
橫斷山脈亞高山帶幾種高山櫟林下叢枝菌根菌(AMF)調查
2023-04-29 15:10:21李鴻博黃耀華康定旭等
林業科學研究 2023年1期
李鴻博 黃耀華 康定旭等

關鍵詞:橫斷山脈高山櫟;叢枝菌根真菌(AMF);系統發育構建;分類地位
中圖分類號:Q945.31 文獻標志碼:A 文章編號:1001-1498(2023)01-0079-12
菌根是建立于植物根系和土壤菌根真菌之間的古老共生體系,在自然界中存在已有4億年之久,有研究統計,地球上大約86%的陸生被子植物能夠與真菌互作形成菌根共生體。叢枝菌根真菌(AMF)作為植物根系所在土壤環境中微生物群落的重要組成者之一,在個體層面可以促進植物對K+、Ca2+、Mg2+離子的有效吸收,幫助植物抵御逆境中來自生物或非生物的脅迫,宏觀群落層面則能夠優化植被分布,森林中地下的龐大菌根系統能夠將林木個體有機相連而形成一個物質資源共享體,加速種群建成并形成植物優勢種,從而改變生態格局,可以說,菌根真菌和植物共生,是維護森林生態系統穩定及發展的基礎。
高山櫟(Quercus spp.)作為一類廣泛分布在我國西部川、滇、藏地區的常綠硬葉櫟類植物,是橫斷山脈地區重要的野生植被群落優勢種,多以成規模的林地出現或以離散點狀方式與其他喬木混生生長于海拔1800~4800 m的地區,其熱值高、生長速度快及萌蘗更新能力強的特點對該地區生態環境保持具有重要意義。當前,外界公認松科植物共生菌根類型為外生菌根已毋庸置疑,而櫟屬植物存在外生菌根共生現象的同時,是否能與內生菌根真菌共生尚鮮有報道。楊淑嬌等對云南香格里拉高山櫟和高山松(Pinus densata Mast.)的林下菌塘微生物分布研究發現,高山櫟林下菌塘有大量AMF存在,且高山櫟林下菌塘中真菌和AMF生物量顯著高于高山松,但AMF種類豐度只有1種,綜合高山櫟和高山松林型下菌塘的多種微生物比較認為,菌塘中的微生物類型是其菌落結構組成的重要影響因素,林型則未影響林下微生物群落特征構成。該研究說明在錯綜復雜的地下根系網絡中,不同類型土壤微生物之間的相互聯系往往對宿主植物林下菌群結構和分布具有重要影響。張中峰將摩西球囊霉(Glomus mosseae (Nicol.&Gerd.)Gerd.&Trapp)和根內球囊霉(Glomus intraradices Schenck&Smith)接種至青岡櫟(Quercus glauca (Thunberg) Oersted)幼苗發現,AMF能夠顯著促進青岡櫟幼苗葉面積和生物量增長,提升幼苗成活率,該結果表明,在人為干擾措施下,櫟屬植物能夠與AMF形成互惠共生關系。基于上述前人研究不難推測,AMF與高山櫟組植物間應當存在相應的聯系。這種聯系是否具有廣泛性和普遍性?同為被子植物門下的櫟屬植物高山櫟根系在非人為干擾的野生生境中,其根際及根系是否同AMF共生等問題值得深入探究。基于此,本研究假設橫斷山脈亞高山地區高山櫟林下同樣具有AMF分布,并且在自然條件下高山櫟植物根系存在AMF。分三步驗證上述假設:第一,利用濕篩沉淀法對采自橫斷山脈不同地區高山櫟林下土壤樣本進行孢子濕篩,利用形態學、分子生物學結合的手段對其種類進行鑒定。