太行山獼猴寰椎橋的類型與特征
2023-04-29 00:44:03趙曉進郭夢丹張夢琪鹿雪涵
人類學學報 2023年6期
趙曉進 郭夢丹 張夢琪 鹿雪涵
摘要:為了確定太行山獼猴(Macaca mulatta) 的椎動脈溝上3 種骨橋(腹橋、側橋和背橋)的分布特征,以期理解在人類寰椎上觀察到的變異,并推測靈長類動物可能的進化趨勢。本文分別測量57 例(雄性17 例,雌性40 例)獼猴寰椎標本(成年骨骼標本)3 個寰椎橋的寬度。并采用SPSS 統計軟件進行數據處理,性別之間差異采用方差分析,觀察寰椎橋的出現率、形態特征和組合類型。統計結果顯示,獼猴寰椎橋變量性差顯著(P<0.05),獼猴的腹橋和背橋是恒定出現(100%),側橋基本恒定,但也有一些缺失個案(90.3%),表明腹橋和背橋是獼猴穩定的特征;獼猴寰椎橋的主要類型為原始的A 型,與主要類型為D 型的人科動物發生分離。我們基于以上特征后得出結論,獼猴的3 個寰椎橋是一種穩定性狀,其出現率明顯高于類人猿。
關鍵詞:寰椎;骨橋;獼猴;靈長類
1 引言
人類寰椎的正常變異和臨床意義受到國內外學者的關注,寰椎作為頸部的重要解剖結構也愈加受到重視。歐海寧等[1] 通過大數據分析了人類寰椎后橋的出現率和臨床表現后得出結論:發生在人類寰椎后橋是頸椎骨質結構的解剖學變異,寰椎后橋的出現及解剖學特征與臨床癥狀有一定的相關性。了解寰椎橋的結構、特征、起源和種群差異,有助于了解靈長類動物的個體發育和系統演化,探討靈長類動物可能的進化趨勢[2]。
薛德明等曾對太行山獼猴寰椎和樞椎進行了初步研究[3,4]。肖洪文等針對人類寰椎橋的測量、分型、變異和臨床意義等展開系列的研究,豐富了寰椎橋的研究內容,推進了寰椎橋的研究進展[5]。Le Minor 等觀察了部分靈長類動物的寰椎標本,探討寰椎橋在靈長類動物中的分布和類型,試圖解釋人類觀察到的寰椎橋變異[6]。目前,對于在人類種群中出現寰椎橋變化,出現寰椎橋變化的真正原因尚不清楚,提出的解釋以理論性為主。Yammoto 等對328 只日本獼猴寰椎橋的研究表明,日本獼猴的寰椎主要類型是背橋,側橋的出現率略低于背橋[7]。推測寰椎背橋是日本獼猴的一個穩定性狀,其背橋出現率明顯高于人類。盡管人們已經認識到寰椎橋結構在人類進化過程中的重要性,但迄今為止,國內關于非人靈長類動物寰椎橋的研究一直較少。
本文研究目的在于觀察太行山獼猴寰椎橋的形態學特征和分布,描述性統計其性別差異,探討不同靈長類動物寰椎橋的種群差異和進化趨勢。
2 材料與方法
2.1 研究材料
本文的研究材料為太行山獼猴寰椎標本57 件(雄17,雌40),均按照標準制備方法完成。標本完整、對稱、無任何病例征象(圖1)。根據齒式和骨骺愈合可以確認標本是成年的,性別是已知的。標本收集時間是1998~2022 年,標本全部來自河南省太行山獼猴自然保護區。目前保存在鄭州工商學院商學院。
2.2 寰椎橋的定義和分類
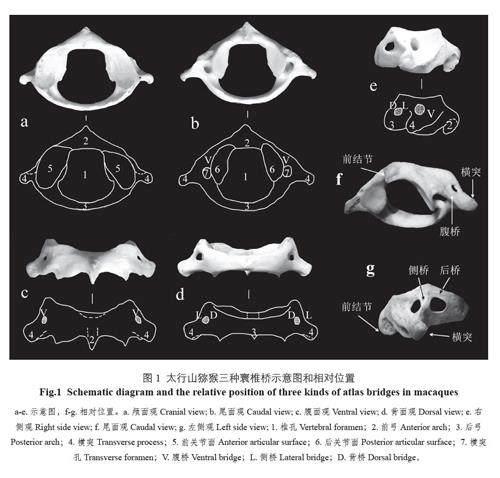
根據文獻對寰椎橋進行定義,理論上寰椎椎動脈溝上可能存在3 種骨橋,分別是腹橋、側橋和背橋[6,7](圖1: a-e)。腹橋由寰椎橫突孔的腹側結構構成,椎動脈通常在此通過,部分對應于橫突的前根,是顱面橫突孔與尾面橫突孔在腹面觀的直線距離,腹橋如果缺失一般認為是橫突前根缺失。側橋位于寰椎側塊的外側,椎動脈溝上方,文獻中常常會出現各種不同的命名(側骨橋、外側橋等);從寰椎的顱面觀上看得清楚,也容易測量,是橫突孔與橫向上孔之間的直線距離。背橋(人類寰椎上通常稱為后橋)位于寰椎側塊的背側,椎動脈溝的上方,在寰椎的背面觀上看得很清楚,是橫向上孔與側塊上緣的直線距離。三種寰椎橋的相對位置見圖1: f-g。
