
無瓣海桑根響應鹽脅迫的轉錄組分析
2023-04-29 00:44:03梁銳濤韓維棟楊少瑕等
林業科學研究 2023年1期
梁銳濤 韓維棟 楊少瑕等
關鍵詞:無瓣海桑;鹽脅迫;轉錄組;差異表達基因
中圖分類號:S728.5 文獻標志碼:A 文章編號:1001-1498(2023)01-0068-11
土壤鹽漬化現象幾乎遍布所有的氣候區域以及地理環境,是影響農業和林業生產的主要問題。土壤鹽漬化使植物受到滲透脅迫、離子毒害、膜透性改變及生理代謝紊亂等危害,嚴重制約了植物的生長,是在植物生產與栽培上一個非常嚴峻的問題。因此,通過對植物抗鹽的分子調控機制進行探析,挖掘耐鹽相關基因,并應用于新種質創制和新品種選育,對于緩解土壤鹽漬化問題具有重要的現實意義。
長期生長在濱海區域潮間帶的紅樹林,由于其特殊的、有別于陸地和淡水植物的生長環境,演化出了一套特殊的鹽逆境適應機制。植物的耐鹽機制涉及滲透物質積累、蛋白積累、抗氧化酶的誘導、植物激素的誘導和光合作用路徑的變化5個方面。對于紅樹植物,不同的紅樹物種具體的耐鹽機制有所不同。無瓣海桑(Sonneratia apetala Buch.-Ham.)是海桑科海桑屬紅樹喬木,具有生長快、耐淹浸、耐貧瘠、耐寒強等特點,是濱海灘涂的先鋒造林樹種,在維持海岸帶生態平衡方面起著重要作用。先前有關其耐鹽特性的研究主要集中在形態、生理生態層面,如廖巖等研究了不同鹽度處理3個月下無瓣海桑根、莖、葉中的蛋白質、可溶性糖、丙二醛含量、抗氧化酶活性的變化情況,并探討了它們的相互關系,為耐鹽樹種的選育提供了參考。然而,目前鮮見關于無瓣海桑耐鹽分子機制方面的研究報道。
轉錄組測序是轉錄組學研究的核心技術,被廣泛應用于新基因的挖掘等。目前,最新的測序技術已經發展到了第三代。三代測序技術具有單分子、超長讀長、無需PCR擴增等優點,已在基因組學、轉錄組學、表觀遺傳學等領域上得到了應用。在轉錄組學上,三代全長轉錄組測序(lso-Seq)的應用,可彌補二代測序技術讀段較短、無法覆蓋整個轉錄本的缺點,從而做到從全長轉錄本水平上研究轉錄組。但三代測序技術存在堿基錯誤率高、成本高等缺點,目前應用并不廣泛。二代測序有通量高、成本低的特點,且發展至今已取得了較好的工作基礎,因此,二代和三代測序結合使用成為很多研究者的策略。