利用胞嘧啶堿基編輯技術(shù)創(chuàng)制耐草甘膦水稻
2023-03-07 12:54:48盧振萬李雪琪黃金光周煥斌
生物技術(shù)通報 2023年2期
盧振萬 李雪琪, 黃金光 周煥斌
(1. 中國農(nóng)業(yè)科學院植物保護研究所 植物病蟲害生物學國家重點實驗室,北京 100093;2. 農(nóng)業(yè)農(nóng)村部桂林作物有害生物科學觀測實驗站,桂林 541399;3. 青島農(nóng)業(yè)大學植物醫(yī)學學院,青島 266109)
水稻(Oryza sativaL.)作為主要的糧食作物之一,全球一半以上人口以水稻為主食。因此,保障和提高水稻產(chǎn)量和品質(zhì)將對緩解糧食壓力和保障糧食安全具有十分重要的意義。隨著耕地和農(nóng)業(yè)勞動力的不斷減少以及水資源日益匱乏[1],水稻的生產(chǎn)栽培方式正逐漸從傳統(tǒng)的移栽向直播轉(zhuǎn)變和發(fā)展[2]。然而,在直播水稻生產(chǎn)過程中,雜草危害變得日趨頻繁和復雜,嚴重影響水稻的產(chǎn)量和品質(zhì)[3]。據(jù)統(tǒng)計,在直播系統(tǒng)中雜草危害造成的水稻產(chǎn)量損失超過80%[4-5]。在傳統(tǒng)農(nóng)業(yè)中,雜草管理主要依靠人工除草和水管理,費時費力。與人工除草相比,利用除草劑需要更少的勞動力和資金,而且效率更高,這使得它成為水稻種植中最廣泛采用的方法[6-7]。
草甘膦是一種廣譜滅生性除草劑,高效且低殘留,雜草不易對其產(chǎn)生抗性,成本較低且易于在環(huán)境中降解[8],這些因素使草甘膦在農(nóng)業(yè)生產(chǎn)中成為使用最廣泛的除草劑。草甘膦作用于植物體內(nèi)莽草酸途徑的5-烯醇丙酮酰莽草酸-3-磷酸合酶(EPSPS),干擾EPSPS 正常酶活功能,阻礙多種蛋白質(zhì)的正常合成而影響植物的生長發(fā)育,導致死亡。由于莽草酸途徑目前只發(fā)現(xiàn)于植物和微生物中,人和哺乳動物中尚未發(fā)現(xiàn)該途徑,因此相對來說,草甘膦對人和哺乳動物具有一定的安全性[9-12]。由于草甘膦對水稻同樣具有毒性,在水稻生長期不能直接施用草甘膦除草,所以培育耐草甘膦水稻對草甘膦用于稻田雜草防治具有重要應用價值。
