agr 系統(tǒng)在單核增生細(xì)胞李斯特菌耐藥及生物被膜形成中的作用
2023-03-07 12:56:24任思雨姜聰一于濤康瑞姜曉冰
生物技術(shù)通報(bào) 2023年2期
任思雨 姜聰一 于濤 康瑞 姜曉冰
(1. 河南師范大學(xué)生命科學(xué)學(xué)院,新鄉(xiāng) 453007;2. 新鄉(xiāng)學(xué)院生命科學(xué)與基礎(chǔ)醫(yī)學(xué)學(xué)院,新鄉(xiāng) 453000)
單核細(xì)胞增生李斯特菌(Listeria monocytogenes,Lm)是一種常見(jiàn)的食源性致病菌,廣泛存在于食品以及食品加工環(huán)境中[1]。Lm 是人畜共患病李斯特菌病(listeriosis)的病原菌,可造成孕婦、新生兒和老年人等免疫力低下人群的感染。隨著抗生素、消毒劑等在臨床和食品加工過(guò)程中的廣泛應(yīng)用,Lm對(duì)這些抗菌藥物的耐藥性逐漸增強(qiáng)[2]。Lm 能夠在不同的食品接觸面形成生物被膜,處于被膜狀態(tài)的Lm 對(duì)外界不利條件的抵抗力更強(qiáng),因此普通的消毒殺菌難以有效清除Lm 生物被膜。Lm 生物被膜在食品加工環(huán)境中的存在增加了食品被Lm 污染的幾率,對(duì)食品安全造成了嚴(yán)重的威脅[3]。
群體感應(yīng)(quorum sensing, QS)是細(xì)菌細(xì)胞間化學(xué)通訊的過(guò)程。細(xì)菌產(chǎn)生并釋放一種被稱為自誘導(dǎo)劑的細(xì)胞外信號(hào)分子,自誘導(dǎo)劑的濃度隨著細(xì)胞密度的增加同步增加,當(dāng)達(dá)到一定閾值時(shí)會(huì)啟動(dòng)相關(guān)基因的表達(dá)[4]。QS 參與調(diào)控細(xì)菌多種重要生理行為[5],如生物發(fā)光、生物被膜的形成、次級(jí)代謝產(chǎn)物產(chǎn)生、DNA 攝取能力和毒力因子的產(chǎn)生等。
目前研究者已在Lm 中發(fā)現(xiàn)兩個(gè)QS 系統(tǒng),即LuxS 系統(tǒng)[6]和Agr 系統(tǒng)[7],其中Agr 系統(tǒng)僅存在于一些革蘭氏陽(yáng)性菌中。Lm 的agr是由agrBDCA組成的操縱子,分別編碼前體加工酶AgrB、前體肽AgrD、組氨酸蛋白激酶AgrC 和反應(yīng)調(diào)控子AgrA。Lm 產(chǎn)生的前體肽AgrD 經(jīng)膜蛋白AgrB 的加工修飾成為具有活性的信號(hào)分子——自誘導(dǎo)肽(autoinducing peptide, AIP)并釋放到胞外。當(dāng)AIP 達(dá)到一定濃度后會(huì)與細(xì)胞膜上的AgrC 結(jié)合同時(shí)觸發(fā)級(jí)聯(lián)磷酸化反應(yīng),磷酸化的AgrA 與下游的靶基因啟動(dòng)子結(jié)合進(jìn)而調(diào)控相關(guān)基因的表達(dá)(圖1)。研究發(fā)現(xiàn),agrA的缺失導(dǎo)致Lm 菌株生物被膜形成能力降低[8];agrD可能參與Lm 的侵襲、生物被膜形成以及幾丁質(zhì)酶的表達(dá)[9-11]。agrC在Lm 生物被膜形成中的作用仍不清楚。此外,agr系統(tǒng)在Lm 對(duì)抗菌藥物耐藥中的作用也尚未見(jiàn)報(bào)道。