如梅瑜等利用基于SMRT技術原理的PacBio平臺三代測序技術,對甘葛藤(Pueraria thomsoruu Benth)進行全長轉錄組測序及分析,從轉錄數據中鑒定了黃酮類生物合成基因、轉錄因子、R基因和SSR標記,為甘葛藤的選育和利用提供了基礎。然而,對于紅樹植物,當前研究僅限于對白骨壤(Avicennia marina (Forsk.) Vierh.)、秋茄(Kandeliacandel (Linn.) Druce/Kandelia obovata)、角果木(Ceriops tagal (perr.)C.B.Rob)、竹節樹(Carallia brachiate (Lour.) Merr.)和老鼠簕(Acanthus ilicifolius L.)等個別物種開展了轉錄組學相關研究。特別是,至今對無瓣海桑僅開展了以三代全長轉錄組數據集為基礎的鹽脅迫應答轉錄因子挖掘工作。因此,本研究利用二代與三代結合的高通量測序技術,對鹽脅迫處理前后的無瓣海桑根系進行轉錄組測序分析,從中鑒定并初步篩選出無瓣海桑根系響應鹽脅迫的候選基因,可為深入研究無瓣海桑的耐鹽基因組學、分子生物學和進一步揭示紅樹植物的耐鹽機制提供理論基礎。
1材料與方法
1.1材料與處理
本實驗原材料采自于廣東省湛江市特呈島紅樹林自然保護區(21°09′~21°10′N,110°25′~110°27′E)。2019年10月采集無瓣海桑的種子,播種于人工配制的營養土中。1a后,選取長勢一致的無瓣海桑幼苗,分別設置對照組(RCK)和處理組(RT)。處理組用500 mmol·L-1的NaCl溶液每3d進行1次鹽脅迫處理,對照組使用等量清水。對照組(RCK_a、RCK_b、RCK_c)和處理組(RT_a、RT_b、RT_c)均進行了3次生物學重復。10 d后分別取對照組和處理組根部組織液氮速凍,于-80℃冰箱保存。
1.2方法
1.2.1測序文庫構建及測序 提取6個樣本的RNA,質檢合格后構建三代全長轉錄組測序文庫和二代RNA-seq測序文庫,并進行測序及分析。二代測序文庫構建方法如下:(1)用mRNA富集法或rRNA去除法對total RNA進行處理;(2)用打斷buffer把獲得的RNA片段化,隨機的N6引物進行反轉錄,合成cDNA二鏈形成雙鏈DNA,再將其末端補平并5'端磷酸化;(3)連接一個3'端有凸出“T”的鼓泡狀的接頭;(4)通過特異的引物進行PCR擴增得到PCR產物,將其熱變性成單鏈,再用一段橋式引物將單鏈DNA環化得到單鏈環狀DNA文庫;(5)利用BGISEQ-500平臺測序。