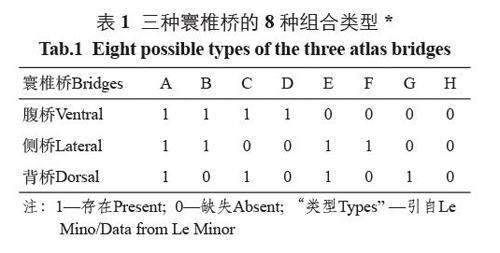
依據是否出現寰椎橋,可以有8 種組合類型,記作A、B、C、D、E、F、G、H(表1)[6]。
2.3 研究方法
本文對寰椎橋進行定量分析,采用人類體質學骨骼直接測量法。測量儀器為國際標準電子數顯卡尺,單位是mm,精確到0.01 mm。所有標本的測量均由本文第一作者獨立完成。變量選擇是參照文獻資料[8],測量變量為三種寰椎橋的橋寬。數據處理采用IBMSPSS 26.0 統計軟件。如果側別之間沒有統計學差異,性別組間統計檢驗可以采用兩側合并數據 [9,10]。性別組間檢驗采用單因素方差分析(Variance analysis, VA)。
3 結果
3.1 寰椎橋描述性統計
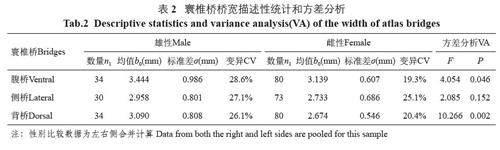
太行山獼猴3 種寰椎橋測量性狀性別組間的比較結果表明,腹橋橋寬的F=4.054,P=0.046;背橋橋寬的F=10.266, P=0.002, 性別組之間腹橋和背橋有統計學差異(P<0.05);側橋橋寬的F=2.085, P=0.152, 性別組間沒有統計學差異;3 個寰椎橋變量都是雄性大于雌性(表2)。
3.2 獼猴寰椎橋的出現率
太行山獼猴3 種寰椎橋的出現頻數和百分比統計結果顯示:獼猴的3 種寰椎橋出現率有一定的規律。腹橋和背橋出現穩定,出現率為100%;側橋出現率略低,雙側合并后出現率為90.4%,雙側同時出現率為86.0%(表3)。
從非測量性狀來看,首先是寰椎橋的數目不同,獼猴一般在寰椎側塊上有3 個骨橋,分別稱為腹橋、側橋和背橋。寰椎橋的數目差異是區別人類與猿類的關鍵。從形狀和大體解剖結構來看,人類與猿類的寰椎橋非常相似,但有一個明顯的差異,即獼猴的寰椎顱面觀一般看不到橫突孔,尾面觀橫突孔顯著,基本上是圓形。在獼猴的寰椎腹面觀可以看到一個比較小的橫突孔(圖1)。綜上所述,太行山獼猴的橫突孔有兩個特征,一是橫突孔在顱面觀和尾面觀不是貫通的;二是顱面觀側塊上的橫突孔一般比尾面觀上的橫突孔要小,而且形狀不規則,有時側橋和腹橋會發生融合,導致該孔消失。
獼猴腹橋在顱尾側也不是扁平的,腹橋的寬幾乎比橫突上向尾面伸出的脊還要小。其重要原因是位于顱面觀上的橫突孔比較小且不規則,和人類的腹橋形態完全不同。獼猴側橋形狀極不規則,有時很難辨認,只能通過橫突側面上的兩個孔來識別,橋寬用兩個孔之間的最短距離來表示。本研究中側橋的缺失率較高,說明變異性較大。側橋缺失的原因可能是側橋在顱面觀的橫突孔一般大小不等,兩側一般不對稱,有時會在單側或雙側消失,所以,側橋和腹橋會融合形成較大的腹橋,獼猴寰椎背橋是獼猴類的典型特征,形狀規則、扁平、穩定,容易測量和識別。
3.3 不同種群靈長類動物寰椎橋比較
由表4 可知,寰椎橋的出現率有明顯的差異。婆羅洲猩猩(Pongo pygmaeus) 腹橋出現率為95.8%,側橋出現率為0,背橋為66.7%。人類(Homo sapiens) 腹橋出現率為92.6%, 側橋為1.0%,背橋為10.1%。在原猴猴亞目(Strepsirhine)、蛛猴亞科(Atelinae) 和絹毛猴科(Callitrichine) 中,側橋卻是恒定出現,或者幾乎穩定出現(99.0%~100%)。猩猩和人類中,后橋出現率明顯減少,分別為66.7% 和10.1%;側橋的出現率更低,分別為0.0% 和1.0%。太行山獼猴(Macaca mulatta) 中,腹橋出現率為100%,側橋為90.3%,背橋為100%。在獼猴亞科(Cercopithecinae) 中,腹橋出現率為100%,側橋為91.5%,背橋為98.9%。在所有的靈長類種群中,除了猩猩和人類外,背橋出現率都非常高,范圍在98.9 %~100%(表4)。