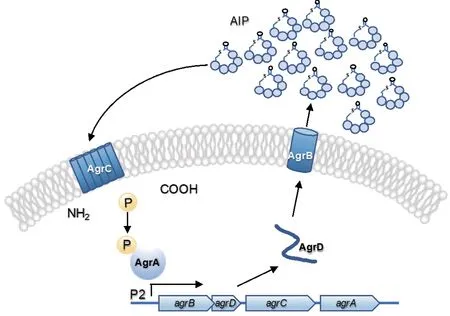
圖1 Lm 中的agr 系統(tǒng)Fig. 1 agr system in L. monocytogenes
本研究以Lm 標(biāo)準(zhǔn)菌株EGD-e 為研究對(duì)象構(gòu)建agr系統(tǒng)基因缺失突變株ΔagrB/D、ΔagrC和ΔagrA,通過(guò)比較野生株和突變株在耐藥性、生物被膜形成能力和運(yùn)動(dòng)性方面的差異,調(diào)查agr系統(tǒng)在Lm 對(duì)抗菌藥物耐藥及生物被膜形成中的作用。
1 材料與方法
1.1 材料
Lm 標(biāo)準(zhǔn)菌株EGD-e 由華中師范大學(xué)羅勤教授惠贈(zèng);大腸桿菌DH5α 感受態(tài),BIOGENE;穿梭質(zhì)粒pMAD,本實(shí)驗(yàn)保存;突變株EGDeΔagrC,本實(shí)驗(yàn)室保存;苯扎氯銨(benzalkonium chloride, BC),國(guó)藥集團(tuán)化學(xué)試劑有限公司;結(jié)晶紫,天津市大茂化學(xué)試劑廠;草酸銨,北京化工廠,細(xì)菌基因組DNA 提取試劑盒,RealMaster Mix(SYBR Green)試劑盒,天根生化科技有限公司;高保真PCR 酶、Taq 酶和dNTPs,TaKaRa 公司;細(xì)菌RNA 提取試劑盒,Omega 公司;質(zhì)粒小提試劑盒、膠回收試劑盒、限制性內(nèi)切酶、T4 DNA 連接酶,Thermo 公司。
腦心浸液培養(yǎng)基(brain heart infusion, BHI),OXOID 公司;胰蛋白胨大豆肉湯培養(yǎng)基(tryptic soy broth, TSB),北京索萊寶科技有限公司;LB 培養(yǎng)基(g/L):酵母提取物5.0,胰蛋白胨10.0,NaCl 10.0;0.5%草酸銨-結(jié)晶紫溶液:A 液:1 g 結(jié)晶紫溶于20 mL 95%的乙醇溶液中,B 液:用80 mL 蒸餾水溶解0.8 g 草酸銨,使用時(shí)將A 液與B 液混勻,靜置72 h。
基因擴(kuò)增儀,北京東勝創(chuàng)新生物科技有限公司;凝膠成像分析系統(tǒng)、電轉(zhuǎn)儀、酶標(biāo)儀,Bio-Rad 公司;實(shí)時(shí)熒光定量PCR 儀,ABI 公司;恒溫培養(yǎng)箱,北京科偉永興儀器有限公司;微生物全自動(dòng)生長(zhǎng)曲線分析儀,Oy Growth Curves Ab 有限公司;倒置顯微鏡,Leica 公司。
1.2 方法