第二,利用分子克隆方法選取針對AMF 18S rRNA gene的特異引物對高山櫟外根際潛在的AMF進行擴增,測序檢驗擴增結果中是否存在AMF。第三,利用染色觀察的方法觀察高山櫟根系樣本內是否存在典型的AMF叢枝、泡囊結構,結合對比單孢提取與克隆擴增結果,以期明確橫斷山脈亞高山帶自然環境中生長的高山櫟林下根圍、根際AMF種類,揭示橫斷山脈亞高山高山櫟林下AMF的演化方向。
1采樣地與材料
1.1采樣地概況
橫斷山脈作為世界年輕的山脈群之一,縱跨川、滇二省西部及西藏東部,山嶺曲折蜿蜒,谷高差懸殊,植被分布以海拔梯度差異明顯,受高空西風環流、印度洋及太平洋季風環流影響,氣候主要分為干濕兩季,降水多集中于5—10月,但因地形地勢復雜,地域遼闊,年均氣溫13.6℃,年降水量942.6 mm,部分地區降雨差異較明顯。由于該地區地勢陡峭,降雨所致的水土流失成為潛在威脅。
1.2樣品采集
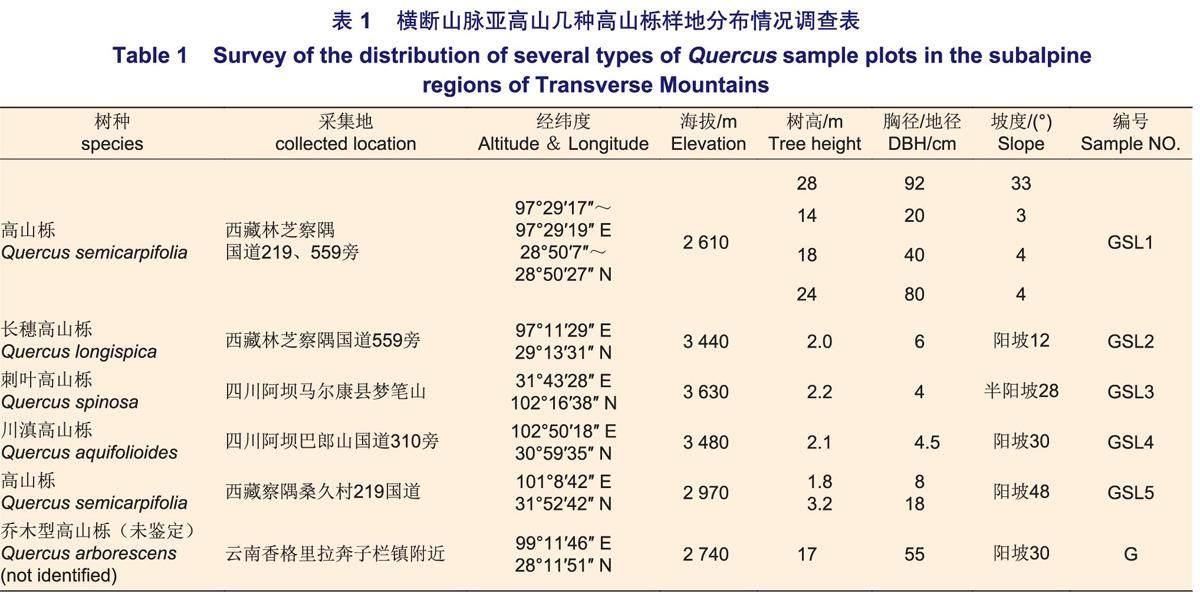
于2020年7月、10月和2021年8月分別前往位于28°45′3″~30°59′35″N,97°40′25″~102°50′18″E(四川、西藏等地)以及99°11′46″E、28°11′51″N(云南香格里拉)地段,橫斷山脈亞高山帶的不同海拔(2340~3630 m)地區(表1)陽坡地段高山櫟天然林,各高山櫟林均為喬木或灌木純林,林下無其他植被。采集其根部10~30 cm處的土壤混合裝入9號自封袋,沿主根對高山櫟根系進行挖掘,找到高山櫟根系側根、須根等與根際土裝入自封袋,放入含冰袋的保溫箱隔離貯存,標記并編號,帶回實驗室將根系于-80℃保存,土壤平鋪風干置于塑封袋用于后續實驗。
2試驗方法
2.1高山櫟根系土壤AMF濕篩與鑒定
2.1.1高山櫟根部土壤AMF孢子篩取 將高山櫟根部土壤過40目篩后,精確稱量20 g土壤,倒入玻璃燒杯,加500 mL超純水浸泡12 h后,利用濕篩沉淀法過4目(100目165μm、140目105μm、200目74μm、300目50μm)篩出并將每一目篩出的孢子及細土倒入容積為1L的燒杯靜置,靜置15min后倒300 mL入封液漏斗,后向燒杯內加水至500 mL。靜置15 min后向漏斗中填200 mLddH2O,此步驟重復3次。后靜置15min并棄漏斗廢液300 mL。將每一目篩孢子用的封液漏斗置于鐵架臺,使用抽濾機(Rocker 600)將孢子抽濾至實驗前預備好的畫有方格的濾紙上,編號標記并放入4℃冰箱保存備用。
2.1.2孢子形態學鑒定與孢子密度統計 在體式鏡(LAICA SE6)下從培養皿中的濾紙上挑出形態、顏色等不同的孢子,單個置于滴加ddH2O的載玻片上在光學顯微鏡(LEICA MC170)下觀察并拍照,記錄孢子顏色、大小,輕輕將其壓破,觀察孢壁顏色、類型、厚度、層數及內含物特征等,載玻片上還可加入Melzer's染液觀察是否存在顯色反應,并拍照保存。同時,每拍攝一個孢子將與其形態相同的孢子挑至1.5mL的離心管內-20℃冰凍保存,以便做分子鑒定時一一對應。參照《VA菌根真菌鑒定手冊》(1988)和http://invam.caf.wvu.edu網址上的圖片對照及近年來所發表的研究文獻成果,進行形態學鑒定。