抗草甘膦作物培育也一直是轉(zhuǎn)基因育種領(lǐng)域最主要的方向。在實驗室內(nèi),目前已培育獲得大量的抗草甘膦轉(zhuǎn)基因水稻,但是由于公眾對轉(zhuǎn)基因作物的安全顧慮而推廣不易。近年來新興發(fā)展起來的基因組編輯技術(shù)則為培育新型水稻抗除草劑材料提供了一條新的思路。借鑒雜草中因除草劑靶標基因發(fā)生點突變從而產(chǎn)生抗藥性的原理,研究人員利用堿基編輯技術(shù)修飾改造作物中的除草劑靶標基因,獲得了無轉(zhuǎn)基因成分的除草劑抗性作物新種質(zhì)。堿基編輯技術(shù)的原理主要為:由切口酶Cas9n(即Cas9(D10A))和堿基脫氨酶組成的堿基編輯器,在sgRNA 引導下結(jié)合在基因組靶位點,此時堿基脫氨酶可將編輯活性窗口內(nèi)的靶堿基進行脫氨形成新嘌呤或嘧啶[13-14]。目前植物中已建立了兩類堿基編輯技術(shù),分別是實現(xiàn)堿基對C:G 向T:A 轉(zhuǎn)換的胞嘧啶堿基編輯技術(shù)、堿基對T:A 向C:G 轉(zhuǎn)換的腺嘌呤堿基編輯技術(shù)[15-16]。研究人員通過利用胞嘧啶堿基編輯技術(shù)對水稻、小麥和西瓜等作物內(nèi)源的ALS 抑制劑類除草劑靶標基因ALS進行編輯,成功獲得了具有雙草醚抗性的水稻OsALS(P171F)、煙嘧磺隆抗性的小麥TaALS(P194F)和苯磺隆抗性的西瓜ClALS(P190S)等材料[17-19];此外,利用腺嘌呤堿基編輯技術(shù)分別對水稻內(nèi)源OsACCase和OsTub2A進行編輯,從而分別獲得具有氟吡甲禾靈抗性和二硝基苯胺類除草劑抗性的水稻[20-21]。
在持續(xù)的草甘膦選擇壓力下,部分雜草也開始出現(xiàn)草甘膦抗性,比如抗草甘膦牛筋草Eleusine indica的EiEPSPS 的第106 位脯氨酸P 突變?yōu)榻z氨酸S(即P106S)可使牛筋草具有草甘膦抗性[22]。因此本研究通過對水稻OsEPSPS 氨基酸序列進行分析和預測可能的草甘膦抗性突變位點,并利用堿基編輯技術(shù)對該位點進行堿基編輯,創(chuàng)制耐草甘膦水稻新種質(zhì),為水稻耐除草劑品種培育和稻田的雜草防治奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 植物材料 水稻粳稻(Oryza sativaL.ssp.geng)早熟模式品種Kitaake。
1.1.2 實驗菌株及載體見表1。

表1 載體與菌株Table 1 Bacterial strains and plasmids
1.1.3 所用農(nóng)藥及引物 植株噴施用的草甘膦異丙胺鹽水劑(速力達,有效成分30%)購自河北中保綠農(nóng)作物科技有限公司,載藥平板使用的95%草甘膦原藥購自上海翊圣生物科技有限公司。
1.2 方法
1.2.1 序列比對分析 從水稻基因組數(shù)據(jù)庫網(wǎng)站(http://rice.uga.edu/index.shtml) 下 載OsEPSPS的DNA 及其編碼的氨基酸序列,同時在NCBI 下載抗草甘膦的牛筋草EiEPSPS-R(P106S)氨基酸序列[22],利用DNAMAN 軟件對上述序列進行序列比對分析以期獲得OsEPSPS 中相對的氨基酸位點。