1.2.1 基因缺失突變株的構(gòu)建 使用細(xì)菌基因組試劑盒提取Lm 野生株EGD-e 的DNA,利用 SOE-PCR技 術(shù)(splicing by overlap extension PCR, SOE-PCR),以EGD-e 基因組DNA 為模板使用引物對(duì)P1 和P2、P3 和P4(引物序列見(jiàn)表1)分別擴(kuò)增目的基因的上、下游同源臂,融合后得到的長(zhǎng)片段經(jīng)酶切純化后與載體pMAD 連接。將重組質(zhì)粒轉(zhuǎn)化至大腸桿菌DH5α 中,篩選出陽(yáng)性克隆菌株并測(cè)序。將測(cè)序正確的重組質(zhì)粒電轉(zhuǎn)至EGD-e 感受態(tài)中,在紅霉素和高溫的雙重壓力下篩選出基因缺失突變株。

表1 PCR 擴(kuò)增引物序列Table 1 Primer sequences for PCR amplification
1.2.2 最低抑菌濃度(minimum inhibitory concentration, MIC)值測(cè)定 根據(jù)臨床實(shí)驗(yàn)室標(biāo)準(zhǔn)協(xié)會(huì)(clinical and laboratory standards institute, CLSI)推薦的微量肉湯稀釋法測(cè)定菌株對(duì)10 種抗菌藥物的MIC 值。
1.2.3 生長(zhǎng)曲線測(cè)定 將培養(yǎng)至對(duì)數(shù)期(OD600為0.5-0.6)的菌液按1∶100 的比例分別轉(zhuǎn)接至新鮮的BHI 和含有1 μg/mL BC 的BHI 中。取200 μL 稀釋菌液轉(zhuǎn)移至100 孔的檢測(cè)板中,置于微生物全自動(dòng)生長(zhǎng)曲線分析儀中,37℃培養(yǎng)48 h,每15 min 測(cè)定一次OD600。
1.2.4 致死曲線測(cè)定 將待測(cè)菌株培養(yǎng)至對(duì)數(shù)期(OD600為0.5-0.6),取1 mL 菌液離心(8 000 r/min,5 min),離心后棄去上清,使用新鮮的5 mL BHI+BC(16 μg/mL)培養(yǎng)基重懸菌體,37℃培養(yǎng),分別在0、10、20、30 和40 min 時(shí)取100 μL 的菌液進(jìn)行梯度稀釋涂板,在37℃培養(yǎng)24 h 后計(jì)數(shù)。
1.2.5 生物被膜定量及觀察 從平板上挑取單克隆接種于新鮮的BHI 培養(yǎng)基中37℃振蕩過(guò)夜培養(yǎng)。次日取過(guò)夜菌液按1%的比例接種至新鮮的BHI 培養(yǎng)基內(nèi),待其培養(yǎng)至對(duì)數(shù)期(OD600為0.6-0.8)時(shí),取1 mL 菌液離心(8 000 r/min, 5 min)、重懸,取重懸菌液按1%的比例稀釋至BHI 中,吸取1.2 mL 稀釋后的菌液加入到24 孔板中,分別在37℃、20℃和4℃下培養(yǎng)生物被膜。
生物被膜的定量:小心棄去孔中的培養(yǎng)基,使用無(wú)菌PBS 緩沖液沖洗24 孔板以去除浮游細(xì)菌,室溫下風(fēng)干40 min 后,向每個(gè)孔中加入的0.5%結(jié)晶紫染液,溶液染色30 min;棄去孔中染液,用無(wú)菌水洗滌4-6 次,室溫干燥;加入95%的乙醇在37℃下孵育30 min,最后使用酶標(biāo)儀檢測(cè)OD595值。
測(cè)定OD595后,棄去孔中的脫色液并置于室溫下干燥,直接使用倒置顯微鏡(放大倍數(shù)為60 倍)觀察生物被膜,并拍照記錄。