在體式鏡下統計每個土樣下的孢子數目,并計算出孢子密度,每個土樣4次重復。
孢子密度(SD):指每克土樣中叢枝菌根真菌的孢子個數。
SD=某土樣中AMF所有孢子數/土壤干質量。
2.1.3孢子的分子生物學鑒定 利用Chelex-100提取單孢DNA,向裝有孢子的離心管中加入600μLddH2O,置入超聲波震蕩洗滌儀器(30Hz)震蕩3min,挑出孢子轉移至新的離心管,重復上述步驟以清洗孢子表面存在的雜質。將清洗后的單孢轉移至新的離心管中(1.5 mL),加入30μL TE buffer,置于盛有液氮的保溫瓶速凍,取已滅菌的研磨棒在體式顯微鏡下將孢子研碎。加入15μL 20% Chelex-100后置于金屬恒溫水浴鍋98℃裂解10 min,放入-20℃冰箱冰浴5 min后取出,置于離心機(Eppendorf Centrifuge 5417R)12000r·min-1離心10 min,所得上清液即為用于擴增的孢子的DNA模板。
參照《菌根學》中的擴增辦法,對孢子DNA模板進行巢式PCR擴增。第一輪擴增上下游引物為GeoLl(5'-ACCTTGTTACGACTTTTACTTCC-3),GeoA2 (5-CCAGTAGTCATATGCTTGTCTC-3')反應體系為50μL(ddH2O 20μL,Taq PCRMaster Mix 25μL,
GeoL1 1.5μL,
GeoA2 1.5μL,DNA 2μL)。94℃預變性4 min,94℃變性30s,54℃退火1min,72℃延伸2 min,其中變性、退火、延伸進行35個循環,后72℃延伸10 min。將擴增后的PCR原液稀釋10~20倍作為第二輪DNA擴增模板,第二輪PCR上下游引物為AM1(5'-GTTTCCCGTAAGGCGCCGAA-3')和NS31-GC(5'-TTGGAGGGCAAGTCTGGTGCC-3')94℃預變性2 min,94℃變性45s,55℃退火1min,72℃延伸45s,其中變陛、退火、延伸進行35個循環,后72℃延伸7 min,回收PCR擴增產物進行1%瓊脂糖凝膠電泳(110V,25min)經凝膠成像系統(Molecular Imager GEL Doc TM XR)檢測,觀察結果,拍照并記錄。將條帶清晰的產物送擎科生物公司(TSINGKE昆明)測序分析。
2.2高山櫟根際潛在的AMF鑒定
2.2.1高山櫟根際AMF分子克隆 為證明高山櫟根系內外根際存在AMF,取-80℃冷凍保存的西藏、四川(GSL1-a,GSL4-a)以及香格里拉地區的高山櫟根系(G)樣本,置于流水下沖洗根系表面后,分別剪成1cm長的小段放入1.5 mL的離心管中,加入75%的酒精后劇烈震蕩30 s,將根系取出至ddH2O下沖洗,后放入已滅菌的研缽加液氮速凍并研磨至粉末狀。