和SbF8H),SbF6H(CYP82D1.1)可以利用4′-羥基黃酮化合物和4′-脫氧黃酮化合物(分別為芹菜素和松屬素)在地上部分和根中分別產生野黃芩素和黃芩素[58]。黃芩的5 號染色體上只有一個SbF6H位點,該位點可能來源于1 號染色體上CYP82D基因的復制和逆轉錄轉座作用,而SbF8H則來源于1 號染色體上串聯復制和新功能化[56]。
黃 芩 中 兩 類OMTs(O-methyltransferases) 參與根特異性4′-脫氧黃酮化合物的生物合成。II 型OMT 依賴于Mg2+,也被稱為苯丙素類和類黃酮OMT(PFOMTs),據報道,它們能將甲基轉移到類黃酮的芳香環的羥基上[79]。在黃芩根中,SbPFOMT 2 和5可以有效地O-甲基化黃酮化合物的C6、C8 和C3 位,分別生成單甲氧基黃酮化合物如千層紙素、漢黃芩素、韌黃芩素Ⅱ和金圣草素[56,60]。此外,在黃芩中發現了3 條有功能的I 型OMTs(flavonoid OMTs,SbFOMTs)可 以 修 飾4′- 脫 氧 黃 酮,SbFOMT3 和SbFOMT6 是7-OMT,前者可將黃芩素轉化為7-甲氧基黃芩素;后者可同時修飾黃芩素和去甲漢黃芩素底物;SbFOMT5 能使黃芩素C5、C6 和C7 上的羥基甲基化;SbPFOMT5 與SbFOMT6 組合或SbPFOMT5加SbFOMT5 組合可在酵母中分別產生黃芩黃酮I 和韌黃芩素I[60]。此外,SbPFOMT5 最近在1 號染色體上也經歷了串聯復制和新功能化[56]。
6 黃芩的合成生物學
隨著合成生物學的發展,通過大腸桿菌、酵母等微生物合成植物天然產物成為一種高效、綠色和經濟的方法[83]。已有研究報道在大腸桿菌、酵母和煙草中表達黃酮合成關鍵基因,重建黃酮生物合成途徑,提高活性黃酮的產量。Zhao 等[58]曾在煙草中瞬時表達黃芩中的SbCLL-7、SbCHS-2、SbCHI、SbFNSII-2、SbF6H,得到了黃芩苷和野黃芩素。Li等[84]在大腸桿菌轉入來自不同物種的黃酮合成關鍵基因,圓紅酵母(Rhodotorula toruloides)的PAL、歐 芹(Petroselinum crispum) 的4CL和FNS I、 矮牽 牛(Petunia hybrida) 的CHS、 苜 蓿(Medicago sativa)的CHI以及黃芩截短的F6H,在大腸桿菌中重構了黃芩素和野黃芩素的生物合成途徑,在以0.5 mg/L 的丙苯氨酸和酪氨酸為前體48 h 發酵后,得到了8.5 mg/L 的黃芩素和47.1 mg/L 的野黃芩素;經過優化丙二酰輔酶 A 的利用率后,黃芩素和野黃芩素的搖瓶產量達到了23.6 mg/L 和106.5 mg/L[84]。之后,還開發了順序自組裝酶反應器,利用大腸桿菌中蛋白質-肽相互作用將RtPAL 和Pc4CL 進行自組裝,有效地減少了中間體的積累,大大的提高黃芩素和野黃芩素的產量,黃芩素的搖瓶產量從21.6 mg/L 提高到143.5 mg/L,提高了6.6 倍;野黃芩素的搖瓶產量從84.3 mg/L 提高到120.4 mg/L,提高了1.4 倍;通過發酵罐分批補料,黃芩素和野黃芩素的產量能達到271 mg/L 和288.9 mg/L;此外,還在大腸桿菌中首次實現了從葡萄糖到黃芩素的從頭合成,通過分批補料發酵,黃芩素的產量能達到214.1 mg/L[85]。Qian 等[86]在畢赤酵母中分別引入黃芩根中4′-脫氧黃酮合成中的多個催化酶:PAL、CLL-1、CHS-2、CHI、FNSII-1、F6H、F8H、OMT5,結果發現其產物除了黃芩素、去甲漢黃芩素和漢黃芩素外,首次得到了千層紙素A、6-甲氧基漢黃芩素、6-甲氧基去甲漢黃芩素,表明SbF8H 和SbPFOMT5 具有多種催化作用;此外,他們在畢赤酵母中引入完整的黃芩素的生物合成途徑,并以乙醇為碳源,實現了黃芩素的從頭合成;為了提高黃芩素和千層紙素的產量,他們進一步將黃芩素生物合成途徑分成5個模塊,還構建了一系列的乙醇誘導轉錄信號放大裝置(ESADs)和組成性轉錄信號放大裝置(CSADs)來驅動和平衡模塊,顯著減少了副產物和中間產物的積累,黃芩素和千層紙素A 的產量達到報道過的最高水平401.9 mg/L 和339.5 mg/L。
7 總結與展望
黃芩作為傳統的藥用植物,已有數千年的使用歷史,在現代醫學中,黃芩仍擁有著巨大的開發潛力,如黃芩素能夠很好地結合新型冠狀病毒的3CLpro 靶點,阻止引起全球大流行的新冠病毒的復制[24]。此外,黃芩素和漢黃芩素等還具有良好的抗腫瘤、抗炎、神經保護等作用。目前,黃芩中根特異黃酮的生物合成途徑已經完整解析,全基因組序列和轉錄組數據已發表,這些工作不僅為黃芩后續的研究提供了參考,也為開發利用其他藥用植物提供了借鑒。到目前為止,黃芩黃酮骨架結構合成途徑及A 環上的修飾機制已經研究清楚,但其B 環修飾反應還需進一步研究。如韌黃芩素I、黃芩黃酮I、甘肅黃芩素I、側花黃芩素I,這些黃酮在B 環的2′、3′、5′和6′位置發生羥基化,該羥基化的酶尚未鑒定,且其中部分黃酮(韌黃芩素I 和黃芩黃酮I)在誘導癌細胞的細胞凋亡方面具有比黃芩素更強的細胞毒性[60],其生物合成途徑及臨床研究非常值得探索。
除黃芩外,黃芩屬中還有許多其他入藥的植物,如半枝蓮(Scutellaria barbata)在韓國和朝鮮常被用來治療咽喉腫痛、水腫和痔瘡和蛇咬傷以及一些癌癥;側花黃芩(Scutellaria lateriflora)在北美被用來治療癲癇、焦慮等神經疾病;韓信草(Scutellaria indica)在中國、韓國和印度被用來解毒、鎮痛和促進血液循環[65]。黃芩屬植物除含有黃酮化合物外,還含有豐富的二萜和生物堿,這些化合物具有良好的抗腫瘤活性,其生物合成途徑也有待研究。飛速發展的第三代測序技術、代謝物分析技術、基因編輯技術將有助于我們進一步研究黃芩屬植物活性成分的生物合成和調控,以及黃芩屬植物的多樣化和進化。
猜你喜歡
四川蠶業(2021年3期)2021-02-12 02:38:46
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中成藥(2018年11期)2018-11-24 02:57:00
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
中國衛生(2016年3期)2016-11-12 13:23:26
湖南師范大學自然科學學報(2015年1期)2015-02-27 14:50:04