不同靈長類種群寰椎橋類型比較見表5。靈長類動物中的寰椎橋類型有8 種,其中主要類型有3 種,分別是A 型、C 型和D 型。
A 型 3 種寰椎橋同時存在,是原猴亞目(Strepsirhini) 和猴科(Cercopithecines) 的主要類型,在黑猩猩(Pan troglodytes)、長臂猿(Hylobates) 和人類(Homo sapiens) 中偶然出現,而在婆羅洲猩猩(Pongo pygmaeus) 和大猩猩(Gorilla gorilla) 中不曾出現。太行山獼猴主要類型是A 型。
C 型 腹橋和背橋同時存在,側橋不存在,是婆羅洲猩猩(Pongo pygmaeus) 和黑猩猩(Pan troglodytes) 的主要類型,在長臂猿(Hylobates) 的出現率為33.3%,在人類(Homosapiens) 和猴科(Cercopithecines) 中偶然出現(出現率分別是9.0% 和5.7%),在大猩猩(Gorilla gorilla) 中不存在。
D 型 只存在腹橋,不存在側橋和背橋,通常僅在一些類人猿物種中占主導地位。獼猴(Macaca mulatta) 和猴科(Cercopithecines) 動物中的D 型十分罕見或缺失(0.0%~0.3%)。
H 型 3 種寰椎橋同時消失,在大猩猩(Gorilla gorilla)、黑猩猩(Pan troglodytes) 和人類(Homo sapiens) 中偶然出現,出現率在6.9% ~11.9% 之間(表5)。
4 討論
將寰椎橋簡單地確定為存在或缺失,顯然不能完全反映出骨骼形態特征所表達的生物學意義[6]。孫靜宜等對人類寰椎后橋進行了形態學分析后認為,背橋是人類進化過程中的退化痕跡,狹窄的寰椎動脈溝環可能是造成頸性眩暈的主要原因[2]。Yammoto 等研究了日本獼猴寰椎橋個體變化和地理變異,同時證明日本獼猴寰椎橋與年齡之間的對應關系[7]。研究結果表明,成年日本獼猴背橋和側橋普遍存在。Kikuchi 等研究發現,不同靈長類動物頸椎的形態不同,通過寰椎的體型指數和形態指數可以推測其種群的個體大小和運動類型,進一步可以推測他們的進化程度[11]。
從測量性狀來看,寰椎橋側別之間沒有統計學差異;性別之間腹橋和背橋有統計學差異,差異最大的變量是背橋寬。寰椎橋性差的特征可以解釋為種群遺傳因素和系統進化的結果,是其種群生物學屬性之一,是長期性別選擇和青春期后雌雄兩性異速生長的結果[12]。側橋寬性差不顯著,可能與側橋的變異性較大有關。側橋寬的變異系數是25.90%,極差是3.66 mm;對應背橋寬的變異系數是23.58%,極差是3.13 mm。關于寰椎骨橋的測量性狀的意義,可以解釋為這是解剖學和人類學常用的方法。很多文獻資料也對人類和非人靈長類寰椎的形態進行定量和定性研究[2,4-6]。盡管大多數骨橋的測量是以人類寰椎為對象,但獼猴是人類的近親,寰椎橋的結構,尤其是背橋是相似的,資料也顯示人類的寰椎后橋與非人靈長類的寰椎背橋是同源的。另外,通過對骨橋寬度的定量分析,還可以得到很多有價值的信息。例如,可以進行判別分析、多元回歸分析和種間定量分析等。