利用試劑盒(EZUP柱式植物基因組DNA抽提試劑盒,SANGONG),嚴格按照盒內使用說明書的操作步驟提取高山櫟根系總DNA,使用AMF的特異上下游引物AML1,AML2對高山櫟內外根際潛在的AM真菌DNA進行擴增。反應體系為50μL(ddH2O 24.8μL,TaqPCRMasterMix16.2μL, AML1、AML2各2.5μL,DNA 4μL)。步驟為94℃預變性4 min,94℃變性30s,54℃復性1min,72℃退火2 min,其中變性、復性、退火進行35個循環,后72℃延伸10 min。之后經凝膠電泳(同2.1.3)檢測后,利用PCR產物純化回收試劑盒,純化回收產物,使用克隆試劑盒(零背景TOPO-TA克隆試劑盒)嚴格按照使用說明將回收的DNA插入質粒載體。于37℃在LB液體培養基中培養1h(將液體培養基放入搖床180~200 rpm震蕩培養)。培養完成后在超凈工作臺下取200μL菌液涂布于提前加入氨芐青霉素(AMP)的LB固體培養基上,待培養基吸收后封存,37℃恒溫培養12~16h。
在超凈工作臺下將培養好的菌落用無菌的移液器吸頭(量程為20μL)挑人提前加裝10μL ddH2O的PCR管并抽吸混勻,利用AML1和AML2引物對DNA模板進行擴增。反應體系為50μL(ddH2O22.8μL, Taq PCR Master Mix 20.2μL, AML1、AML2各2.5μL,DNA2μL)。94℃預變性2min,94℃變性45s,65℃復性1 min,72℃退火45 min,其中變性、復性、退火進行35個循環,后72℃延伸10 min。經凝膠電泳檢測后將擴增結果良好的樣本送至測序公司(SanGong昆明)測序。
2.2.2高山櫟根系AMF染色觀察 分別將2次采集的根系須根樣本剪成0.5~1 cm的小段,置入盛有10% KOH溶液的試管于水浴鍋90℃脫色處理,直至根系顏色變淡,用蒸餾水反復沖洗后加入0.5%的苯胺藍染液繼續90℃染色1 h,重復沖洗步驟用3%乳酸、甘油脫色2~3 d至半透明后,置于顯微鏡下觀察根系內的AMF泡囊叢枝結構。
2.3數據處理
利用Chromas2.22、觀察序列峰圖、DNAman 8剪貼與拼接序列。將序列提交至NCBI基因數據庫比對并下載同源序列。使用Mega X多序列比對并篩選建樹模型,選擇參數值最高的推薦模型,利用最大似然法1000次迭代循環檢驗構建系統發育樹。
運用EXCEL2016統計孢子密度數據及擴增樣本序列比對數據。
3結果與分析
3.1幾種高山櫟土壤AMF孢子形態及土壤孢子密度特征
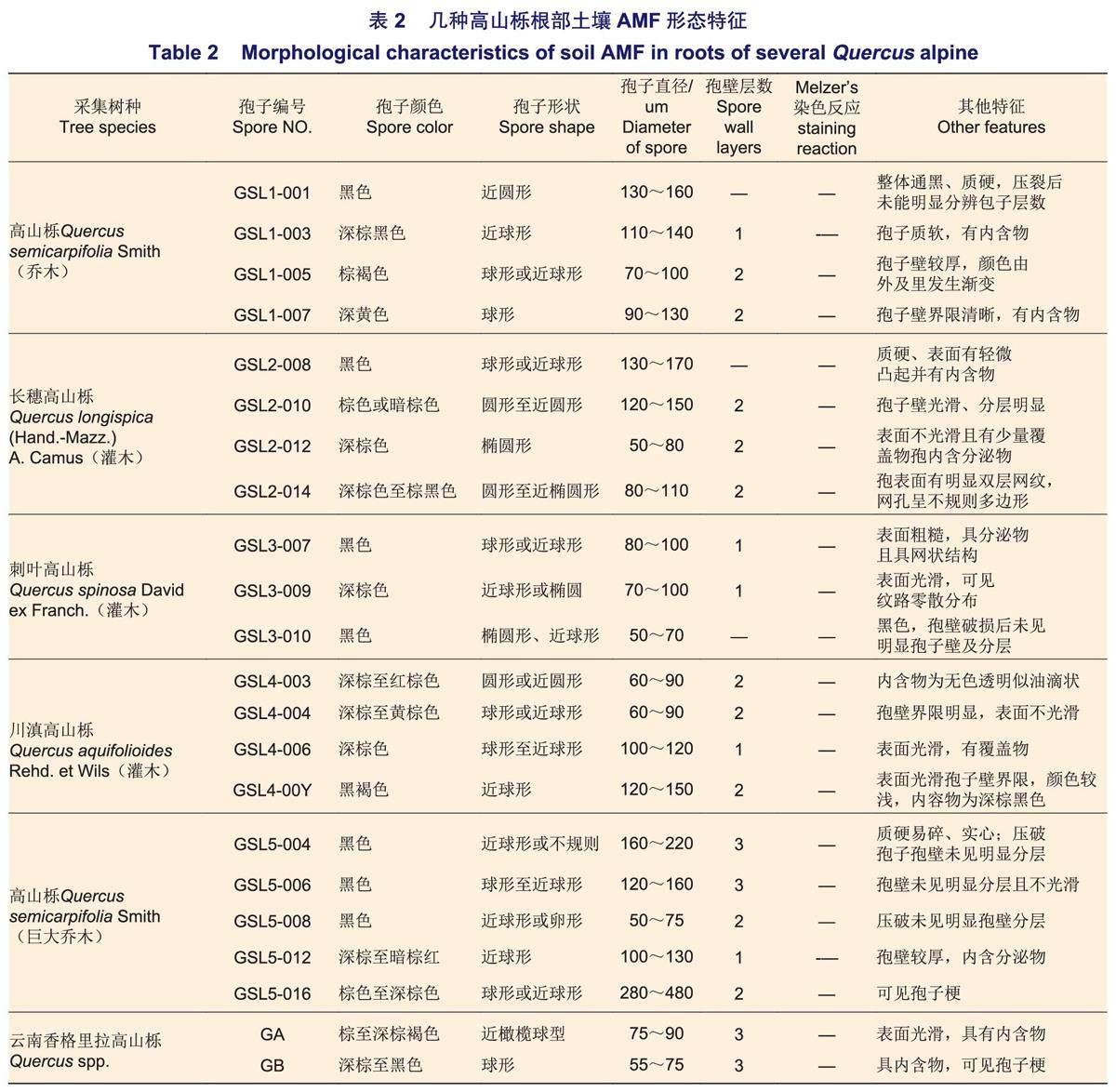
對高山櫟根部10~30cm土壤的篩取結果表明:各高山櫟樣地下均有大量AMF孢子存在,孢子直徑為50~220μm,形狀以球形、近球形為主。顏色多以黑色和深棕色出現,偶見棕黃或深棕紅,其中,部分孢子呈黑色,該類孢子在顯微鏡下壓裂未見明顯孢壁層數,部分孢子有分泌物滲出(表2,圖1)。目前,經實驗觀察的孢子均未見Melzer's染液染色反應。5種高山櫟根系土壤樣本中孢子密度分別為GSL2(205.49)>GSL4(178.90)>GSL1(88.39)>GSL3(43.84)>GSL5(16.16)個·g-1,不同高山櫟土壤樣本具有明顯差異,且呈喬木高山櫟大于灌木的趨勢(圖2),其中,GSL3為灌木但表現出孢子密度相對較低的特征。各樣本孢子100目下(直徑>165μm)以黑色硬質孢子為主,多數孢子密集分布于300目篩(50~74μm)內,呈深棕色。
