miRNA-155靶向SOCS5對無乳鏈球菌誘導羅非魚腦星形膠質細胞炎癥的影響
2023-05-06 09:17:02簡紀常
廣東海洋大學學報 2023年2期
關鍵詞:檢測
張 祥,黃 瑜,蔡 佳,簡紀常,王 蓓
(廣東海洋大學水產學院/廣東省水產動物病害防控與健康養殖重點實驗室/水產經濟動物病害控制廣東省普通高等學校重點實驗室,廣東 湛江 524088)
無乳鏈球菌(Streptococcus agalactis)屬革蘭陽性菌,是一種人、畜及水產動物共患的致病菌,近年在羅非魚養殖中呈高暴發態勢,造成巨大的經濟損失[1,2]。腦是無乳鏈球菌感染的一個重要靶器官[3],感染機制是無乳鏈球菌通過黏附吞噬細胞逃避免疫,后隨巨噬細胞游走侵染其他組織,并透過血-腦屏障引發腦膜炎破壞腦-神經系統,最終由于過度炎癥反應,魚體免疫系統崩潰[4]。
微小核糖核酸155(microRNA-155,miR-155)是由23 個核苷酸組成的一組單鏈小分子RNA,在基因表達的轉錄后水平上起著重要的調節作用[5],其在炎癥反應的發生發展過程中具有重要意義。研究發現,miRNA-155通過與靶mRNA的3'-非翻譯區(3'-untranslated region,3'-UTR)堿基互補配對,阻礙靶mRNA的翻譯或導致其降解[6],miR-155與炎癥反應等病理過程息息相關[7,8]。本課題組前期工作發現,感染無乳鏈球菌后包括miR-155 在內的多個miRNA 在尼羅羅非魚(Oreochromis niloticus)腦及免疫器官中呈顯著的時間依賴性表達,這進一步說明miR-155 的表達水平與炎癥反應密切相關[9]。目前關于miR-155 與靶基因互作調控機體炎癥反應的報道多集中于哺乳動物中,如在小鼠的特應性皮炎和人體的潰瘍性結腸炎中均有miR-155參與并發揮作用[10,11]。細胞因子信號轉導抑制因子5(Suppressor of cytokine signaling 5,SOCS5)屬于免疫抑制分子家族,是一類調控細胞因子信號通路的負反饋調節劑,可以被多種細胞因子誘導而產生,又可反過來抑制眾多促炎癥因子[12,13]。如TNF-α、IL-6等是JAK/STAT 通路上重要的負性調節因素。目前關于SOCS5參與炎癥反應的相關報道主要集中在人、小鼠等哺乳動物中[14,15],關于SOCS5與miRNA 互作參與炎癥反應的機制尚未見報道。
基于此,本研究通過生物信息學分析和qRTPCR、雙熒光素酶報告基因實驗,檢測無乳鏈球菌感染后尼羅羅非魚腦星形膠質細胞中miR-155 和SOCS5基因表達模式,預測miR-155 和SOCS5的靶結合位點,構建尼羅羅非魚SOCS5基因野生型和突變型報告質粒,驗證miR-155和SOCS5的靶向關系,通過進一步過表達和敲降miR-155,檢測miR-155對SOCS5基因表達的影響和調控模式,為后續深入研究miR-155靶向SOCS5在無乳鏈球菌誘導機體炎癥反應中的作用機制提供理論依據。
1 材料與方法
1.1 材料
1.1.1 實驗細胞及菌株 尼羅羅非魚星形膠質細胞系TA-02 由本課題組分離并建系成功[16]。細胞在含體積分數10%的胎牛血清、體積分數1%青鏈霉素混合液的L15 培養基中培養。HEK-293T 細胞系由廣東省水產動物病害防控與健康養殖重點實驗室保存。細胞在含體積分數10%胎牛血清、體積分數1%青鏈霉素混合液的DMEM 培養基中培養。無乳鏈球菌ZQ0910 株由廣東省水產動物病害防控與健康養殖重點實驗室保存[17]。