PacBio測序文庫構建方法如下:取6個樣本的RNA,等量混合,使用SMARTerTM PCRcDNA Synthesis試劑盒將混合的RNA樣品反轉錄合成第-鏈cDNA,經PCR擴增合成第二鏈cDNA,然后對雙鏈DNA經過2次PCR擴增后的片段進行損傷修復、末端修復,并連接SMRT適配體后構建全長轉錄組測序文庫,利用PacBio平臺進行測序。
文庫構建及測序工作由華大基因公司完成。
1.2.2測序數據組裝和基因定量 為保證數據質量,需對測序得到的原始數據進行過濾、參考基因比對等前期處理。本研究使用華大自主研發的過濾軟件SOAPnuke對6個樣本進行過濾,去除包含接頭污染、未知堿基N含量大于10%、低質量的reads。使用Bowtie2將clean reads比對到三代全長轉錄組數據庫;利用RSEM計算各個樣品的基因表達水平。樣本間的相關性分析通過皮爾遜相關系數(Pearson correlation coefficient)進行。
1.2.3差異基因的選擇與功能注釋分析 本研究基于負二項分布原理的Deseq2模型分析方法,對2組樣本進行差異檢測分析,并以差異倍數值(fold change, FC)≥2或≤0.5, P-value<0.01,Q-value<0.05作為篩選差異基因的標準,并將該篩選得到的差異基因用于后續的GO和KEGG Pathway富集分析,前者通過agriGO網站(http://systemsbiology.cau.edu.cn/agriGOv2/)進行,后者借助基因數據庫網站(https://www.kegg.jp/)完成。
2結果與分析
2.1測序質量分析
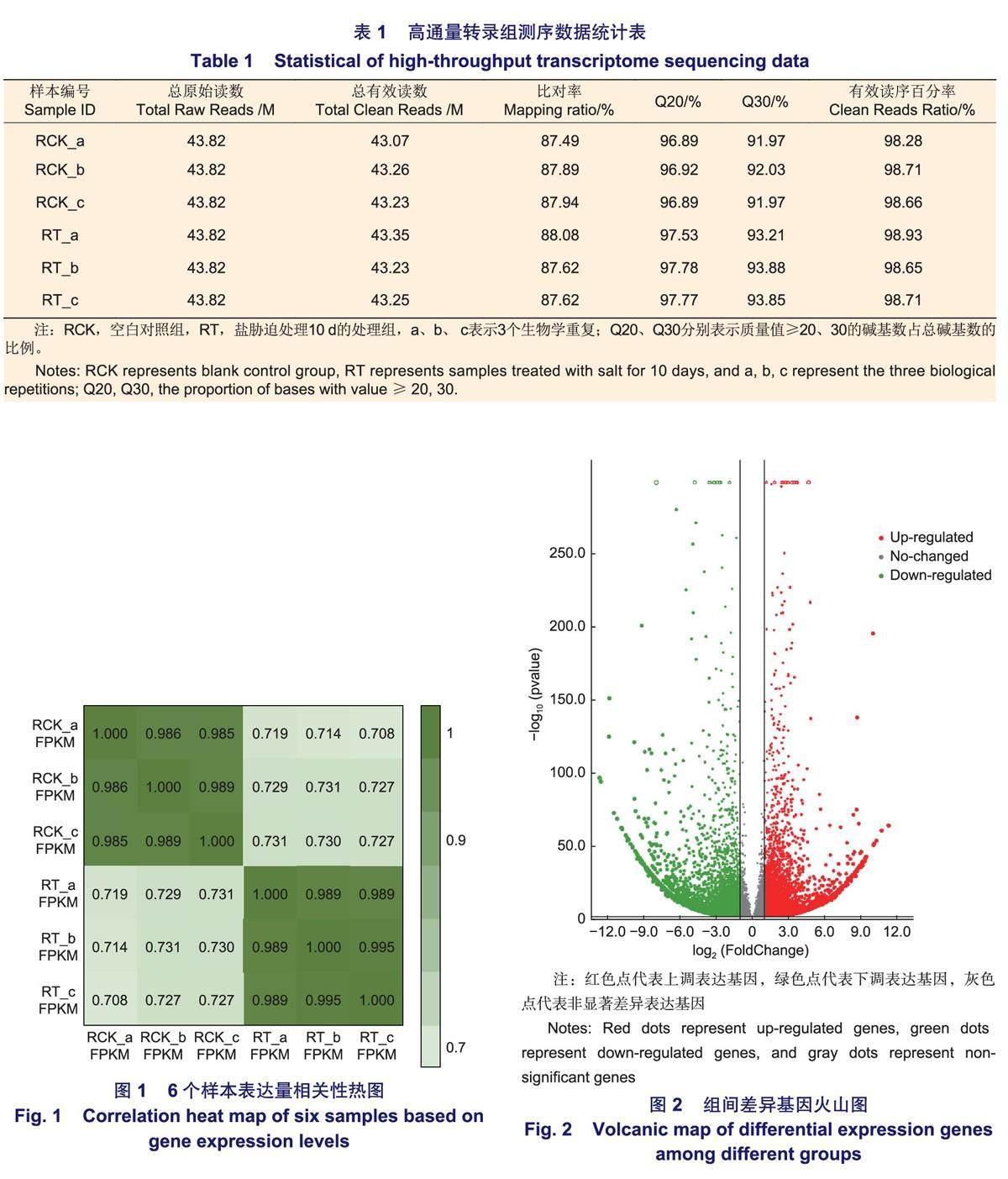
通過SOAPnuke軟件對原始數據進行過濾后,如表1所示,有效讀序百分率均在98%以上,Q20(堿基量≥20%)均大于96%且Q30(堿基量≥30%)均>91%,比對率均在87%以上。6個樣品轉錄本reads覆蓋度的峰值在90~100;空白對照RCK組和鹽處理RT組的三次生物學重復間的相關系數極高(>0.98,圖1)。以上數據均說明6個樣品的測序質量較高,測序數據可靠,可用于后續的差異表達分析。
2.2樣本差異基因表達情況