3.2結合形態學和分子生物學手段對高山櫟林下菌塘AMF孢子及根系AMF克隆的鑒定
對西藏、四川各高山櫟根圍土壤中的AMF分子鑒定結果表明:橫斷山脈不同海拔地區采集的高山櫟土壤AMF均與類球囊霉科(Paraglomeraceae)類球囊霉屬(Paraglomus)具有較近的親緣關系(表3)。經西弗吉尼亞大學孢子比對網站(occultum|Davis-INVAM|West Virginia University (wvu.edu))比對發現,Paraglomus屬下目前收錄的菌種僅有巴西類球囊霉(Paraglomus brasilianum(Spain&J.Miranda)J.B.Morton&D.Redecker)和隱類球囊霉(Paraglomus occultum (C.Walker)J.B. Morton&D.Redecker),但這2種AM孢子在顏色、質地、孢壁層數與Melzer's染液反應效果與目標菌種均明顯不同。利用Mycobank檢索AMF Paraglomus屬,選取Gosling等和Mello等發表的AMF Paraglomus屬的幾種序列以及MAARJAM (ut.ee)中與本研究孢子相似度高于90%(相似度為90.9%~99.2%)的序列對幾種高山櫟林下AMF孢子進行系統發育樹構建。結果表明:明確到種的Paraglomus序列聚為一支,MAARJAM (ut.ee)中ParaglomeraceaeParaglomus sp.屬序列與樣本GSL2-014聚為一支(圖3),其余所有孢子與根系AMF克隆樣本能夠單獨聚為一支,其中,同源序列相似度高于97%的個體極有可能為類球囊霉屬內某種真菌。
3.3云南香格里拉高山櫟根際AMF孢子和根系AMF的分子克隆
利用AMF特異引物AML1、AML2對香格里拉地區的高山櫟根系潛在AMF的ITS片段DNA克隆,結果表明:經75%濃度酒精對高山櫟根系表面消毒30s后,其根系表面仍存在AMF的DNA。通過MAARJAM (ut.ee)網站比對發現,存在真菌普遍與球囊霉屬Glomus在Genbank中的標準序列相似度較高,部分目標菌種序列與Genbank中的序列對比有相似度較高的類球囊霉屬Paraglomus序列存在(表4),但其在覆蓋度(Query coverage)或最大識別度(Max identity)2個指標并非最高。利用Bio-edit軟件檢驗克隆所得目標AM菌種同源性,發現17個菌種均為同一菌株,故選擇G1為代表,與該樣本下AMF孢子序列(GA、GB)一同構建系統發生演化樹(圖4),分析表明:目標菌種樣品G1、GA和GB與Glomus versiforme(Y17651.1)聚為一支,說明其親緣關系與該屬較近,置信度可達98%,推測高山櫟能夠和與Glomus versiforme同源性較高的AMF形成共生關系,且其與林下菌塘孢子類似,二者均未能鑒定到種。說明能夠與香格里拉地區高山櫟根系共存的AMF極可能為球囊霉屬和類球囊霉屬,或與其并列的某一新屬真菌。