1.1.2 主要試劑 pGL3-Basic、PRL-TK 雙熒光素酶質粒(本實驗室保存);雙熒光素酶報告基因檢測試劑盒(碧云天生物技術公司);RNA 提取(TransZol Up Plus RNA Kit)、cDNA合成(TransScript One-Step gDNA Removal and cDNA Synthesis Super Mix)及熒光定量(TransStart Green qPCR Super Mix)試劑(北京全式金生物公司);DMEM高糖培養基和胎牛血清(Gibico);LipofectamineTM3000(賽默飛世爾科技);Endo-free Plasmid Mini Kit I質粒小提試劑盒、Universal DNA Purification Kit DNA 純化回收試劑盒(Omega Bio-Tek);KpnI、SmaI 內切酶(寶日生物技術(北京)有限公司);所有引物和尼羅羅非魚miR-155 mimics、miR-155 inhibitor 及其mimics NC、inhibitor NC 均購自生工生物工程(上海)股份有限公司。
引物由生工生物工程(上海)有限公司合成,見表1。

表1 引物序列Table 1 Primer sequence
1.2 方法
1.2.1 無乳鏈球菌感染TA-02 細胞實驗 將無乳鏈球菌ZQ0910 菌株和培養基按體積比1∶100 的比例接種到BHI 液體培養基中進行活化、擴大培養,37 ℃震蕩培養至飽和,離心收集菌體,用1×磷酸鹽緩沖液(PBS,pH 7.3±0.1)沉淀菌體,重復兩次將培養液清洗干凈,用PBS重懸菌體至終濃度為1×107cfu/mL 備用。取生長狀態良好的TA-02 細胞,消化,計數,接種于6 孔細胞培養板中,細胞密度達80%時。每孔加入5 μL無乳鏈球菌菌懸液與TA-02細胞共孵育,分別在0、2、4、6 h收集細胞。吸去培養液,用PBS(pH 7.3 ± 0.1)洗滌2次,每孔加1 mL Trizol Reagent混勻,室溫放置5 min,轉移到無RNase 的EP 管中,-80 ℃凍存,待同一批各個時間點收集完畢后一并提取RNA。
1.2.2 TA-02 細胞RNA 提取與反轉錄 根據TransZol Up Plus RNA Kit 說明書提取羅非魚腦星形膠質細胞RNA。取1 μL 的RNA 檢測其濃度,瓊脂凝膠電泳檢測RNA 的質量。按照TransScript One-Step gDNA Removal and cDNA Synthesis Super Mix說明書將提取的羅非魚腦星形膠質細胞總RNA 反轉成cDNA。
通過NCBI 設計尼羅羅非魚SOCS5定量引物,從miRNA Base 數據庫(http://www.mirbase.org)獲取尼羅羅非魚miR-155 成熟體序列,莖環法合成miR-155 莖環引物和定量引物。miRNA 須加入特異莖環引物進行逆轉錄為模板,miRNA 逆轉錄方法參考廖寧馨[18]的方法進行。
1.2.3 無乳鏈球菌誘導TA-02 細胞中miR-155、炎癥因子和SOCS5基因表達量檢測 通過羅氏LightCycler96實時熒光定量PCR 儀檢測感染無乳鏈球菌的TA-02中miR-155和SOCS5基因的相對表達量,觀察其隨時間變化而變化的規律。以TA-02 細胞cDNA為模板,qPCR 反應體系參考TransStart Green qPCR Super Mix 說明書進行,反應程序為:95 ℃預熱60 s;95 ℃10 s、60 ℃20 s、72 ℃20 s,40 個循環。檢測SOCS5基因和miR-155 的相對表達量,以β-actin 為內參基因,每組設3 個重復,用2-ΔΔCT法計算結果。