1.2.2 sgRNA 的設計和載體構(gòu)建 為實現(xiàn)對OsEPSPS 第177 位脯氨酸P 密碼子CCA 的堿基編輯,選擇的靶標序列應使得該密碼子的C 處于2-8 位(靶標序列遠離PAM 序列的一端為1)的編輯窗口內(nèi),同時需要滿足現(xiàn)有堿基編輯器的識別PAM 要求。最后參考Zhou 等[23]方法,通過在靶標序列的正負鏈加上BsaI 酶切黏性末端,從而獲得sgRNA 引物gOsEPSPS-F1/R1,該引物經(jīng)磷酸化和退火形成雙鏈DNA 后,酶切插入到中間載體pENTR4:sgRNA5,獲得重組載體g5:gOsEPSPS-F1/R1,再利用gateway 重組反應將sgRNA 表達盒整合到pUbi:rBE22,獲得最終的堿基編輯載體pUbi:rBE22-gOsEPSPS-F1/R1。
1.2.3 水稻的遺傳轉(zhuǎn)化 將載體pUbi:rBE22-gOsEPSPS-F1/R1 經(jīng)電擊轉(zhuǎn)入到根癌農(nóng)桿菌EHA105感受態(tài)細胞中,參考Hiei 等[24]方法進行農(nóng)桿菌介導的水稻遺傳轉(zhuǎn)化,獲得T0代轉(zhuǎn)基因水稻。
1.2.4 堿基編輯植株的檢測 采用十六烷基三甲基溴化銨(CTAB)方法提取水稻材料的基因組DNA,利用OsEPSPS基因的特異引物OsEPSPS-F2/R2 進行PCR 擴增和Sanger 測序,鑒定靶位點是否發(fā)生堿基編輯。在T1代水稻植株中,參照表2 中的引物對外源轉(zhuǎn)基因元件上的Cas9、潮霉素抗性基因Hyg 和sgRNA 表達盒進行PCR 檢測。
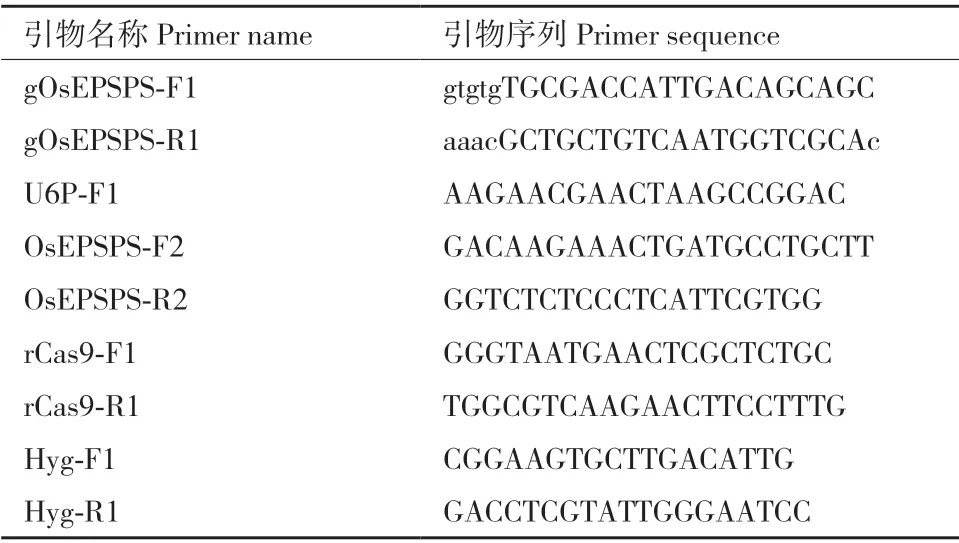
表2 引物信息Table 2 Primer information
1.2.5 堿基編輯材料的草甘膦抗性檢測 (1)載藥培養(yǎng)基檢測:將堿基編輯材料和Kitaake 種子去殼經(jīng)50%的84 消毒液表面消毒處理后,轉(zhuǎn)移到含不同濃度(0 mg/L、2 mg/L、4 mg/L、8 mg/L、12 mg/L)草甘膦的1/2MS 固體培養(yǎng)基上,置于25-28℃、24 h光照下培養(yǎng)。14 d 后觀察種子的萌發(fā)生長情況。
(2)草甘膦噴施檢測:采用噴霧塔(壓力為0.3 MPa),對2-3 葉期水稻幼苗噴施草甘膦異丙胺鹽水劑,草甘膦異丙胺鹽的田間推薦劑量為3 000-6 000 mL/hm2,本研究選取4 500 mL/hm2(即1 350 g a.i./ha)作為田間推薦劑量,按田間推薦劑量的4 倍進行噴施,于噴施后14 d 觀察水稻植株的生長情況。