3.4幾種高山櫟根系AMF泡囊叢枝結構觀察
幾種高山櫟根系苯胺藍染色結果表明:100~200倍放大時可見明顯的類泡囊和叢枝結構,此類結構在高山櫟根系內主要呈圓形或近圓形(圖5a、b),部分呈橢圓形,部分泡囊分布密集,并伴隨“向無規則方向伸展”的橫向菌絲結構,推測其為高山櫟根系于AMF共生所形成的泡囊和叢枝。
4討論
4.1高山櫟根圍土AMF的驗證
自然界中能夠獨立生存的生物少之又少,幾乎所有生物之間普遍存在共生關系。AM作為一種重要的功能型土壤微生物,其與宿主植物間形成的共生體系能夠使宿主植物在群落構成及其對外界生境在生態適應性層面產生深遠影響。調查野生生境中的AMF不僅能夠擴充AMF菌種資源庫,有利于發掘AMF在不同宿主植物根系下的群落構成及多樣性,且在探究其對于生態系統平衡穩定的可持續發展層面具重要意義。
橫斷山脈一直以來是生物多樣性研究的熱點地區,不少學者研究認為,4000~5000萬年前印度次大陸與歐亞板塊碰撞導致的青藏高原隆起,隨后該地區可能成為東亞北溫帶植物的避難所。楊欽周認為,橫斷山脈縱向深切及垂直氣候帶的明顯分布促使硬葉櫟類古老櫟群的殘遺保留和新生類群的分化形成。因此,選擇古老的高山櫟組植物對其根系潛在的AMF進行研究,從演化發育的角度能夠更有助于理解并揭示該地區AMF的起源、營養類型、生活方式及分類問題。
本研究利用濕篩沉淀法從形態學和分子生物學結合的視角證實高山櫟林下菌塘中存在AMF真菌孢子,這與楊淑嬌等基于磷脂脂肪酸(RLFP)方法研究香格里拉高山櫟林下菌塘微生物的結果一致,驗證了AMF在野生條件下能夠廣泛存在于橫斷山脈亞高山地區高山櫟林下的假設。西藏、四川等地各土壤樣本中的孢子18S rRNA gene分子證據表明,所有孢子均與類球囊霉屬Paraglomus的親緣關系較接近,通過將各孢子的形態學特征與已證實或發表的孢子形態對比發現,其在顏色、質地、孢壁層數和Melzer's染液反應效果等方面均與目前已知的巴西類球囊霉、隱類球囊霉及其他幾種該屬下發表的新種具有明顯差異,故認為該地區高山櫟林下AMF很有可能為類球囊霉屬下的未知種。由于其所處地理位置植被分布的古老與獨特性,結合18S rRNA gene序列相似度比對分析認為,其或在漫長演化進程中在Glomeraceae和Paraglomeraceae科便已出現分化。基于系統發育樹(圖3)對橫斷山北部西藏四川等地區孢子的演化進行推測,GSL2-14的演化介于Paraglomus和Archeaspora之間,而GSL1-001、2-008、3-007、4-003等則獨立分為一支,與前者呈并列關系,這與前人基于真菌分類學為“科”級別將Paraglomus和Archeaspora從Glomus中分離出去的分類學定位相吻合。