1.2.4 生物信息學分析 從miRNA Base 數據庫(http://www.Mirbase.org)獲得尼羅羅非魚miR-155成熟體序列,對比在各個物種間的保守性,利用生物信息學方法(TargetScan、PicTar、miRanda)分析預測SOCS5與miR-155 的靶向關系及靶位點,NCBI數據庫(https://www.ncbi.nlm.nih.gov/)獲取尼羅羅非魚SOCS5基因3'UTR序列。
1.2.5 引物設計與PCR 擴增 根據NCBI 中查找的尼羅羅非魚SOCS5基因3'UTR 序列設計PCR 擴增引物,在序列兩端添加KpnI、SmaI 酶切位點,克隆得到野生型序列(WT)。同時對SOCS5結合位點堿基進行突變,使用搭橋法(重疊PCR)設計SOCS5基因PCR 擴增引物,克隆得到突變型序列(mut)。并根據SOCS5基因3'UTR 序列設計定量引物(包含結合位點),送生工生物工程(上海)有限公司合成,擴增片段大小為533 bp。以TA-02細胞cDNA為模板,擴增SOCS53'UTR 序列,PCR 擴增體系:(總體積20 μL)Extaq 酶10 μL,上下游引物(33 μg/mL)添加各1 μL,模板1 μL,ddH2O 7 μL。擴增程序:95 ℃預變性3 min;95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸20 s,共34個循環;72 ℃再延伸5 min。PCR 擴增產物進行質量分數1%瓊脂糖電泳分析。
1.2.6 產物純化與酶切 根據Universal DNA Purification Kit DNA 純化回收試劑盒說明書進行產物純化,用限制性內切酶KpnI、SmaI分別對純化產物和pGL3-Basic 質粒進行雙酶切,對酶切產物進行回收純化。酶切體系(50 μL):載體和SOCS5純化產物各1 000 ng/μL,KpnI、SmaI 各2 μL,10×Buffer 5 μL,ddH2O 補足50 μL。37 ℃酶切30 min。酶切后的載體和SOCS5基因片段純化步驟同產物純化。
1.2.7 連接轉化和測序鑒定 根據T4 DNA Ligase說明書將酶切純化后的目的片段與pGL3-Basic載體進行連接,產物轉入DH5α 感受態細胞,加入LB(無抗生素)培養基,37 ℃、150 r/min 震蕩培養1~2 h。離心后留100 μL 菌液混勻沉淀,涂于含100 μg/mL Amp 抗生素的LB 固體瓊脂培養基上,37 ℃倒置培養12~16 h。挑取單菌落,進行菌落PCR。對陽性克隆進行測序,序列比對分析正確后,提取質粒保存,質粒提取根據Endo-free Plasmid Mini Kit I 質粒小提試劑盒說明進行。
2011年,在立足博愛竹林資源現狀的基礎上提出的博愛竹林發展目標、總體布局和保護利用等重點工程,通過了我國首個國家級專家評審的縣級產業規劃——《博愛縣竹林資源保護利用總體規劃》,對于保護利用竹林稀缺資源、弘揚竹產業文化具有重要意義[29]。