無瓣海桑根系在500 mmol·L-1的NaCl脅迫10 d后,共檢測到20 289個基因表達量發生了改變(圖2)。2組樣本間共有14 401個基因差異表達,其中,7 153個差異基因上調表達,lOg2FC最大值為11.32(isoform_13 051);7 248個差異基因下調表達,lOg2FC最低值為-12.69(isoform_11 580)。結果表明,下調表達的差異基因數量多于上調的數量。
2.3差異基因的GO分類和顯著性富集分析
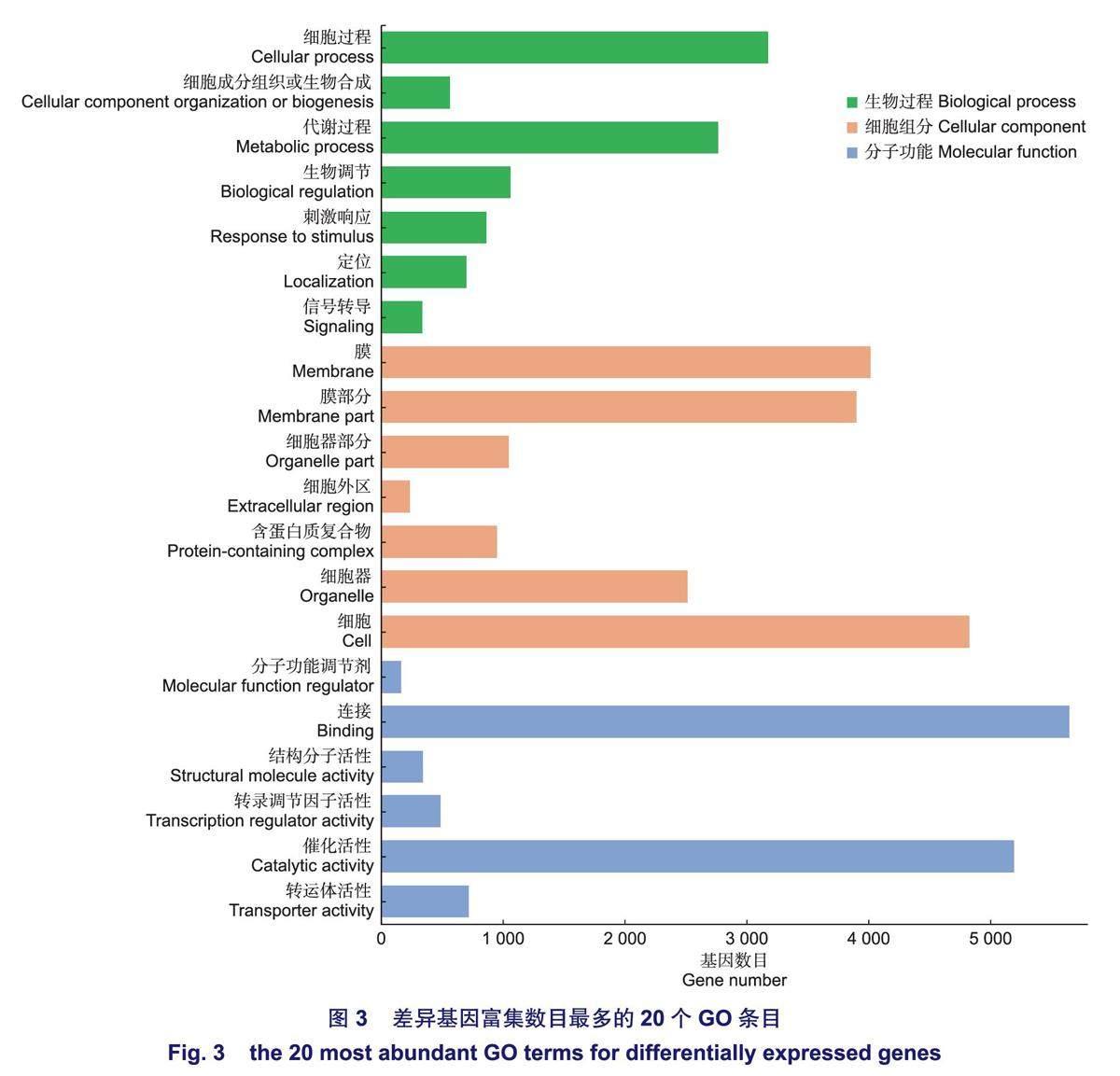
將差異基因比對到Gene Ontology (GO)數據庫,共有11068個差異基因注釋到47條GO條目,注釋率為76.9%,涉及3大主要功能分類:生物過程、細胞組分和分子功能,占比分別為25.02%,43.76%,31.21%,由此可知,大多數差異基因的功能與細胞組分具有較大的相關性。根據GO功能注釋分類結果(圖3),這些差異基因在參與生物過程中,主要集中在細胞過程和代謝過程;在細胞組分中,主要集中在細胞、膜、膜部分;在分子功能中,主要集中在連接、催化活性。表明在高鹽脅迫下,上述的亞類中涉及到的基因可能在無瓣海桑根系中響應強烈。
2.4差異基因的KEGG富集分析
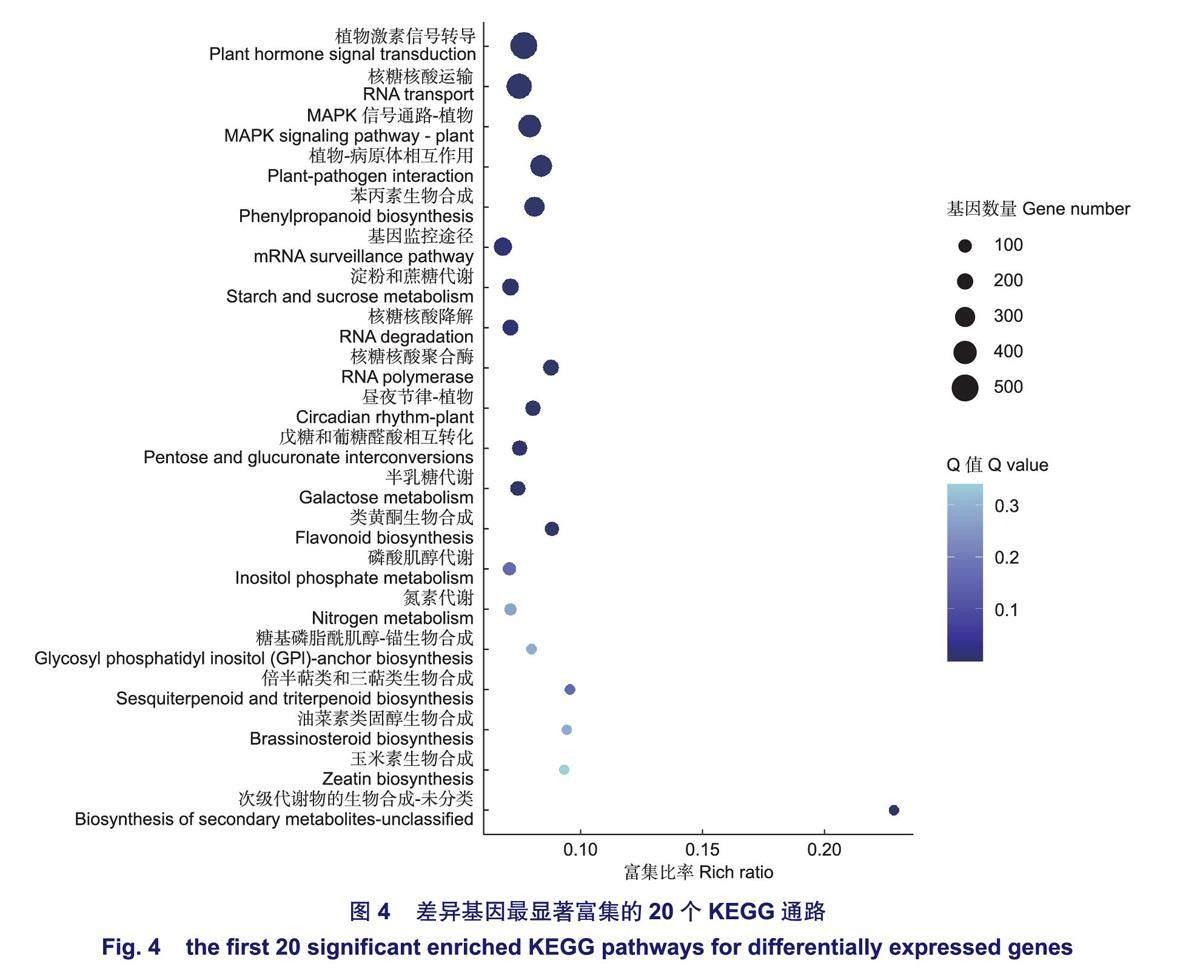
在KEGG分析結果中,共有6189個差異基因富集到134條通路,以Q-value<0.05作為顯著富集標準,共有14條顯著富集的通路(圖4),其中,富集差異基因數量最多的是植物激素信號轉導(ko04075),涉及512個基因,最顯著富集的植物一病原體相互作用(ko04626)涉及367個基因。富集分析結果表明,無瓣海桑根系基因主要通過14條顯著通路參與鹽脅迫的響應,并且其中涉及的基因可能對無瓣海桑根系耐鹽性的調控發揮作用,關注這些顯著通路,有助于對無瓣海桑根系耐鹽候選基因的篩選。
2.5響應鹽脅迫的差異表達基因功能分析
通過進一步對差異基因進行功能注釋分析,本研究共篩選出89個無瓣海桑根系抗鹽候選功能基因,涉及12條KEGG通路(表2,圖5)。
2.5.1抗氧化基因的差異表達 正常情況下,植物體內存在著活性氧(reactive oxygen species,ROS)產生和清除的動態平衡,但外界的不良環境會打破這個平衡,導致ROS的積累,產生毒害。植物體內存在2種ROS清除機制——酶促清除和非酶促清除,前者主要涉及超氧化物歧化酶(SOD)、過氧化氫酶(CAT)等,后者主要涉及抗壞血酸、類黃酮物質等。本研究發現6類共24個活性氧清除基因(圖6),其中,上調表達基因數量最多的是過氧化物酶基因,總數為7,表達量差異變化最大的為銅鋅家族超氧化物歧化酶基因,差異倍數達到6.46。這些上調表達的基因有利于無瓣海桑在鹽脅迫前期,合成多種抗氧化酶類物質,提高體內抗氧化和ROS清除的能力,增強耐受性。