從系統發育的角度來看,這三個寰椎橋在靈長類動物和其他哺乳動物中具有一定的同源特征,深入研究不同動物類型的寰椎骨橋,對于進一步理解動物的系統發育具有重要意義。目前,國內這一領域研究結果相對較少,有待進一步加強。
從非測量性狀來看,太行山獼猴通常具有穩定的3 種寰椎橋結構,其中背橋和腹橋的出現率均為100%。側橋偶有缺失,缺失率往往超過10.0%;這一現象表明,太行山獼猴側橋的結構并不穩定。側橋缺失的主要原因在于寰椎顱面觀的橫突孔一般大小不等,兩側的橫突孔也總是不對稱的。另外,側橋的形狀極不規則,一般是通過寰椎橫突脊側面的兩個小孔來確認,在寰椎橫突截面上的側橋會構成錐形。有時由于側橋和腹橋融合,形成較寬的腹橋,這種特殊的寰椎腹橋有待進一步觀察。太行山獼猴寰椎后橋是獼猴屬的穩定結構,這一點與日本學者Yammoto 等對日本獼猴寰椎橋的研究結果是一致的[7]。
探討不同靈長類動物寰椎橋的起源、種間差異、影響因素和進化趨勢是有意義的。參照以前的研究資料可以推測,太行山獼猴寰椎橋的結構是一種相對原始的結構,其主要依據是:1) 許多哺乳動物中出現了類似獼猴寰椎橋的結構,與人類的寰椎橋結構完全不同[5];2) 從表4 可以看出太行山獼猴(Macaca mulatta) 的3 種寰椎橋的出現率與一些低等靈長類動物的十分相似,如原猴亞目(Strepsirhini)、蜘蛛猴亞科(Atelines) 和絨猴亞科(Callitrichines) 等[5]。
非人靈長類動物的三個寰椎橋同時存在為原始類型(A 型)。在靈長類動物中能夠觀察到的從原始類型(A 型)以及派生出來的6 種寰椎橋的類型(表1)。A 型主要出現在原猴亞目(Prosimians) 的一些科和屬、闊鼻類(Platyrrhines) 的一些科、猴科(Cercopithecinae)以及獼猴屬(Macaca mulatta) 等。由原始類型(A 型)派生出來的其它類型包括3 個層次,第一層次是C 型(只有腹橋和背橋),這一類型主要包括跗猴科(Tarsiids, 100%)、疣猴屬(Colobus, 85%) 和猩猩屬(Pongo pygmaeus, 62.5%);第二層次是D 型,這一層次主要包括大猩猩(Gorilla gorilla, 89.5%)、人科(Homo sapiens, 82.8%) 和長臂猿屬(Hylobates,63.9%) 等;第三層次是H 型,這一類型只在人猿超科中出現,大猩猩(10.5%)、黑猩猩(Pantroglodytes, 11.9%) 和人科(6.9%)。
從進化趨勢上來看,可以將所有的靈長類動物大致分為3 個組。1) 原猴亞目- 闊鼻類-猴科組,主要是原始類型A 型;2) 疣猴屬- 猩猩屬- 黑猩猩屬組,主要是由A 型派生出來的C 型;3) 長臂猿- 大猩猩- 人科組,主要是由C 型派生出來的更高級別的D 型。從整體結構上看,與人類最近似的猿類就是大猩猩,這點確定無疑[13]。總之,寰椎橋的逐漸消失似乎構成了靈長類動物的一種進化趨勢。
考慮到靈長類的系統發育,各種寰椎橋類型之間也存在著內在的演變關系。例如,原猴類被公認是現存靈長類動物中最原始的,寰椎橋表現出原始類型(A 型)。眼鏡猴科(Tarsiidae) 屬靈長目簡鼻亞目跗猴型下目的一個科,出現于距今6000 萬年以前,一直延續到現代,是一支高度特化的靈長目動物。簡鼻亞目與原猴亞目約于6300 萬年前演化分開,所以眼鏡猴科會有一些原猴亞目的特征,同時也混合了類人猿的一些特征。例如,眼鏡猴的嬰兒相較于原猴更大,但較依賴母親,這可能是因它們復雜的行為及自然歷史所致。眼鏡猴科的寰椎橋也確實表現出介于原猴類與類人猿之間的模式(C 類)。