1.2.8 細胞轉染與雙熒光素酶活性檢測 對數生長期的HEK-293T細胞在細胞密度達到80%左右時接種到24 孔細胞培養板中,每孔加入500 μL 在37 ℃(含體積分數5% CO2)的細胞培養箱培養(濃度為1×106mL-1)。細胞匯合度70%~90%時進行轉染。轉染方法和用量按照Lipofectamine?3000轉染說明書進行。實驗共分為6組:pGL3-Basic-NC+miR-155 mimics-NC 組、pGL3-Basic-NC+miR-155 mimics組、SOCS5-3'UTR-WT+miR-155 mimics-NC 組、SOCS5-3'UTR-WT+miR-155 mimics 組、SOCS5-3'UTR-mut+miR-155 mimics-NC 組、SOCS5-3'UTRmut+miR-155 mimics組,每組設3個復孔,每孔總體積50 μL。6 h 后,更換含血清的新鮮培養基繼續培養,48 h 后收集細胞培養物。轉染48 h 后吸出培養基,根據雙熒光素酶報告基因檢測試劑盒說明書檢測各組熒光素酶活性。
1.2.9 miR-155 對SOCS5 表達量的影響 將miR-155 mimics、mimics NC、miR-155 inhibitor、inhibitor NC 分別轉染HEK-293T 細胞,48 h 后收集細胞樣本,進行RNA 提取和反轉錄,熒光定量PCR 檢測miR-155 的表達量,評估mimics 及inhibitor 的效果。將miR-155 mimics、mimics NC、miR-155 inhibitor、inhibitor NC 分別和SOCS5共轉染HEK-293T 細胞,48 h 后收集細胞樣本,進行RNA 提取和反轉錄,利用熒光定量PCR 檢測SOCS5基因的相對表達量。以甘油醛-3-磷酸脫氫酶(GAPDH)為內參,每組設置3 個重復,用2-ΔΔCT方法計算結果。
1.2.10 質粒和miR-155 inhibitor 共轉染HEK-293T細胞 將miR-155 mimics 和miR-155 inhibitor 以及SOCS5野生型質粒共轉染HEK-293T 細胞。miR-155 mimics/inhibitor轉染濃度為20 μmol/μL,每孔以體積比1:1 的比例將miR-155 mimics/inhibitor 與SOCS5-WT-3'UTR 質粒共轉染HEK-293T 細胞,共轉染miR-155 mimics 和SOCS5-WT-3'UTR 質粒做對照。分別在轉染后6 h 和12 h 收集細胞樣本,實時熒光定量PCR檢測miR-155和SOCS5表達量。
1.2.11 HEK-293T 細胞RNA 提取與反轉錄 吸去細胞培養液,PBS(pH 7.3±0.1)洗滌2 次,加入1 mLTrizol Reagent,充分混勻,室溫放置5 min,轉移到無RNase 的EP 管中,-80 ℃凍存,待同一批各個時間點樣品收集完畢后一并提取RNA,反轉錄后cDNA放-20 ℃保存。
1.2.12 轉染miR-155 inhibitor 后尼羅羅非魚miR-155 和SOCS5 表達量檢測 通過羅氏LightCycler96實時熒光定量PCR 儀檢測共轉染miR-155 inhibitor后HEK-293T 細胞中miR-155 和SOCS5基因的相對表達量。以上述反轉錄的HEK-293T 細胞cDNA 為模板,檢測SOCS5和miR-155 的相對表達量,以GAPDH 為內參,每組設置3 個重復,用2-ΔΔCT方法計算結果。
1.3 數據處理與分析