1.2.6 運(yùn)動(dòng)性測(cè)定 采用軟平板法測(cè)定菌株的運(yùn)動(dòng)性。群集運(yùn)動(dòng)性軟平板含0.5%瓊脂,泳動(dòng)運(yùn)動(dòng)性平板含0.3%瓊脂。待測(cè)菌株在TSB 培養(yǎng)基中于37℃振蕩過(guò)夜培養(yǎng);次日按1%的比例接種至新鮮的TSB 培養(yǎng)基中,繼續(xù)培養(yǎng)至對(duì)數(shù)期,取1 μL 稀釋后的菌液接種于平板中心,在室溫下放置30 min 待菌液吸收后,于25℃靜置培養(yǎng)24 h,之后用游標(biāo)卡尺測(cè)定菌圈直徑。
1.2.7 RT-qPCR 檢測(cè)相關(guān)基因轉(zhuǎn)錄水平 按照細(xì)菌RNA 提取試劑盒提取細(xì)菌總RNA。使用反轉(zhuǎn)錄試劑盒將總RNA 反轉(zhuǎn)錄為cDNA。以cDNA 為模版,使用RealMaster Mix 試劑盒進(jìn)行RT-qPCR,以16S rRNA 為參照基因。以野生株EGD-e 的基因轉(zhuǎn)錄水平為對(duì)照。
1.2.8 數(shù)據(jù)統(tǒng)計(jì)分析 使用SPSS 23.0 統(tǒng)計(jì)軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行t檢驗(yàn)分析,P<0.05 則差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 基因缺失突變株的構(gòu)建與分子鑒定
使用引物1-4 分別擴(kuò)增agrB/D和agrA的上下游同源臂,通過(guò)SOE-PCR 得到融合片段(圖2-A)。融合片段與pMAD 質(zhì)粒連接獲得重組質(zhì)粒,雙酶切驗(yàn)證結(jié)果如圖2-B 所示。將測(cè)序結(jié)果正確的重組質(zhì)粒電轉(zhuǎn)至Lm 感受態(tài)細(xì)胞,高溫和抗生素雙重壓力篩選突變株(圖2-C)。測(cè)序結(jié)果顯示成功缺失agrB/D和agrA,獲得基因缺失突變株ΔagrB/D和ΔagrA。
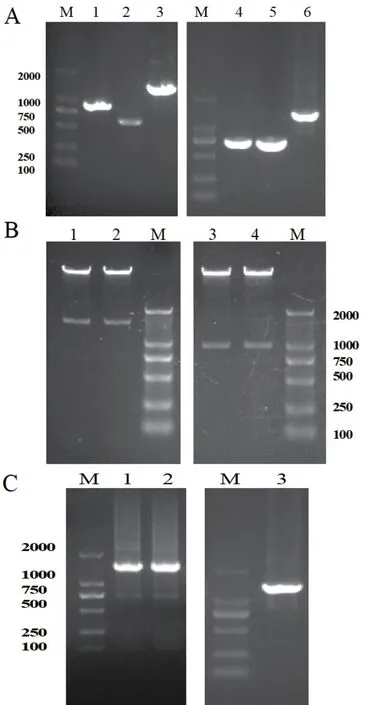
圖2 agrA 和agrB/D 基因缺失株的構(gòu)建Fig. 2 Construction of ?agrA and ?agrB/D
2.2 抗菌物質(zhì)對(duì)Lm菌株的MIC值
如表2 所示,3 株缺失突變株對(duì)氨芐西林、頭孢噻吩、紅霉素、卡那霉素、四環(huán)素、氯霉素、乳酸鏈球菌素以及BC 的MIC 值與野生株無(wú)差異。與野生株相比,ΔagrA對(duì)頭孢噻肟的MIC 值下降,ΔagrB/D和ΔagrC對(duì) 環(huán) 丙 沙 星 的MIC 值 降 至0.5 μg/mL。
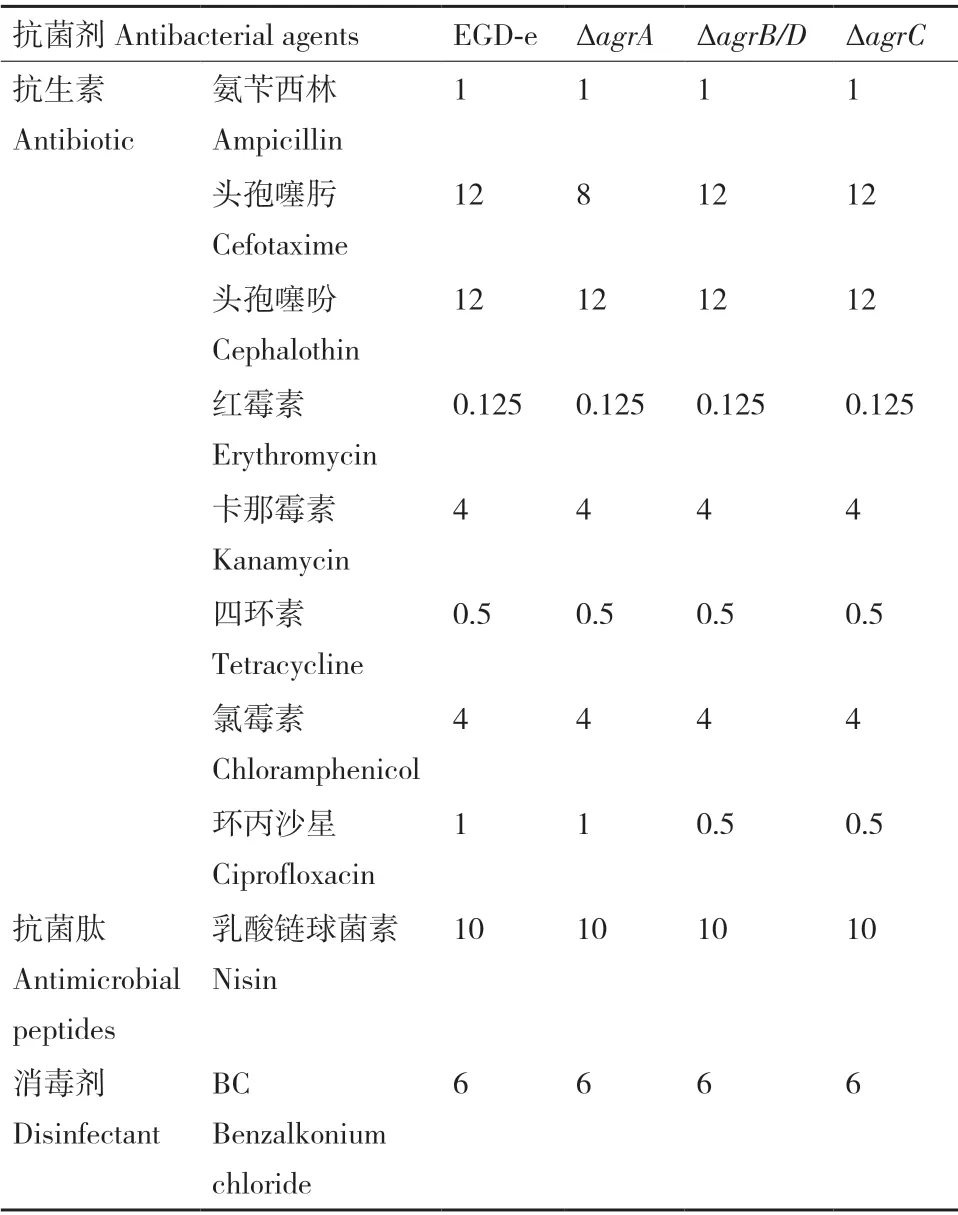
表2 抗菌物質(zhì)對(duì)Lm 菌株的MIC 值Table 2 MICs of antimicrobial agents against L. monocytogenes