本文對觀察到的太行山獼猴寰椎橋的分布也可以進行解釋。寰椎橋主要類型為原始的A 型,同時,出現少量的C 型,沒有B 型,因此推斷太行山獼猴寰椎橋介于原始靈長類動物與高等靈長類動物之間,與Le Minor 等和Yammoto 等研究結果一致[6,7]。類人猿亞目也明顯表現出寰椎橋的多態性,黑猩猩的寰椎橋的多態性似乎比人類要更大一些。寰椎橋的多態性曾一直被人們所忽視,實際上寰椎橋多態性對于物種的形成或進化具有特殊的意義,是物種進化和生物多樣性形成的基礎。
有關寰椎橋的起源一直存在爭議。部分學者認為是哺乳動物先天正常發育的結果,特別是翼手目和食蟲目頸椎上斜突的同源物。Yamamoto 等卻認為寰椎背橋是由寰枕后膜骨化形成,很明顯這種骨化過程與年齡增長有關[14]。Traize 等的研究進一步證明青年人似乎是以部分不完整的橋為主,而老年人主要以完整橋為主;部分橋可能是完整橋的前體[8]。目前,還沒有證據證明,成年人的寰椎背橋會隨著年齡的不同而發生變異。本研究觀察發現,幼年太行山獼猴的寰椎標本上的確可以看到不完整的背橋,但更像是動物正常發育的過程;成年太行山獼猴有著完整穩定的背橋結構(100%),而這似乎與衰老過程無關;寰椎背橋應是獼猴的一種正常結構,具有種遺傳屬性,是系統演化和長期適應環境的結果。
本文依據Finnegan 等提出的寰椎橋分類方法進行統計描述[15]。這是一種新的嘗試,也產生了一些新的認識,概述如下:1) 簡單地將寰椎橋分為存在與缺失似乎與實際情況不符,因為在寰椎橋存在一些中間過渡類型(例如半橋)。Taitz 等的研究結果顯示人類不同種群的部分橋出現率不同,白人是1.4%~5.2%,黑人是6.0%~10.4%,印度人是37.4%,中東人是55.2%,總體上來看,部分橋或不完整橋的出現率是25.9%[8]。太行山獼猴寰椎橋的觀察中也發現了這種情況,值得深入研究。2) 將非人靈長類動物的寰椎橋定義為8 種類型是為了進行研究對比,并非唯一的選擇;一些寰椎橋根本不出現,或者一些橋類很難歸類。 3) 除了考慮是否存在外,還要考慮寰椎橋非測量性狀的形態特征。
例如,太行山獼猴3 種寰椎橋的形態各異,腹橋寬扁且向上隆起,側橋形狀極不規則,橋面向上突起構成橫突,這些橋之間的空間非常小,形成了一個微小的孔。4) 關于寰椎橋出現率是否存在性別差異,尚無定論的文獻資料。有資料顯示人類的側橋不存在性別差異。對于背橋的研究,不同學者的研究結果不同[3-5]。5) 全面研究靈長類動物的寰椎形態特征,可以讓我們得到更多有價值的信息。例如,人類寰椎背橋的出現很可能是一種返祖現象,因為這種結構在低等的原始靈長類動物的寰椎上普遍存在;另外,通過寰椎一些變量可以計算出體型大小指數,可用于推算靈長類動物的體型大小,還可以計算出相應的形狀指數,去推測不同靈長類動物的形狀差異,包括行為和運動類型[8]。最后,本文作為一項基礎性研究,可以為國內同行進行形態比較和種間比較,為考古工作者鑒定靈長類動物化石等提供相關參考資料[14]。
參考文獻
[1] 歐海寧,黃志明,劉向華,等.寰椎后橋X 線表現及臨床意義[J].現代醫用影像學,2018, 27(3): 705-709
[2] 孫靜宜,張瓊珍,葉靜.寰椎椎動脈溝環與頸性眩暈[J].中華外科雜志,1990, 28 (10): 592-594
[3] 薛德明,趙曉進,郭玫,等.太行獼猴寰椎的測量及相關性研究[J].河南師范大學學報,1996, 24(4): 101-103