每組實驗進行3 次重復,數據采用SPSS 23.0 分析數據,結果用均值± 標準差表示。采用單因素方差分析進行多組間比較,采用t檢驗進行組間比較,以P<0.05 作為差異顯著的標準,使用GraphPad Prism 8.0.1軟件作圖。
2 結果
2.1 無乳鏈球菌感染對TA-02 細胞中miR-155 和SOCS5基因表達的影響
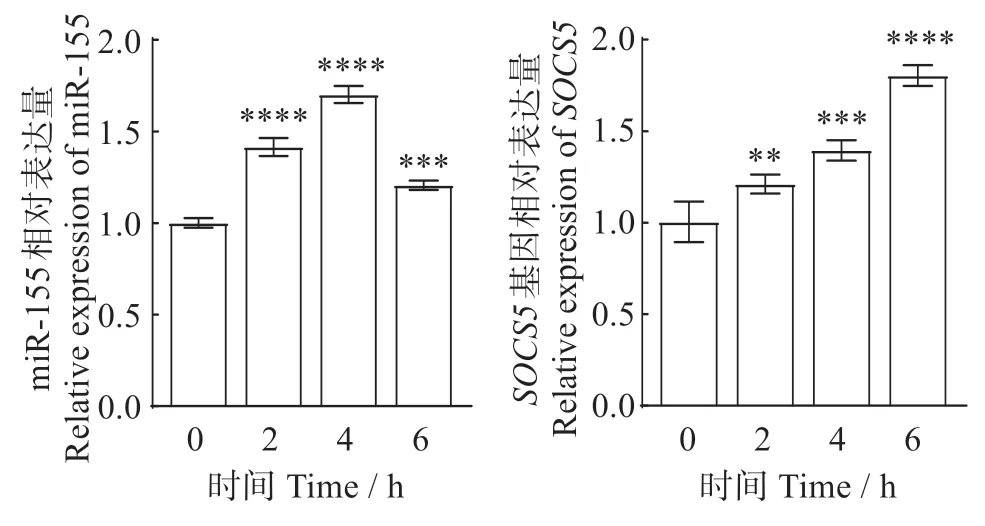
圖1 無乳鏈球菌感染TA-02細胞中的miR-155和SOCS5表達Fig.1 Expression of miR-155 and SOCS5 in TA-02 cells infected with Streptococcus agalactiae
2.2 無乳鏈球菌感染對TA-02細胞中炎癥因子表達的影響
無乳鏈球菌ZQ0910感染TA-02細胞0、2、4、6 h,實時熒光定量PCR 檢測炎癥因子表達量如圖2 所示。隨著感染時間增長,促炎癥因子IL-1β和TNF-α表達量與對照組相比逐漸升高,IL-1β在2、4 和6 h分別升高0.74、0.62和0.39倍,TNF-α分別升高0.04、0.51 和0.42 倍。而抑炎癥因子TGF-β和IL-10與對照組相比逐漸降低,TGF-β在2、4 和6 h 分別降低0.06、0.10 和0.40 倍,IL-10在2、4 和6 h 分別降低0.22、0.30和0.52倍。
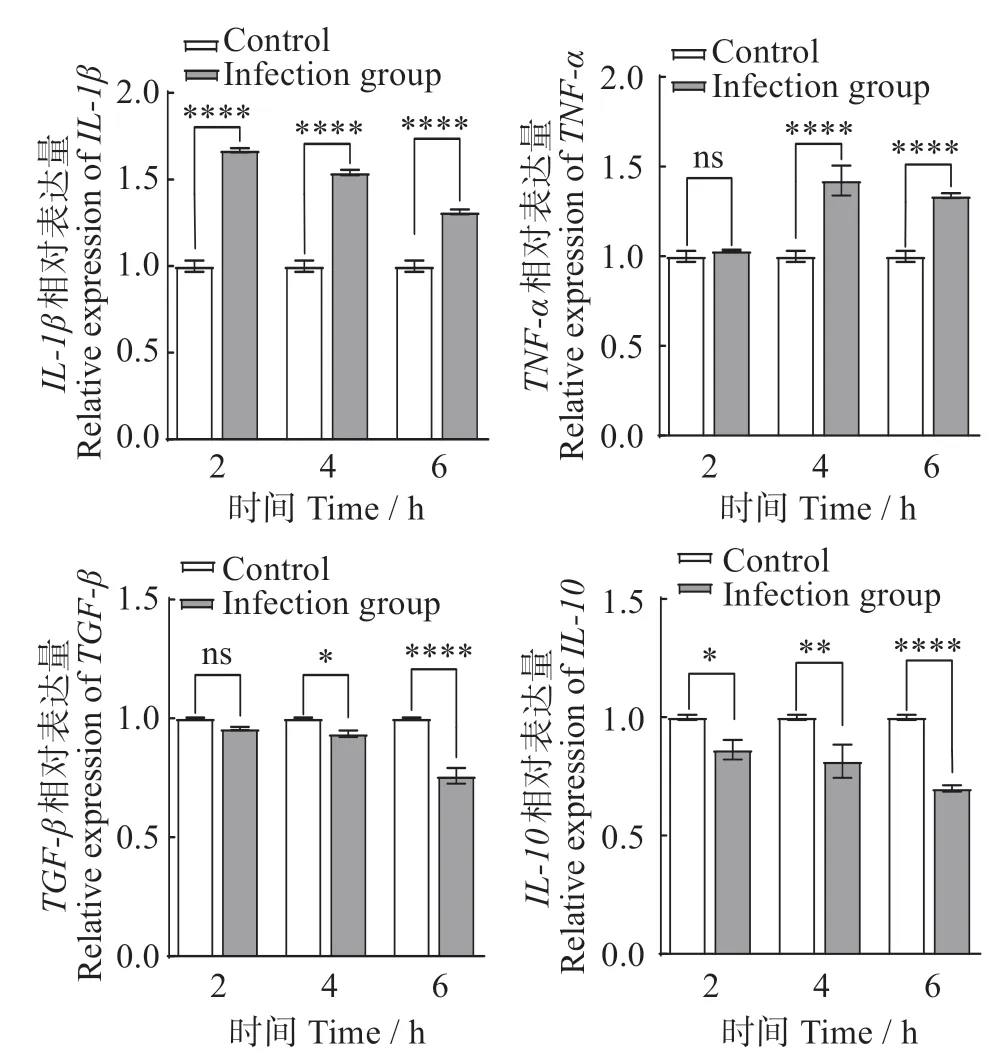
圖2 無乳鏈球菌感染TA-02細胞中的炎癥因子表達Fig.2 Expression of inflammatory cytokines in TA-02 cells infected with Streptococcus agalactiae
2.3 miR-155和SOCS5靶向結合位點預測
通過比對證明miR-155成熟體序列在包括人在內的多個物種間具有高度保守性,通過TargetScan等數據庫查詢到miR-155與SOCS5存在堿基互補序列,具有一個潛在結合位點(圖3),這提示SOCS5可能是miR-155的靶基因。
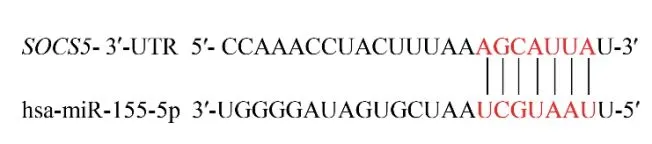
圖3 miR-155-5p與SOCS5結合位點預測Fig.3 Prediction of binding sites between miR-155-5p and SOCS5
2.4 PCR擴增與酶切產物鑒定
PCR 擴增尼羅羅非魚SOCS5野生型和突變型目的片段長度應為533 bp,瓊脂糖凝膠電泳檢測表明經酶切純化后的SOCS5野生型和突變型片段大小與預期片段長度一致,為533 bp,將目的片段構建至pGL3-Basic載體中,挑取單菌落進行菌落PCR 鑒定,將符合目標片段大小的樣品送生工進行測序分析,測序結果與預期片段一致(圖4)。
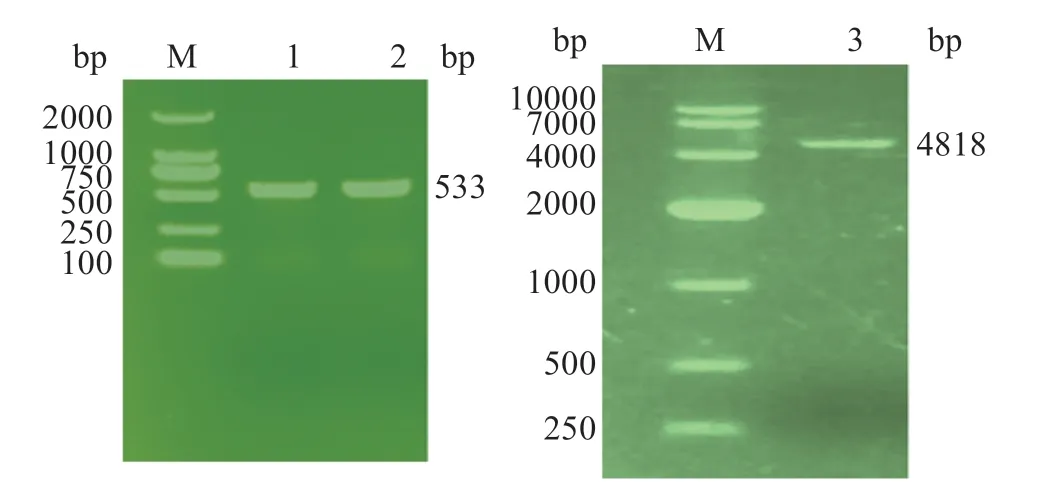
圖4 SOCS5基因3'UTR區和pGL3-Basic載體酶切純化產物電泳條帶Fig.4 Electrophoretic map of the 3'UTR region of SOCS5 gene and the purified product digested by pGL3-Basic vector
2.5 測序分析
對含有尼羅羅非魚SOCS5基因結合位點的野生型序列及缺失尼羅羅非魚SOCS5基因結合位點的突變型序列進行克隆測序,尼羅羅非魚SOCS5-WT 載體和SOCS5-mut 載體經測序驗證均構建成功,成功將靶序列TGCATTA 突變為CTAGCGC,可用于后續靶點驗證實驗(圖5)。