2.3 BC脅迫下Lm的生長(zhǎng)曲線和致死曲線
生長(zhǎng)曲線結(jié)果如圖3 所示。野生株與3 株突變株在BHI 中的生長(zhǎng)無(wú)差異;當(dāng)BC 濃度為1 μg/mL時(shí),與野生株相比,ΔagrC的生長(zhǎng)完全受到抑制;ΔagrB/D的延遲期明顯變長(zhǎng);而ΔagrA的生長(zhǎng)延遲期則明顯變短(表3)。
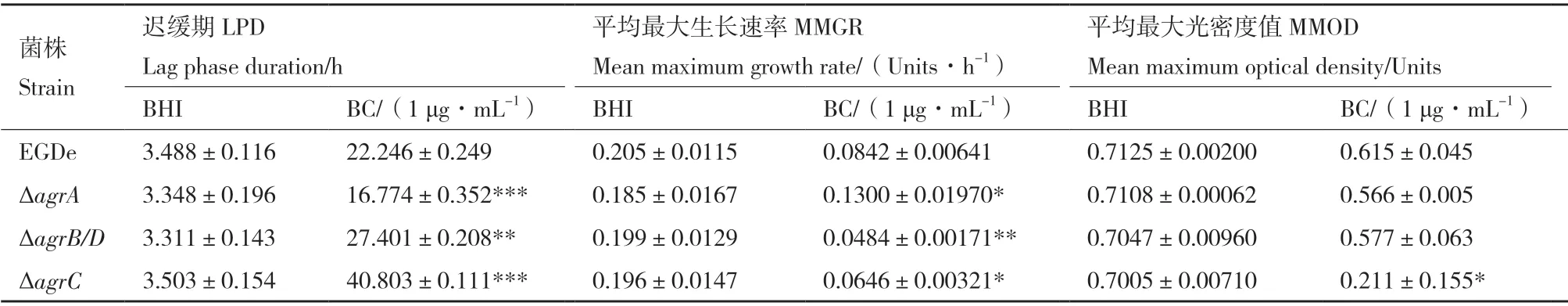
表3 生長(zhǎng)曲線分析Table 3 Growth curve analysis

圖3 Lm 的生長(zhǎng)曲線Fig. 3 Growth curves of L. monocytogenes
由圖4 可知,當(dāng)BC 濃度為16 μg/mL 時(shí),暴露10 min 后野生株的存活率為49.12%;ΔagrC、ΔagrA和ΔagrB/D 的存活率均低于野生株,分別為33.83%、26.29%和14.12%。暴露30 min 后,野生株的存活率為17.9%,而所有突變株的存活率則趨近于0。
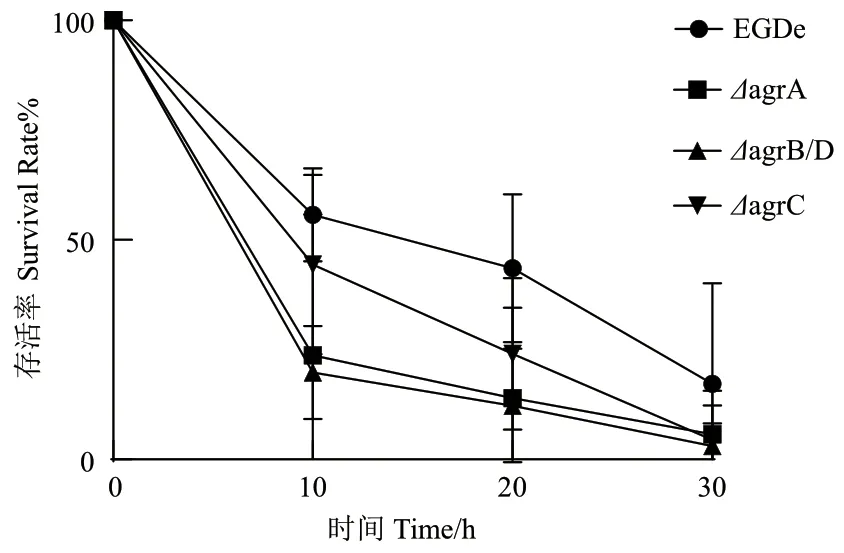
圖4 Lm 對(duì)BC 的致死曲線Fig. 4 Death curves of L. monocytogenes exposed to BC
2.4 Lm生物被膜形成能力
37℃培養(yǎng)條件下Lm 生物被膜形成如圖5 和表4 所示。在培養(yǎng)1、2 和3 d 的過(guò)程中,ΔagrA和ΔagrB/D在各個(gè)時(shí)間段的生物被膜形成量均明顯低于野生株;ΔagrC的生物被膜量在培養(yǎng)3 d 后顯著下降(圖5-A)。倒置顯微鏡觀察培養(yǎng)2 d 后野生株和基因缺失株的被膜結(jié)構(gòu)如圖5-B 所示,野生株形成的生物被膜結(jié)構(gòu)完整致密,生物量多,而agr缺失突變株形成的生物被膜結(jié)構(gòu)疏散,生物量少。
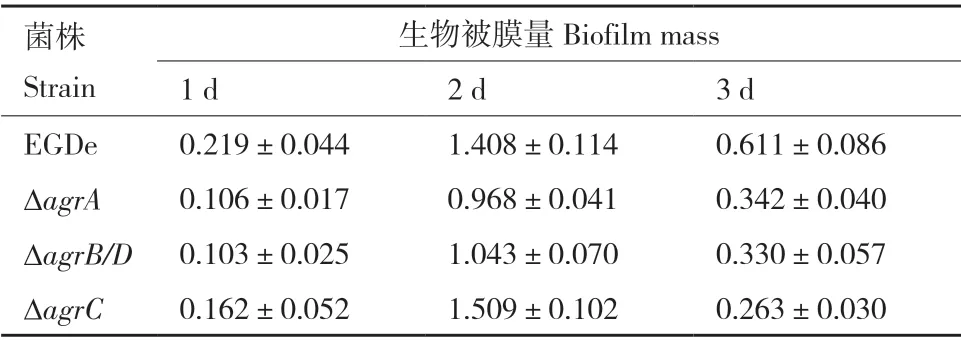
表4 野生株和突變株在37℃下形成的生物被膜量Table 4 Biofilm biomasses of wild type and mutant strains at 37℃
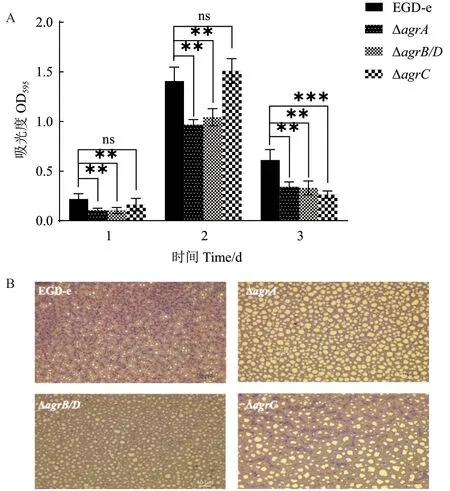
圖5 Lm 菌株在37℃下形成的生物被膜Fig. 5 Biofilm formation of L. monocytogenes strains at 37℃
20℃培養(yǎng)條件下Lm 生物被膜形成如圖6 和表5 所示。只有在培養(yǎng)3 d 后,ΔagrA、ΔagrB/D和ΔagrC的生物被膜形成量均明顯低于野生株。倒置顯微鏡觀察培養(yǎng)6 d 后野生株和基因缺失株的被膜結(jié)構(gòu)如圖6-B 所示,野生株形成的生物被膜結(jié)構(gòu)相對(duì)完整致密,生物量多,而agr缺失突變株形成的生物被膜結(jié)構(gòu)疏散,網(wǎng)狀結(jié)構(gòu)明顯,生物量少。
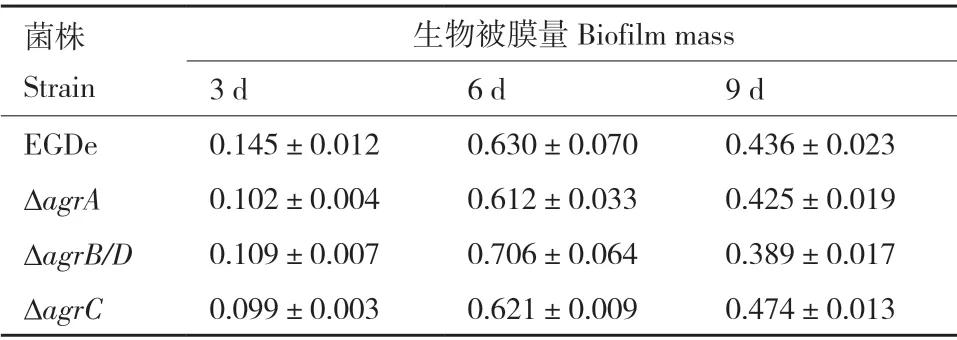