2.5.2滲透調節物質基因的差異表達 植物在對鹽脅迫的適應過程中,細胞會積累一定量的可溶性有機物質,作為滲透調節物質參與滲透調節,以適應外界的低水勢。在本研究中,發現10種共22個編碼生成有機滲透調節物質的基因(圖6),涉及水蘇糖、棉子糖、蔗糖、海藻糖、脯氨酸、甜菜堿、熱激蛋白等滲透調節物質。在這些基因中,部分基因存在上調表達,表明在鹽脅迫下,無瓣海桑可通過合成一些滲透調節物質,以調節鹽漬環境下植物細胞內的滲透勢。
2.5.3植物激素基因的差異表達 植物體內的多種激素在其生長發育、代謝等生理過程中發揮著重要的調控作用。本研究發現7種共19個涉及2種植物激素(IAA與ABA)的基因(圖6),其中,IAA途徑相關的有生長素應答因子(auxin response factor,ARF)、生長素應答GH3家族基因(auxin responsive GH3 gene family, GH3)、SAUR家族蛋白(SAUR family protein,SAUR)和生長素響應蛋白(auxin-responsive protein IAA,IAA)共11個基因;涉及ABA途徑的有脫落酸應答元件結合因子(ABA responsive element bindingfactor,ABF)、脫落酸受體PYR/PYL家族(abscisic acid receptor PYR/PYL family,PYL)和蛋白磷酸酶2C(protein phosphatase 2C,PP2C)共8個基因。IAA相關基因除GH3基因均上調表達外,其他同時存在上下調表達,而ABA相關基因大部分下調表達。上述結果表明,植物激素能夠在無瓣海桑的信號轉導途徑中發揮作用,不同激素共同調控鹽脅迫響應過程。