1.2.6 突變體植株的經(jīng)濟性狀調(diào)查 將堿基編輯材料和Kitaake 種植在玻璃溫室中,待其成熟后,測量土面至最高穗頂(不連芒)的平均高度,即為株高。在培養(yǎng)皿內(nèi)鋪2 層濾紙,加入適量的水,然后均勻的擺上100 粒種子,每天觀察1 次,5 d 后統(tǒng)計正常幼苗數(shù)。種子發(fā)芽率(%)=發(fā)芽的種子數(shù)/供檢測的種子數(shù)×100%。
2 結(jié)果
2.1 堿基編輯靶點的選擇
在牛筋草中已發(fā)現(xiàn)并報道其EiEPSPS 的第106位脯氨酸P 突變?yōu)榻z氨酸S(即P106S)可使雜草對草甘膦產(chǎn)生抗性。那么利用堿基編輯技術(shù)將水稻的內(nèi)源OsEPSPS 蛋白進行類似的突變,是否會使水稻產(chǎn)生草甘膦抗性呢?對此,我們對從水稻基因組數(shù)據(jù)庫網(wǎng)站(http://rice.uga.edu/index.shtml)下載的OsEPSPS(登錄號為LOC_Os06g04280.1)氨基酸序列與牛筋草的EiEPSPS 氨基酸序列進行比對分析,發(fā)現(xiàn)牛筋草中已報道的抗性變異位點P106 對應水稻OsEPSPS 的P177 位點(圖1)。

圖1 水稻OsEPSPS 和牛筋草EiEPSPS 的氨基酸序列比對Fig. 1 Amino acid sequence alignment of rice OsEPSPS and Eleusine indica EiEPSPS
水稻OsEPSPS 的P177 位點密碼子為CCA,通過胞嘧啶堿基編輯(C>T),可實現(xiàn)P177S(CCA>TCA)或P177L(CCA>TTA)。根 據(jù)CRISPR/Cas9 系 統(tǒng) 介導堿基編輯器sgRNA 設計原則,即靶堿基位于堿基編輯活性窗口內(nèi)和靶點附近具有合適PAM 序列,結(jié)合水稻OsEPSPS 的P177 位點密碼子附近的DNA序列,我們選擇NGT PAM,以“ATGCGACCATTGACAGCAGCCGT”(CCA 為P177 的 密 碼 子,CGT為PAM 序列)為靶標序列,利用識別NG PAM 的rBE22 對其進行靶堿基定向替換(圖2-A)。利用網(wǎng)站“http://www.rgenome.net/cas-offinder/”與http://skl.scau.edu.cn/offtarget/對該靶標序列的潛在脫靶位點進行預測,同時在水稻基因組數(shù)據(jù)庫中進行Blast 比對,結(jié)果均顯示未獲得和靶標序列具有0-2 個堿基錯配的潛在脫靶位點,表明該靶標序列在水稻基因組中具有很好的特異性。