表5 野生株和突變株在20℃下形成的生物被膜量Table 5 Biofilm biomasses of wild type and mutant strains at 20℃
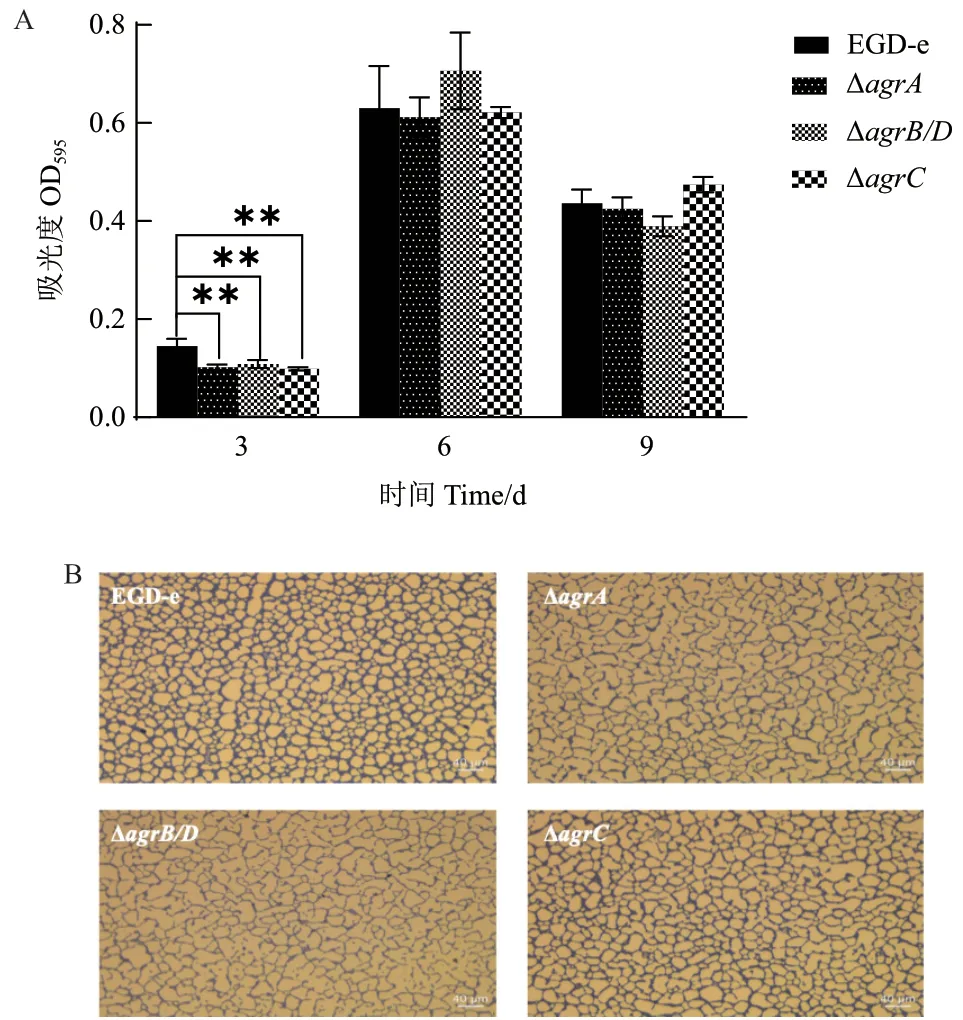
圖6 Lm 菌株在20℃下形成的生物被膜Fig. 6 Biofilm formation of L. monocytogenes strains at 20℃
4℃培養(yǎng)條件下Lm 生物被膜形成如圖7 和表6 所示。在培養(yǎng)10、20 和30 d 的過(guò)程中,ΔagrA、ΔagrB/D和ΔagrC在各個(gè)時(shí)間段的生物被膜形成量均明顯低于野生株(圖7-A)。倒置顯微鏡觀察培養(yǎng)20 d 后野生株和基因缺失株的被膜結(jié)構(gòu)如圖7-B 所示,野生株形成的生物被膜結(jié)構(gòu)完整致密,孔隙少,生物量多,而agr缺失突變株形成的生物被膜結(jié)構(gòu)疏散,孔隙大,生物量少。
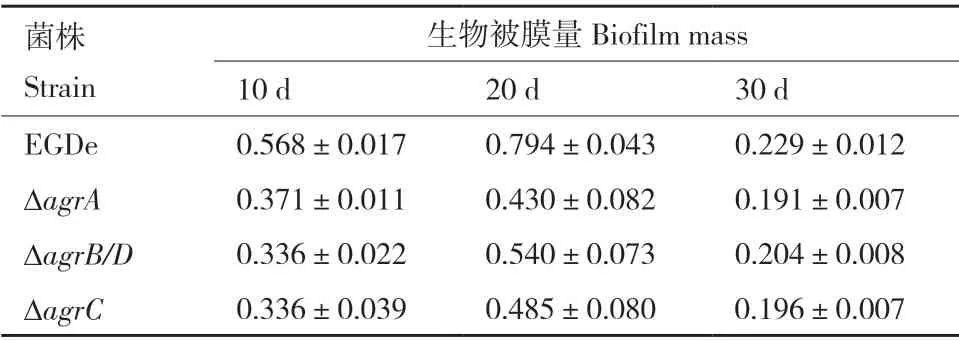
表6 野生株和突變株在4℃下形成的生物被膜量Table 6 Biofilm biomasses of wild type and mutant strains at 4℃
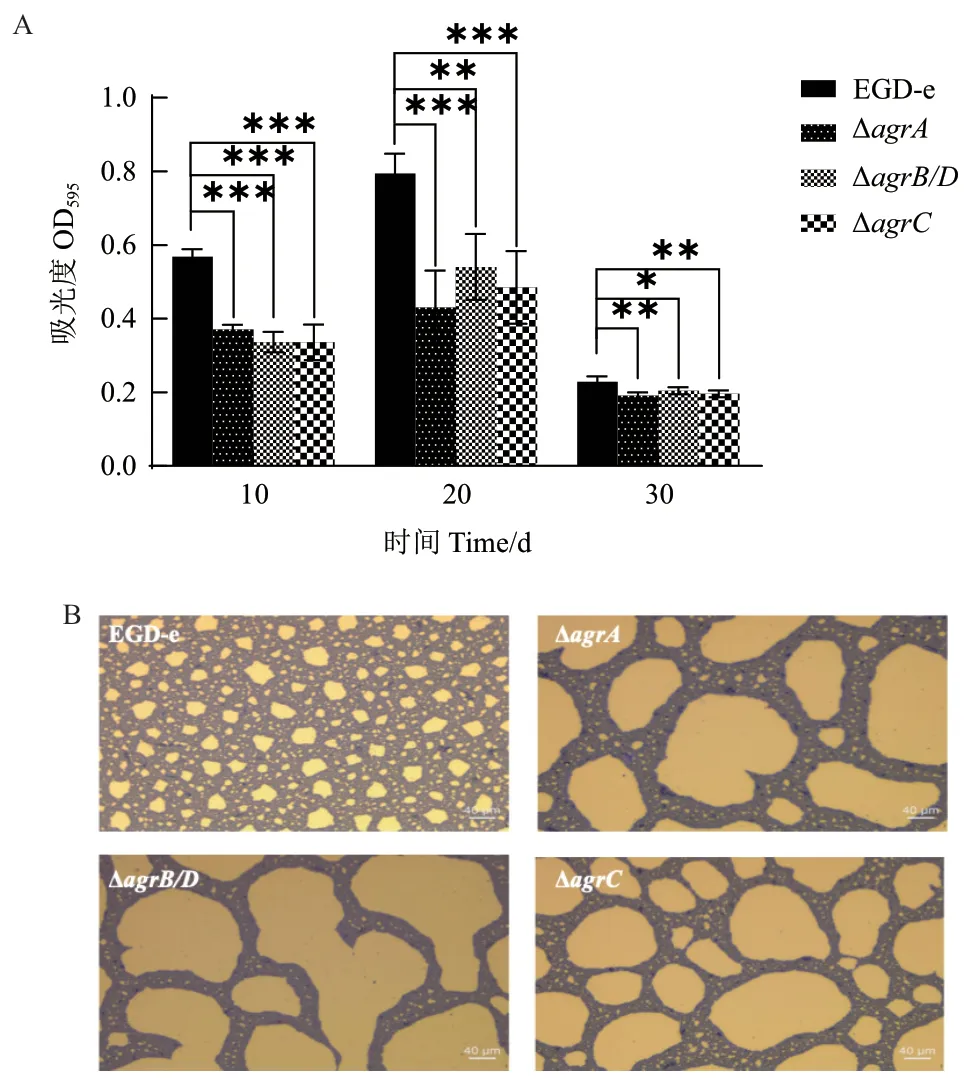
圖7 Lm 菌株在4℃下形成的生物被膜Fig. 7 Biofilm formations of L. monocytogenes strains at 4℃ 20 d
2.5 運(yùn)動(dòng)性檢測(cè)結(jié)果
由圖8-A 可知,突變株的泳動(dòng)能力與野生株無(wú)明顯差異;由圖8-B 可知,ΔagrC的群集運(yùn)動(dòng)能力顯著高于野生株。
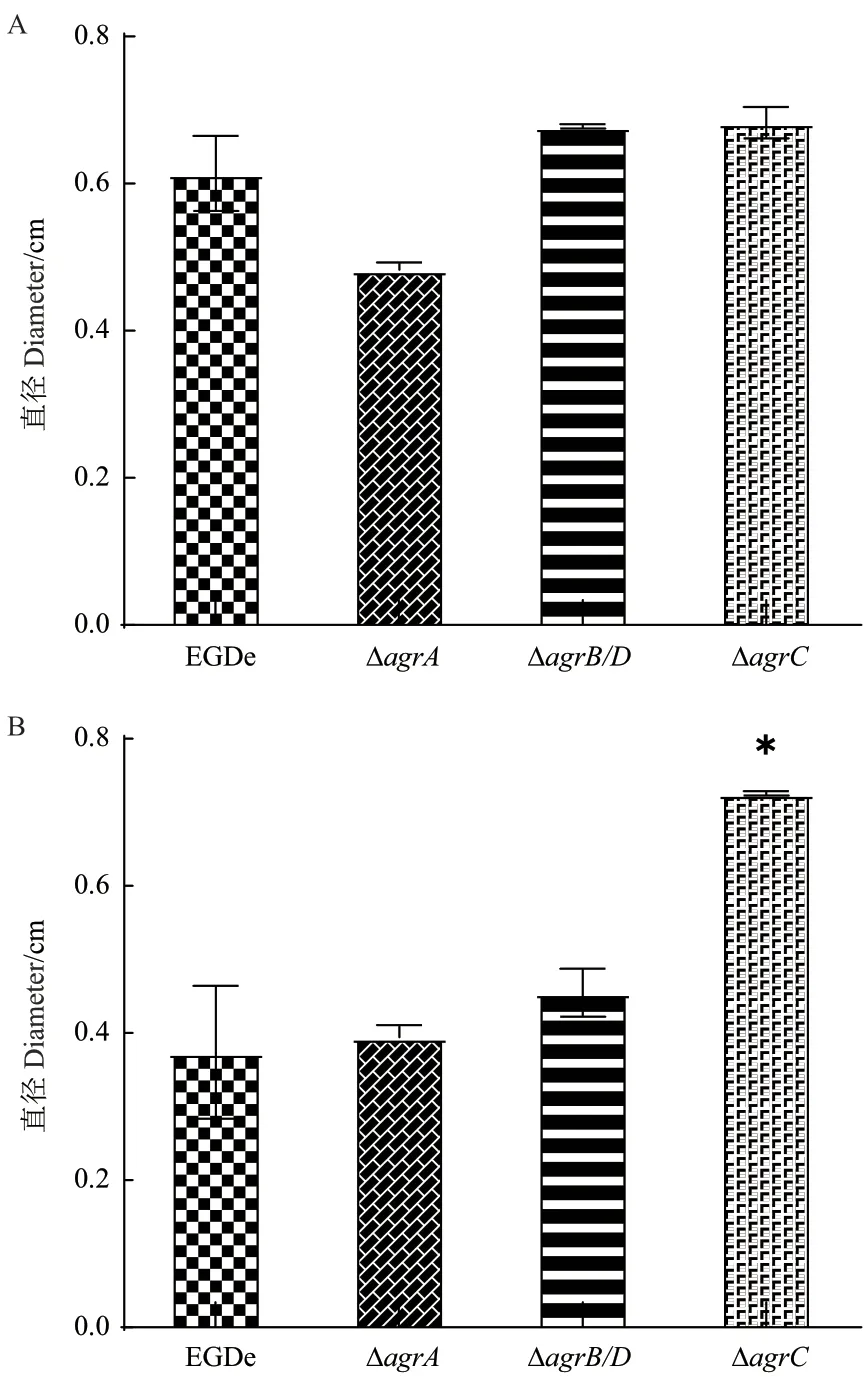
圖8 野生株與agr 基因缺失突變株的泳動(dòng)和群集運(yùn)動(dòng)性Fig. 8 Swimming and swarming of the wild-type strain and agr gene deletion mutants
2.6 運(yùn)動(dòng)相關(guān)基因的轉(zhuǎn)錄水平
由圖9 可知,與野生株相比,motB在ΔagrB/D中的轉(zhuǎn)錄水平顯著下調(diào);flaA在ΔagrA、ΔagrB/D和ΔagrC中的轉(zhuǎn)錄水平均顯著下降,分別為0.38 倍、0.09倍和0.54 倍。
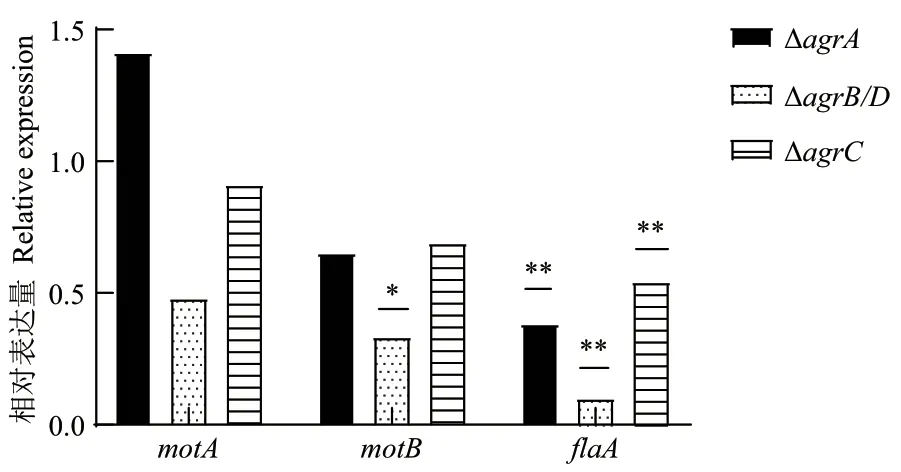
圖9 野生株與agr 基因缺失突變株中運(yùn)動(dòng)相關(guān)基因的轉(zhuǎn)錄水平Fig. 9 Transcription levels of motility-associated genes in the wild type and agr gene deletion mutants
3 討論