2.5.4植物蛋白激酶基因的差異表達 蛋白激酶可調控植物的生長,在逆境中起到調控感受脅迫信號、啟動各種非生物逆境響應的作用。本研究發現6種共10個蛋白激酶相關基因(圖6),其中,2種促分裂原活化蛋白激酶基因(mitogen-activated protein kinases,MAPK)MPK4、MPK6和2種絲氨酸,蘇氨酸蛋白激酶基因PBS1、CTR1均上調表達,1種促分裂原活化蛋白激酶基因(mitogen activated protein kinase,MEKK)MEKK1和1種鈣依賴性蛋白激酶基因(calcium-dependent protein kinase, CDPK)中均有2個上調表達,1個下調表達。可見這些蛋白激酶在無瓣海桑根系響應高鹽脅迫時,發揮著尤為重要的調控作用。
2.6響應鹽脅迫的轉錄因子分析
轉錄因子是一類功能蛋白質,具有調控基因表達的作用。當植物受到逆境脅迫時,轉錄因子通過與相應的順式作用元件相互作用,調控并減輕逆境脅迫帶來的傷害,以適應各種逆境。本研究發現8種共14個差異表達的轉錄因子(圖6),涉及5個家族,分別為WRKY家族、MYB家族、GRAS家族、bHLH家族和EREBP家族,其中,WRKY家族發現的成員數量最多,暗示WRKY家族在無瓣海桑響應鹽脅迫中起著重要的調控作用。
3討論
植物對鹽脅迫的響應是一個多基因參與的復雜過程,從微觀上的基因響應到宏觀上生理的變化,經歷了一系列的信號傳遞與調節過程,最終適應鹽逆境。通過轉錄組測序技術對植物鹽脅迫下的轉錄調控機制進行研究分析,能夠快速準確地初步篩選出其中的耐鹽相關基因,為進一步利用分子生物學手段深入探究其耐鹽機制奠定數據基礎。然而,對于紅樹植物,當前研究對無瓣海桑僅限于利用轉錄組學技術開展了鹽脅迫相關轉錄因子的挖掘,而對轉錄因子以外的其他可能參與調控鹽逆境適應過程的基因并未關注。本研究聯合二代和三代轉錄組測序技術,獲得了無瓣海桑對照組及500mmol·L-1鹽脅迫處理10d后的根系轉錄組數據,在此基礎上進行生物信息學分析,最終篩選出89個無瓣海桑根抗鹽候選基因,其中一些基因對鹽脅迫的響應起著十分重要的作用。
在植物的ROS酶促清除機制中,SOD、過氧化物酶(POD)、CAT是常見的酶。研究表明,NaCl脅迫后耐鹽性強的A34株系葡萄根系中的SOD、CAT、POD活性表現出快速升高并在較高水平后小幅度降低的趨勢,這有利于維持活性氧代謝的平衡,緩解細胞膜脂過氧化。魯琳等通過高通量測序技術對鹽脅迫下花煙草(Nicotiana alata Link et Otto)的活性氧清除基因進行挖掘,發現編碼SOD、POD、CAT的基因均發生了顯著的上調表達,同時存在下調表達的基因。本研究也發現了類似的結果,在SOD中,SOD1 2個上調,1個下調,SOD2 1個上調表達;CAT5個基因全部上調表達;在POD中,存在較多下調表達的基因。上述結果推測可能是由于植物體內的抗氧化機制存在著正負2種調控方式,且不同抗氧化酶在不同植物中的調控方式可能有所不同;對于無瓣海桑來說,CAT可能是以正向調控為主導,而POD可能主要是以負向調控為主。