4.2高山櫟根系AMF擴增結果的提示
由于目前AMF分子研究技術仍舊有一定的局限性,即缺乏一套通用的既能擴增所有AM家族,又能排除非AM真菌的引物。因此,為保證克隆質量,選擇AML1和AML2作為引物擴增高山櫟根系(樣本GSL1-a、GSL4-a、G)AMF的18srRNA基因,發現與四川、西藏等地高山櫟克隆菌株相似度較高的AMF均為類球囊霉屬Paraglomus,與香格里拉地區高山櫟克隆菌株相似度較高的AMF則均為球囊霉屬Glomus。系統發育樹構建結果表明:Glomus versiforme與后者AMF同源性極高(圖4),西藏、四川與香格里拉等地高山櫟根系AMF結果存在一定差異。這種現象出現的原因可能是AMF在長期演化過程中隨地域環境變化(從西藏、四川到云南)和不同高山櫟種(表1)而逐漸發生分化。同時,本研究克隆結果與張中峰等將摩西球囊霉和根內球囊霉接種至青岡櫟幼苗的結果類似,說明櫟屬植物在非人為干擾的野生環境下根系也存在AMF。第二,PCR擴增過程中由于DNA模板中存在殘留的酚類及腐殖酸等,此類物質難以去除并會對DNA擴增造成一定影響。引物不同,擴增過程中的靶向序列堿基對存在差異,并且擴增時堿基錯配的概率也可能呈倍放大,從而造成序列比對時發生單孢提取與分子克隆出現結果上的差異。此外,高山櫟根系、土壤樣本均分別僅有一種AMF出現,可能提示不同高山櫟同時僅與一種AMF在內外根際范圍產生聯系。有研究表明,碳源和植物內源激素能夠對AM產孢能力產生重要影響,二者條件豐富的情況下孢子萌發率較高。同理,高山櫟生長在半陽坡地區,當地氣候、海拔及水熱條件使得土壤生境中的碳源及高山櫟宿主植物內源激素發生變化,進而一定程度干擾AMF產孢,而Paraglomus和Glomus在高山櫟根際微生物的生態位競爭過程中占據了相對優勢地位,以此來確保自身所需碳源的充分供給。
綜合橫斷山脈亞高山帶幾種高山櫟林下菌塘AMF孢子調查及其根際AMF的克隆結果發現,高山櫟組植物作為典型的被子植物,在與外生菌根真菌發生共生現象的同時,極有可能在自然環境中保留了與AMF存在共生的現象。在喜馬拉雅造山運動后,橫斷山區長時間與外界環境隔離,土壤微生物在漫長的演化進程中由于地理環境的相對封閉而發生分化,最終形成了現有存在的AMF孢子,而該類孢子隨區域(從西藏、川西到云南)變化顯示出一定的演化差異,具體體現在其形態及分子學方面的不同。因此,在系統發育進化關系的構樹結果中,該類孢子以單獨匯聚為一支呈現。
5結論
本研究利用形態學與分子生物學的手段證明,自然條件下橫斷山脈亞高山地區高山櫟林下菌塘普遍存在AMF,利用分子克隆手段發現AMF可分布于高山櫟植物根系,且發現西藏、四川等地該類孢子在形態上與已知AMF屬種存在較大差異,極有可能為Paraglomus屬下的新種。同時,云南香格里拉地區高山櫟根系AM與Glomus屬更接近,說明AM在長期演化過程中隨地域遷移出現分化。本研究結果為揭示AMF對橫斷山脈亞高山帶高山櫟生態適應性機制的影響提供新的證據,為進一步了解橫斷山脈亞高山帶高山櫟林下AMF的形成與演化提供理論參考。