2.2 堿基編輯材料的獲得
根據(jù)上述靶標序列合成sgRNA 引物對gOsEPSPS-F1 和gOsEPSPS-R1,并經(jīng)分子克隆獲得堿基編輯載體pUbi:rBE22-gOsEPSPS-1,經(jīng)農(nóng)桿菌介導的遺傳轉(zhuǎn)化獲得T0代轉(zhuǎn)基因水稻植株。提取T0代水稻植株的基因組DNA 并利用OsEPSPS-F2/R2 引物對靶基因進行PCR 擴增,PCR 產(chǎn)物的Sanger 測序結(jié)果表明在76 株T0代水稻中,有4 株發(fā)生了C 向T 的突變,均為雜合突變,編輯效率為5.26%。其中植株#96 在靶標序列(遠離PAM 序列為1)的第4位C 突變?yōu)門,使得第176 位精氨酸(Arginine,R)密碼子CGA 突變?yōu)榻K止密碼子TGA;植株#9 和#97 在靶標序列的第4、7 和8 位均發(fā)生C 向T 的替換,使得R176 密碼子CGA 突變?yōu)榻K止密碼子TGA和P177 密碼子CCA 突變?yōu)門TA(編碼亮氨酸L);只有植株#105 僅在靶標序列的第7 和8 位發(fā)生的C向T 替換實現(xiàn)了P177L 突變,且未造成提前終止翻譯(圖2-B)。因此,選擇植株#105 進行后續(xù)的研究工作。
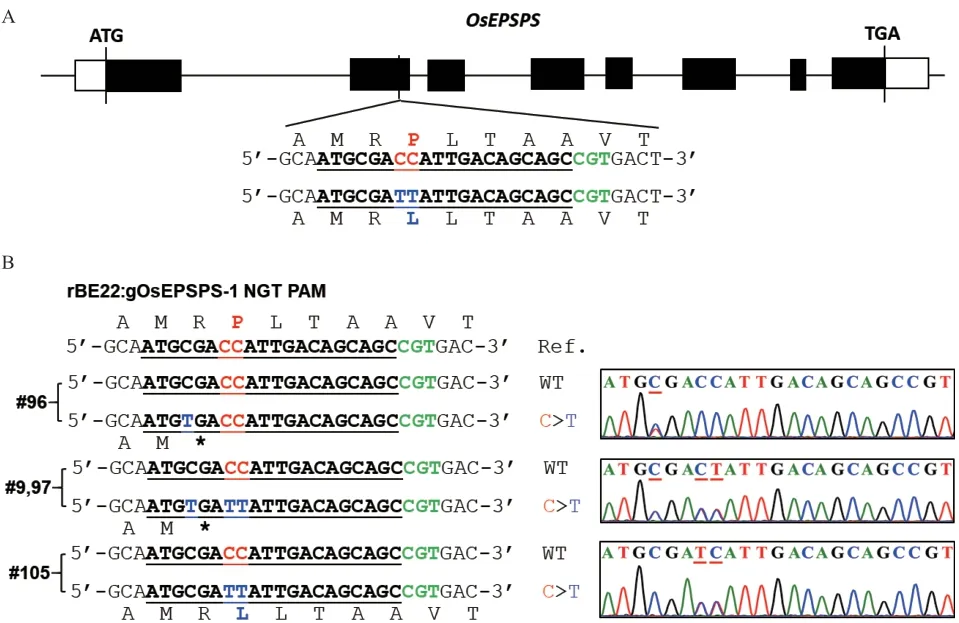
圖2 T0 代堿基編輯材料的鑒定Fig. 2 Identification of base editing materials in T0
為了獲得堿基編輯位點純合突變且去轉(zhuǎn)基因成分的基因編輯水稻材料OsEPSPS(P177L),我們對T0代植株#105 的自交后代植株(即T1代)進行單株提取基因組DNA,分別用胞嘧啶堿基編輯器擴增引物rCas9-F/R、潮霉素B 抗性基因擴增引物Hyg 和sgRNA 表達框特異擴增引物U6p-F1/gOsEPSPS-R1 進行PCR 擴增,檢測各T1代植株是否含有外源轉(zhuǎn)基因元件。檢測結(jié)果表明,32 株T1代植株中有6 株未檢測到外源轉(zhuǎn)基因元件的存在(圖3-A)。隨即對該6 株進行靶位點測序檢測,成功拿到了純合突變體(圖3-B)。
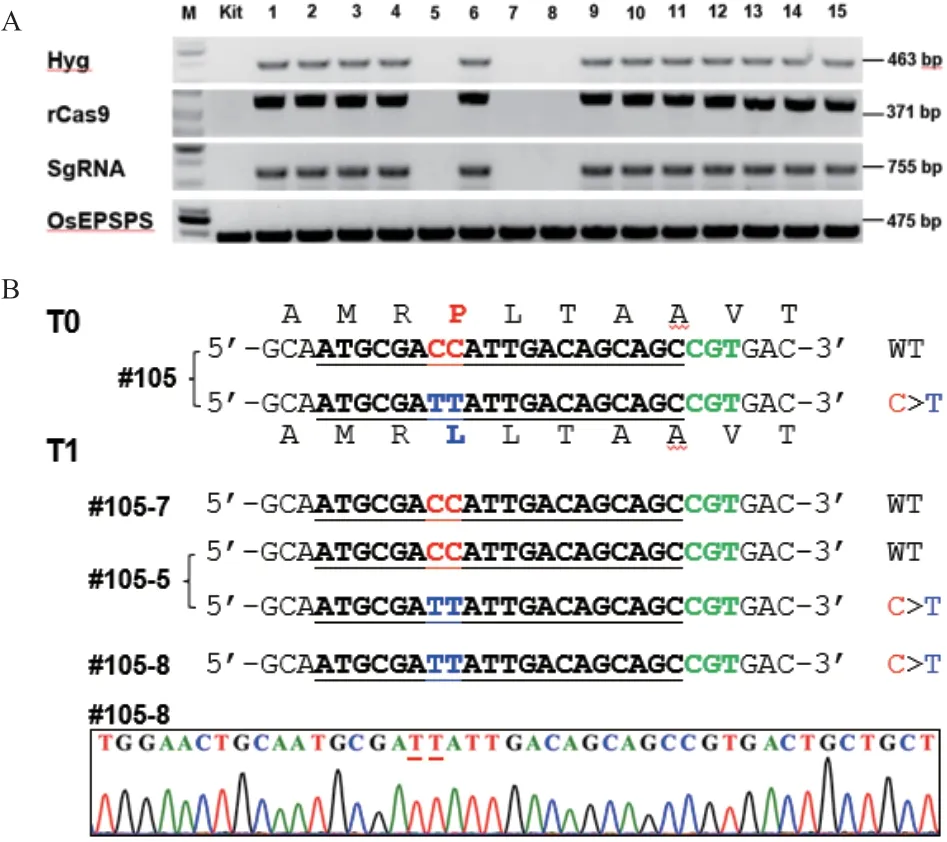
圖3 無外源轉(zhuǎn)基因成分且純合突變的堿基編輯材料OsEPSPS(P177L)鑒定Fig. 3 Identification of base editing materials OsEPSPS(P177L)without exogenous components and homozygous mutations