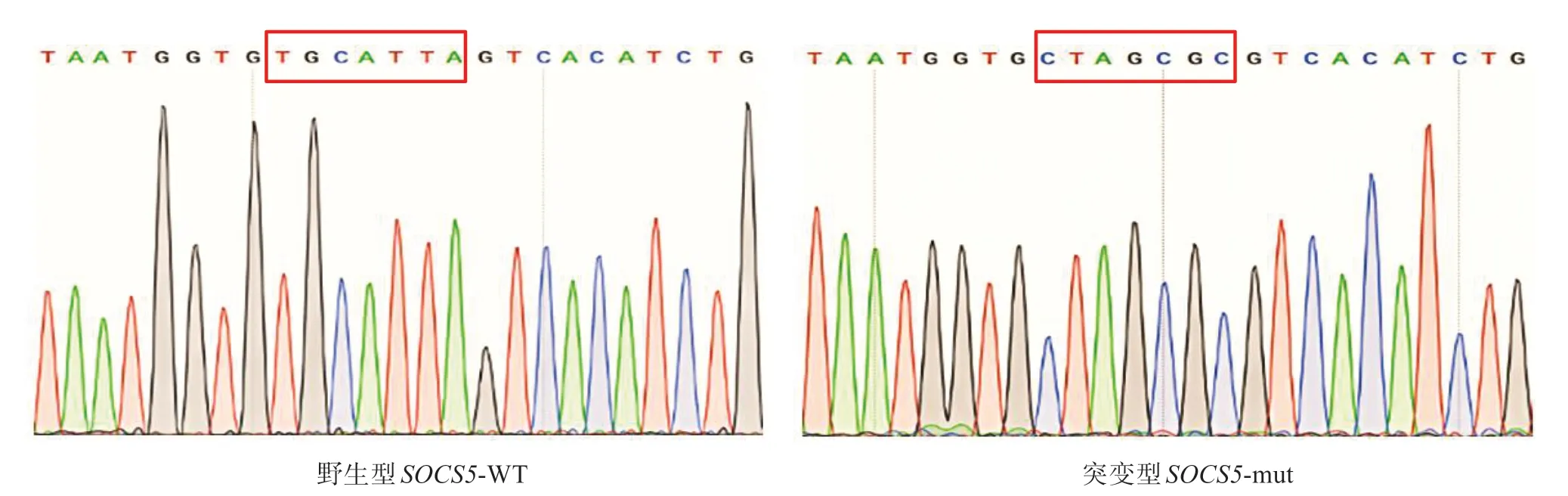
圖5 SOCS5野生型與突變型重組質粒部分測序結果Fig.5 Partial sequencing of SOCS5 wild type and mutant recombinant plasmid
2.6 miR-155對SOCS5的調控作用
雙熒光素酶實驗結果表明(圖6),共轉染miR-155 模擬體和野生型報告質粒后,與轉染的miRNA的陰性對照相比,miR-155 模擬物可極顯著(P<0.000 1)抑制其活性,抑制效果達72.9%。miR-155模擬體在和突變型報告質粒共轉染的情況下,miR-155模擬體抑制作用顯著(P<0.000 1)減弱,與野生型組相比減弱53.7%。以上結果初步表明miR-155可以靶向調控尼羅羅非魚SOCS5基因。

圖6 各組轉染后細胞的熒光素酶表達Fig.6 Luciferase expression of cells after transfection in each group
2.7 miR-155對SOCS5表達量的影響
如圖7所示,與mimics NC 組相比,miR-155 mimics 極顯著(P<0.000 1)的提高miR-155 表達量,升高5.24倍。與inhibitor相比,miR-155 inhibitor極顯著(P<0.01)的抑制miR-155 的表達,降低2.05倍。
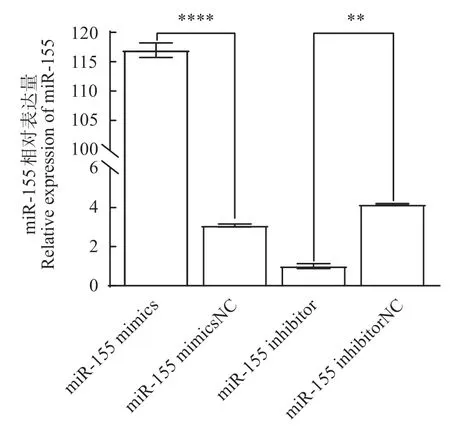
圖7 miR-155相對表達量Fig.7 Relative expression of miR-155
miR-155 mimics、mimics NC、miR-155 inhibitor、inhibitor NC 分別和SOCS5-WT-3'UTR 共轉染HEK-293T 細胞,實時熒光定量PCR 檢測尼羅羅非魚SOCS5基因表達量,實驗結果如圖8 所示。與mimics NC組相比,miR-155 mimics組SOCS5的表達量極顯著(P<0.001)下調,降低0.93 倍,表明miR-155 能顯著抑制尼羅羅非魚SOCS5基因的表達。而加入miR-155 inhibitor 后,SOCS5的表達量顯著(P<0.05)上調,升高0.62倍,證明miR-155 inhibitor能抑制miR-155的表達,從而降低對SOCS5的調控作用。
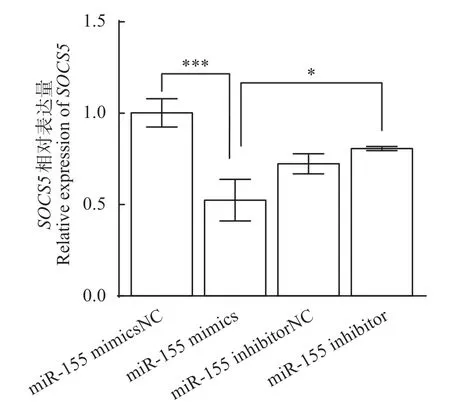
圖8 SOCS5基因相對表達量Fig.8 Relative expression of SOCS5 gene
2.8 轉染miR-155 inhibitor 后尼羅羅非魚miR-155對SOCS5的調控作用
miR-155 mimics和miR-155 inhibitor以及SOCS5野生型質粒共轉染HEK-293T 細胞,實時熒光定量PCR 結果表明,與對照組相比,實驗組在6、12 h 時極顯著降低了miR-155 的表達量(P<0.01),分別降低0.43 和0.64 倍,表明轉染miR-155 inhibitor 后,能成功抑制miR-155 表達。而SOCS5表達量在6 h 時顯著上調(P<0.05),12 h 時表達量極顯著上調(P<0.000 1),分別升高0.42和2.47倍(圖9)。