除了頭孢噻肟和環(huán)丙沙星,3 株突變株對(duì)其余抗菌藥物的MIC 值均與野生株相同。考慮到微量肉湯法的靈敏度可能不足以反映突變株和野生株的耐藥表型差異,本研究又調(diào)查了這些菌株在亞抑菌濃度抗菌藥物作用下的生長(zhǎng)曲線。結(jié)果顯示,野生株與突變株僅在亞抑菌濃度BC 脅迫下的生長(zhǎng)出現(xiàn)明顯差異。BC 屬于季銨鹽類化合物,是一種高效廣譜的消毒劑,主要用于食品加工環(huán)境、食品生產(chǎn)用具設(shè)備、食品貯藏環(huán)境及工業(yè)水的消毒殺菌[12-14]。生長(zhǎng)曲線結(jié)果表明,當(dāng)BC 的濃度為1 μg/mL 時(shí),ΔagrC的生長(zhǎng)完全受到抑制,ΔagrB/D的生長(zhǎng)延遲期明顯變長(zhǎng),表明agrB/D和agrC與Lm 對(duì)BC 耐藥有關(guān),并且agrC在Lm 對(duì)BC 耐藥中發(fā)揮更大的作用。缺失agrB/D后agrCA的存在仍能保證菌株在亞抑菌濃度BC 下生長(zhǎng),據(jù)此推測(cè)組氨酸蛋白激酶AgrC 可能會(huì)感應(yīng)到胞外BC 的存在,并通過(guò)磷酸化反應(yīng)將信號(hào)傳遞至胞內(nèi);而當(dāng)agrC缺失后,AIP 無(wú)法通過(guò)與AgrC 結(jié)合激活agr系統(tǒng),同時(shí)細(xì)胞也無(wú)法感應(yīng)胞外BC 的存在,因此ΔagrC在亞抑菌濃度BC 下無(wú)法生長(zhǎng)。相反地,agrA的缺失促進(jìn)了Lm 在BC 脅迫下的生長(zhǎng),這可能是由于在ΔagrA中AgrC 與其他的反應(yīng)調(diào)控子發(fā)生信號(hào)串聯(lián)所致。本研究還調(diào)查了Lm在致死濃度BC 作用下的生長(zhǎng)。致死曲線結(jié)果表明,3 株突變株在不同時(shí)間點(diǎn)的存活率均低于野生株。以上結(jié)果表明,agr系統(tǒng)參與Lm 對(duì)BC 耐藥。需要指出的是,本研究利用框內(nèi)敲除構(gòu)建了agr突變體,但是沒(méi)有構(gòu)建相應(yīng)的回復(fù)突變株。在后續(xù)研究中將會(huì)構(gòu)建回復(fù)突變株以排除基因敲除的極性效應(yīng)。
生物被膜是細(xì)菌定植在固體表面的一種常見(jiàn)狀態(tài)[15]。生物被膜的形成給細(xì)菌在不利環(huán)境中的生長(zhǎng)提供了保護(hù),導(dǎo)致消毒劑等抗菌物質(zhì)無(wú)法殺滅被膜內(nèi)部的細(xì)菌,給食品加工造成安全隱患[16-18]。溫度是影響細(xì)菌生物被膜形成的重要環(huán)境因素,因此本研究調(diào)查了Lm 野生株和突變株在不同溫度下的生物被膜形成能力。37、20 和4℃分別代表人體內(nèi)的溫度、食品加工環(huán)境的溫度和食品冷藏的溫度。在37℃條件下,ΔagrC的生物被膜量在培養(yǎng)1 和2 d時(shí)與野生株相比沒(méi)有差異,而到第3 d 后顯著下降,表明agrC可能在Lm 生物被膜形成后期發(fā)揮作用。在20℃條件下,3 株突變株與野生株生物被膜形成能力的差異僅在培養(yǎng)前期較為明顯。在37℃和4℃條件下,與野生株相比,突變株的生物被膜形成能力降低。總體來(lái)說(shuō),agr系統(tǒng)參與Lm 在不同溫度條件下的生物被膜形成。