至此,我們獲得了水稻內(nèi)源草甘膦靶標蛋白基因OsEPSPS的無外源轉(zhuǎn)基因成分且純合突變的堿基編輯材料OsEPSPS(P177L)。
2.3 堿基編輯材料OsEPSPS(P177L)對草甘膦的抗性檢測
為驗證水稻堿基編輯材料OsEPSPS(P177L)對草甘膦的抗性,首先利用載藥平板檢測分析了其種子在草甘膦處理下的萌發(fā)生長情況。如圖4-A 所示,未添加草甘膦時,OsEPSPS(P177L)和Kitaake的生長情況一致,沒有明顯的差異;但隨著培養(yǎng)基中草甘膦濃度的增加,Kitaake 生長受抑制的程度比OsEPSPS(P177L)更敏感和更嚴重。當草甘膦濃度達到8 mg/L 時,野生型Kitaake 已完全不能正常發(fā)芽且種子全部變黑死亡,此時OsEPSPS(P177L)生長受到抑制的程度仍較輕,當草甘膦濃度升到12 mg/L 時,OsEPSPS(P177L)幼苗表現(xiàn)出生長受到抑制。

圖4 堿基編輯材料OsEPSPS(P177L)對草甘膦的抗性檢測Fig. 4 Glyphosate-tolerance assay of base editing materials OsEPSPS(P177L)plants
為進一步模擬大田間草甘膦的施用情況,我們采用噴霧塔對野生型Kitaake 和OsEPSPS(P177L)水稻植株進行噴施處理,以大田推薦劑量為4 500 mL/hm2設定其濃度為1×(1 倍)。噴施后14 d,在4 倍(5 400 g a.i./ha)的大田推薦劑量情況下,Kitaake 全部枯萎死亡,而OsEPSPS(P177L)在該倍數(shù)的噴施情況下依然存活。以上結(jié)果表明,水稻EPSPS基因中第177 位脯氨酸突變?yōu)榱涟彼崮軌蛸x予水稻一定程度的草甘膦抗性。
2.4 突變體植株經(jīng)濟性狀調(diào)查
獲得的優(yōu)良性狀植株投入應用的基礎(chǔ)是不能影響其經(jīng)濟性狀,為驗證本研究獲得的OsEPSPS(P177L)材料的氨基酸突變是否會影響經(jīng)濟性狀,我們選擇了株高和發(fā)芽率作為評估指標(圖5)。
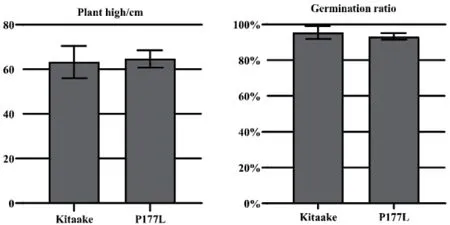
圖5 P177L 純合突變體經(jīng)濟性狀測定Fig. 5 Assay of P177L homozygous mutants’ economic traits