圖9 轉染miR-155 inhibitor后尼羅羅非魚中miR-155和SOCS5基因相對表達量Fig.9 Relative expression of miR-155 and SOCS5 genes in Oreochromis niloticus after miR-155 inhibitor transfection
3 討論
無乳鏈球菌感染羅非魚可以誘導大量炎癥反應的發生,導致急性死亡從而增加致病性[19,20],其中腦是無乳鏈球菌感染的一個重要靶器官,當無乳鏈球菌感染細胞后,細胞的模式識別受體與病原體相關分子模式結合,啟動固有免疫應答[21,22]。因此本實驗以TA-02 細胞為實驗模型,探究無乳鏈球菌感染尼羅羅非魚腦細胞誘發炎癥反應的炎癥水平變化及miR-155靶向SOCS5基因參與機體炎癥反應的調控機制。目前無乳鏈球菌感染羅非魚主要有兩種途徑:一種是通過在宿主血液中增殖后侵染其他組織引發病變;另一種是透過血-腦屏障,侵入羅非魚腦組織誘發腦膜炎,進而引發一系列生理和病理變化[23,24]。本研究通過無乳鏈球菌感染尼羅羅非魚腦星形膠質細胞實驗發現,miR-155 在不同時間點的表達上調,證明在清除無乳鏈球菌病原菌的免疫反應中,miR-155 確實參與了尼羅羅非魚腦星形膠質細胞的免疫反應。同時在無乳鏈球菌誘導的腦星形膠質細胞中,檢測到炎癥因子IL-1β、TNF-α的釋放,并且這兩種炎癥因子的表達模式與miR-155一致,其表達量都是隨著時間增加。這說明在應對無乳鏈球菌入侵時,miR-155 是起到保護機體、對抗其入侵的作用。同時檢測到IL-10、TGF-β的表達量降低,這與陳秀慧等[25]關于子宮內膜炎研究中對IL-10的表達水平檢測結論一致;另趙瑾[26]研究證明經治療后的膝骨性關節炎患者體內TGF-β表達水平下調,說明IL-10、TGF-β的下調可以適量減輕炎癥反應,使炎癥水平維持在一個適度范圍內。
研究發現,miRNA主要通過與靶基因mRNA的3'-UTR堿基互補配對,來阻礙靶基因mRNA 翻譯成蛋白質或導致其降解,即參與基因轉錄后的表達,實現對靶基因的調控作用[27,28]。已有研究證明miR-155 與炎癥反應密切相關,在機體先天性免疫和獲得性免疫中均發揮重要調控作用[29,30],廣泛參與炎癥反應等多種生理活動,因此對靶基因預測和相互作用關系掌握是了解miR-155參與機體炎癥反應的前提[31]。目前關于miR-155與SOCS5靶向關系的研究報道較少,石瑜等[32]通過實驗證明miR-155-5p 靶向調控SOCS5促進大鼠(Rattus norvegicus)腦缺血-再灌注損傷。本研究通過生物信息學方法成功預測尼羅羅非魚miR-155與SOCS5之間存在一個靶位點,并利用雙熒光素酶報告基因實驗驗證發現miR-155 能極顯著抑制SOCS5基因的表達,但突變結合位點后,miR-155 對SOCS5基因的抑制能力顯著減弱。這證明SOCS5是miR-155 的靶基因。將miR-155 mimics、mimics NC、miR-155 inhibitor、inhibitor NC 分別轉染HEK-293T 細胞過表達miR-155 后,檢測到SOCS5表達量與對照組相比下降0.93 倍,而敲降miR-155后,SOCS5表達量上調0.62倍,證明miR-155 能靶向調控SOCS5的表達,結合雙熒光素酶報告基因實驗進一步說明miR-155可以通過與SOCS5基因3'UTR特異性結合,從而抑制SOCS5基因表達。