在生物被膜形成的初期,浮游細(xì)菌可在自身鞭毛的作用下接觸到物體表面,鞭毛介導(dǎo)的運(yùn)動(dòng)性可以使菌體聚集,促進(jìn)生物被膜的形成[19]。由鞭毛介導(dǎo)的泳動(dòng)運(yùn)動(dòng)性與群集運(yùn)動(dòng)性與細(xì)菌生物被膜的形成密切相關(guān)。泳動(dòng)運(yùn)動(dòng)性是細(xì)菌個(gè)體而非合作性的運(yùn)動(dòng)行為,而群集運(yùn)動(dòng)性則是一種群體運(yùn)動(dòng)行為。運(yùn)動(dòng)性增強(qiáng)可能會(huì)促進(jìn)細(xì)菌生物被膜形成,但兩者之間并沒(méi)有絕對(duì)的正相關(guān)關(guān)系[20-21]。本研究結(jié)果表明agr系統(tǒng)與Lm 泳動(dòng)運(yùn)動(dòng)性無(wú)關(guān)。缺失agrC后,菌株的群集運(yùn)動(dòng)能力顯著性增強(qiáng),而運(yùn)動(dòng)相關(guān)基因motB和flaA在?agrC中的轉(zhuǎn)錄水平卻顯著下調(diào),這表明agrC的缺失可能導(dǎo)致其他運(yùn)動(dòng)性相關(guān)基因的轉(zhuǎn)錄水平升高,從而增強(qiáng)菌株的群集運(yùn)動(dòng)性。
Lm 是一種重要的食源性致病菌,隨著殺生物劑的廣泛使用,在食品及環(huán)境中分離出大量的耐受菌株,耐受菌株的出現(xiàn)給人類帶來(lái)極大的安全挑戰(zhàn)。QS 是細(xì)菌細(xì)胞間的通訊系統(tǒng),也是細(xì)胞調(diào)控網(wǎng)絡(luò)中重要系統(tǒng)。本研究的結(jié)果顯示,與野生株相比,缺失agr系統(tǒng)導(dǎo)致Lm 對(duì)頭孢噻肟、環(huán)丙沙星及BC 的敏感性增強(qiáng);突變株生物被膜形成能力下降;缺失agr系統(tǒng)對(duì)Lm 的運(yùn)動(dòng)性沒(méi)有顯著影響。以上結(jié)果表明,agr系統(tǒng)在Lm 對(duì)抗菌藥物耐藥及生物被膜形成中發(fā)揮作用。
4 結(jié)論
本研究利用同源重組技術(shù)構(gòu)建了agrA、agrB/D、agrC基因缺失突變株。與野生株相比,突變株對(duì)頭孢噻肟、環(huán)丙沙星及BC 的耐藥性降低;突變株在不同溫度下的生物被膜形成能力降低。這些結(jié)果表明agr系統(tǒng)參與Lm 對(duì)抗菌藥物的耐藥以及生物被膜的形成。
猜你喜歡
天天愛(ài)科學(xué)(2022年9期)2022-09-15 01:12:54
天天愛(ài)科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
小讀者(2021年2期)2021-03-29 05:03:48
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04