將Kitaake 和OsEPSPS(P177L)材料的熟種子經(jīng)過前期催芽后,同時種植在溫室中直至成熟。整個生長階段二者的株高沒有明顯差異,Kitaake 的平均株高為63.27 cm,OsEPSPS(P177L)的平均株高為64.68 cm(P>0.05)。Kitaake 和P177L 純合突變體的成熟種子在土壤基質(zhì)中的發(fā)芽率分別為95.56%和93.33%(P>0.05)。上述分析結(jié)果表明,水稻EPSPS基因中Pro177 被Leu 替代后不會對經(jīng)濟性狀產(chǎn)生不良影響。
3 討論
雜草能夠?qū)λ镜纳L產(chǎn)生不利影響,雜草不僅爭奪水稻的水分和養(yǎng)分,而且還是傳播病原菌的重要媒介,因此消滅雜草無疑是水稻種植過程中的重要田間管理措施。盡管除草劑的使用極大的提高了勞動效率,但是由于草甘膦對水稻的毒性作用,導致其不能直接應用于水稻田中。因此將草甘膦應用于稻田中的雜草防治一直以來是一個難題。而通過創(chuàng)制耐草甘膦的水稻種質(zhì)資源,是經(jīng)濟有效的解決方式。
目前創(chuàng)制抗除草劑作物主要有以下3 個主要途徑:修飾除草劑靶蛋白以抑制其對除草劑的親和力;提高除草劑靶蛋白的表達量以維持其功能;引入或增加除草劑解毒蛋白,保護靶蛋白免受危害。而第一種方法無疑是直接有效的方法,也是創(chuàng)制抗草甘膦水稻中應用最為廣泛的途徑,然而這種方法并不能阻止草甘膦在植物體內(nèi)的轉(zhuǎn)運和積累。目前,創(chuàng)制抗草甘膦水稻的研究中大多數(shù)利用的是引入微生物的外源基因,例如I.variabilis-EPSPS*[25],然而其還沒有商業(yè)化推廣的條件。
本研究通過堿基編輯技術(shù)直接對水稻內(nèi)源基因OsEPSPS進行精準編輯,從而獲得了耐草甘膦的水稻材料,這種方法不僅快速安全,而且減少了引入外源基因所帶來的擔憂,為抗除草劑作物邁上商業(yè)化創(chuàng)造了條件。
4 結(jié)論
本研究利用胞嘧啶堿基編輯系統(tǒng)rBE22 對水稻內(nèi)源草甘膦靶標基因OsEPSPS進行堿基定向替換,成功創(chuàng)制獲得了對草甘膦具有耐受性且經(jīng)濟性狀不受影響的水稻新種質(zhì)OsEPSPS(P177L)。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
今日農(nóng)業(yè)(2019年15期)2019-01-03 12:11:33
現(xiàn)代園藝(2017年19期)2018-01-19 02:50:21
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:36
營銷界(2015年23期)2015-02-28 22:06:18