在逆境下,植物可通過調節海藻糖的含量,穩定蛋白質的結構與功能,提高在逆境中的生存能力。李輝等克隆了海藻糖合成關鍵酶基因海藻糖-β-磷酸磷酸酶基因(trehalose-β-phosphate phosphatase,TPP),并研究其在非生物脅迫下的作用,結果表明該基因表達顯著上調。研究表明,高鹽脅迫主要促進了龍須菜(Asparagus schoberioides Kunth)海藻糖-6-磷酸合成酶/磷酸酶1基因(trehalose 6-phosphatesynthase/phosphatase 1,TPS1)、海藻糖-6-磷酸合成酶,磷酸酶2基因(trehalose 6-phosphatesynthase/phosphatase 2,TPS2)和海藻糖-6-磷酸合成酶/磷酸酶4基因(trehalose 6-phosphatesynthase/phosphatase 4,TPS4)的表達,而滲透脅迫抑制了TPS1、TPS2和海藻糖-6-磷酸合成酶,磷酸酶3基因(trehalose 6-phosphate synthase/phosphatase 3,TPS3)的表達。本研究發現2種共7個合成海藻糖的關鍵基因:海藻糖-6-磷酸磷酸酶基因(otsB)和海藻糖-6-磷酸合成酶基因(TPS)。在這些基因中,3個表現出下調表達,可能是由于鹽脅迫產生的滲透脅迫抑制了這些基因的表達。
脯氨酸是植物中常見的有機滲透調節物質,而脯氨酸的積累是植物應對鹽脅迫的普遍反應。在大多數植物物種中,δ-1-吡咯啉-5-羧酸合成酶(P5CS)催化谷氨酸生物合成脯氨酸的第一步。在Funck等的研究中,發現在擬南芥(Arabidopsis tha/iana (L.) Heynh)NaCl處理的p5cs2突變體中,幾乎沒有Na+的積累,植株既沒有褪綠,也沒有光合作用減弱,可見P5CS2介導的脯氨酸合成在調節葉片Na+積累和抗鹽脅迫方面具有重要調控功能。本研究中發現2個P5CS基因,1個上調,1個下調,表明P5CS基因在滲透調節中發揮了作用。