[4] 薛德明.太行山獼猴寰椎和樞椎的初步研究[J].動物學雜志,2003, 38(2): 74-76
[5] 肖洪文,吳國雄,張西北,等.寰椎變異的觀測I 中國人寰椎后橋和側橋的觀測[J].瀘州醫學院學報,1990, 12(6): 416-422
[6] Le Mino JM, Trost O. Bony bridge of the atlas (C1) over the groove for the vertebral artery in humans and primates: Polymorphism
and evolutionary trends[J]. American Journal of Physical Anthropology, 2004, 125: 16-29
[7] Yamamoto A, Kunimatsu Y. Geographic variation of atlas bridging of Japanese monkey (Macaca fuscata) [J]. Primate Research,
2002, 18: 215-224
[8] Taitz C, Nathan H. Some observations on the posterior and lateral bridge of the atlas[J]. Acta Anatomica, 1986, 127: 212-217
[9] 趙曉進,王鳳產,田華香,等.獼猴掌骨和指骨長度性差[J].解剖學報,2017, 46(5): 673-678
[10] Barrio PA, Trancho GJ, Sanchez JA. Metacarpal sexual determination in a Spanish population[J]. Journal of Forensic Sciences,
2006, 51(5): 990-994
[11] Kikuchi Y, Nakano Y, Nakatsukasa M, et al. Functional morphology and anatomy of cervical vertebrae in Nacholapithecus kerioi, a
middle miocene hominoid from Kenya[J]. Journal of Human Evolution, 2012, 62: 677-695
[12] 趙曉進,韓霄帆,王鳳產.獼猴胸骨柄性差與鎖骨、股骨長相關性分析[J].解剖學報,2019, 50(2): 123-126
[13] Huxley TH.人類在自然界的位置[M].蔡重陽,王鑫,傅強(譯).北京:北京大學出版社,2010, 41-42
[14] Yamamoto A, Kunimatsu Y. Ontogenetic change and geographical variation of atlas bridging in Japanese Macaques (Macaca
fuscata) [J]. Anthropological Science, 2006, 114: 153-160
[15] Finnegan M. Non-metric variation of the intracranial skeleton[J]. Journal of Anatomy, 1978, 125: 23-37