SOCS5屬于免疫抑制分子家族(包括SOCS1-7和CIS),是由細胞因子誘導產生并反饋性調控細胞因子信號傳遞過程的負調節因子[33]。SOCS 家族基因的異常表達與機體炎性疾病及惡行腫瘤的發生發展密切相關[34,35],已有研究證明SOCS5在機體癌癥等疾病中發揮重要作用[36,37],而癌癥發展早期都伴有炎癥反應的發生,二者有潛在聯系,這提示未來SOCS5可能作為癌癥診斷和治療的切入點。陳景行等[15]發現SOCS5在小鼠氣道炎癥反應中發揮重要作用。本研究在無乳鏈球菌感染的TA-02細胞中檢測到SOCS5表達量升高,提示SOCS5參與了TA-02 細胞炎癥反應,有研究表明,SOCS5是一種對細胞因子調節通路有負性調節作用的因子,能有效抑制炎癥因子表達[38]。這說明在信號通路中可能存在某種抑制SOCS5過度激活的機制,正是這種機制的協同作用,才避免在病原菌感染后產生的炎癥反應初期,miR-155過度調控SOCS5表達,抑制抗炎因子的產生,從而導致免疫失衡。本研究與廖寧馨[18]關于新生隱球菌(Filobasidiella bacillispora)介導的巨噬細胞炎癥反應中SOCS1的表達模式相同。
本研究通過轉染miR-155 的inhibitor 抑制尼羅羅非魚腦細胞中miR-155 的表達,觀察miR-155 對靶基因SOCS5的調控作用,在轉染miR-155 inhibitor后,成功抑制了miR-155 的表達,相對應時間點SOCS5表達量升高,說明在敲降miR-155 表達后能部分解除對靶基因SOCS5的抑制作用。miRNA 調控靶基因參與機體生命活動是一個較為復雜的過程,研究表明[39,40],SOCS5是JAK-STAT 通路中起關鍵作用的負調控因子,而JAK-STAT 通路參與了多種與中樞神經系統疾病相關的炎癥反應,因此推測miR-155 可能通過調控SOCS5來調節下游JAKSTAT 信號通路。張琴和張賀[39,40]研究中關于SOCS5參與機體炎癥反應的報道與本研究中SOCS5的研究結果一致,這說明在無乳鏈球菌介導的TA-02 細胞炎癥反應的早期階段,正是由于miR-155 對SOCS5的這種負向調控機制,才將炎癥水平控制在在一個合理范圍內,從而避免過度抑制炎癥因子產生使免疫系統損傷,不能有效清除病原體。
綜上所述,本研究證實miR-155 與靶基因SOCS5在無乳鏈球菌誘導的TA-02細胞炎癥反應中發揮重要作用,且miR-155 負向調控SOCS5參與炎癥發應的發生發展。目前關于miRNA 與靶基因互作參與機體炎癥反應的研究多集中于哺乳動物中[41,42],在魚類中的對其調控機制的研究處于初級階段。但miR-155靶向調控SOCS5的蛋白表達機制及miR-155靶向SOCS5與相關信號通路的關系還有待進一步研究。
4 結論
本研究證實miR-155和SOCS5參與了無乳鏈球菌介導的TA-02細胞炎癥反應。通過生物信息學方法獲得尼羅羅非魚miR-155和SOCS5基因的靶向結合位點,并成功構建尼羅羅非魚SOCS5基因3'-UTR雙熒光素酶報告基因野生型和突變型載體,證明SOCS5基因是miR-155 直接作用的靶基因,且miR-155 結合于SOCS5基因的3'-UTR 區域,在轉錄后水平對SOCS5有直接抑制作用。通過敲降尼羅羅非魚miR-155 表達,證明miR-155 可以靶向負調控SOCS5表達,從而影響炎癥變化的發生發展。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