脫落酸在鹽、干旱、低溫等非生物脅迫下會在植物體內被誘導產生,同時啟動相關的信號途徑,以響應非生物脅迫。ABA信號途徑由ABA受體、PP2C、SnRK2和ABF轉錄因子四部分組成。研究表明,鹽處理抑制了擬南芥中PP2C31基因的表達,且在不同時期鹽處理下PP2C31基因出現了不同程度的下調表達,說明PP2C31蛋白在擬南芥響應鹽脅迫中起負調控作用。本研究發現4個PP2C基因均下調表達,推測PP2C蛋白負調控無瓣海桑的鹽脅迫響應。
絲裂原活化蛋白激酶( MAPK)途徑在鹽、高溫、病原菌等逆境的應答中起著重要作用,它通過MAPKKK→MAPKK→MAPK逐級傳遞放大脅迫信號,由下游的MAPK激活調控相應基因的表達,引起植物對逆境的應答。在野生大麥(Hordeum spontaneumL.)中,MEKK1-MKK2-MPK4/6聯級系統參與了鹽脅迫的響應。擬南芥的MAPK聯級中多個MAPK基因在多種非生物脅迫的響應中起著重要作用。汪芳珍等通過對沙芥根進行6h50 mmol·L-1的鹽處理實驗,結果表明,共有11個MAPK/MAPKK/MAPKKK蛋白激酶基因的表達顯著上調。本研究中發現MEKK1、MPK4、MPK6基因大都上調表達,推測這些基因在無瓣海桑的MAPK聯級途徑響應鹽脅迫中起著重要的作用,但無瓣海桑具體響應鹽脅迫的MAPK聯級系統中的基因及機制,還有待進一步研究。
植物體轉錄因子在鹽脅迫調控下游機制中發揮著重要作用。參與植物鹽脅迫響應的轉錄因子家族有WRKY、MYB、bHLH、NAC等。在鹽脅迫的響應中,WRKY家族轉錄因子存在著正負2種調控機制。在擬南芥中,RtWRKY23可提高自身過氧化物酶水平,以增強鹽脅迫耐受性。轉基因煙草中過表達的CaWRKY27抑制了抗氧化酶合成途徑相關基因的表達,在響應鹽脅迫中起負調控作用。本研究發現WRKY22、WRK Y25均呈上調表達,說明這些轉錄因子可能正向調控無瓣海桑的耐鹽性。此外,WRKY2、WRKY33呈現上下調表達,這表明了WRKY家族轉錄因子參與無瓣海桑耐鹽機制方式的復雜性。
4結論
本研究通過二代RNA-seq和三代全長轉錄組測序技術的聯合使用對無瓣海桑響應鹽逆境的轉錄調控機制進行了解析,獲得了鹽逆境下無瓣海桑根系的差異表達基因及功能注釋信息,并鑒定出一批與活性氧清除、滲透調節、植物激素、蛋白激酶、轉錄因子等相關的潛在耐鹽候選基因,研究結果對揭示紅樹植物的抗鹽機制、耐鹽型紅樹新型品種選育及沿海生態環境保護具有重要意義。
