凡納濱對蝦過氧化物還原酶3基因的分子克隆與功能分析
2023-05-06 09:17:04張秀霞魯耀鵬張澤龍李軍濤鄭佩華冼健安
廣東海洋大學學報 2023年2期
郭 慧,李 騰,,張秀霞,魯耀鵬,張澤龍,李軍濤,鄭佩華,冼健安
(1.廣東海洋大學水產學院,湛江市海洋生態與養殖環境重點實驗室,廣東 湛江 524088;2.中國熱帶農業科學院熱帶生物技術研究所,海南省海洋生物資源功能性成分研究與利用重點實驗室/海南熱帶農業資源研究院海南省熱帶農業生物資源保護與利用重點實驗室,海南 海口 571101;3.中國熱帶農業科學院湛江實驗站,廣東 湛江 524013)
過氧化物還原酶(Peroxiredoxin,Prx)是多數生物的一類多功能抗氧化酶[1],在生物體免疫、發育、炎癥、蛻皮、繁殖和細胞凋亡等過程發揮關鍵作用[2,3]。Prx 由六個亞家族(Prx1~Prx6)構成,通過半胱氨酸(Cys)殘基的數目和保守程度分為三類:Prx1~Prx4,典型的2-Cys Prx;Prx5,非典型2-Cys Prx;Prx6[5-7]。其中,典型2-Cys Prx 是甲殼動物中分布較多的Prx 類型,有較高的催化效應[8],主要通過同源二聚體的方式清除過氧化氫、烷基過氧化氫等過氧化物。
目前,已在脊尾白蝦(Exopalaemon carinicauda)[9]、中國明對蝦(Fenneropenaeus chinensis)[10]、斑節對蝦(Penaeus monodon)[11]和中華絨螯蟹(Eriocheir sinensis)[12]等甲殼動物中獲得Prx基因及其編碼的氨基酸序列。然而,這些研究僅發現Prx基因的其中一個或幾個亞型[13],其表達模式也有一定物種差異和基因亞型差異,蝦蟹類中許多物種的Prx信息還有待挖掘和深入探究[12]。迄今,關于凡納濱對蝦Prx的研究較少,其在凡納濱對蝦遭受病原菌感染及逆境脅迫時的生理功能尚不清楚。
凡納濱對蝦(Litopenaeus vannamei)是我國最重要的養殖蝦種之一。然而,工廠高密度集約化的養殖模式嚴重破壞了對蝦的養殖環境,引起種質資源退化、病害頻發,造成蝦類免疫功能受損及抗應激能力下降,給水產養殖經濟造成較大損失[14-15]。鑒于Prx基因在機體免疫過程中的重要作用,筆者研究其在對蝦抗脅迫和抗病原體免疫防御中的作用,為研究甲殼動物Prx基因的生理功能及作用機制提供參考,并為培育抗病高產對蝦新品種提供理論基礎。
1 材料與方法
1.1 供試對蝦的管理和飼養
凡納濱對蝦購自于海南省文昌市對蝦養殖場,體質量(7.56±0.79)g,體長(9.86±0.28)cm,生長狀況良好。實驗在中國熱帶農業科學院文昌實驗基地進行。所有對蝦暫養在鹽度15、溫度(24±2)℃、pH 7.9~8.0 的循環海水系統中,連續曝氣增氧,并進行循環過濾處理。在實驗前,每天定時定點定量投喂對蝦商業飼料(基本成分質量分數:蛋白質40%,脂肪5%,纖維5%,灰分16%)。
1.2 對蝦總RNA提取及cDNA合成
取健康蝦3尾,剖取肝胰腺,放入液氮中進行研磨,按TRIzol 法提取組織總RNA,利用NanoDrop 2000 分光光度計(Thermo Scientific,USA)檢驗RNA 的純度和質量,并通過瓊脂糖凝膠電泳評估其完整性。選取質量良好的總RNA 用反轉錄試劑盒PrimeScript RT reagent Kit With gDNA Eraser(TaKaRa,大連)反轉錄合成第一鏈cDNA。
1.3 Prx基因cDNA序列克隆
從凡納濱對蝦肝胰腺4-NP 脅迫轉錄組中得到LvPrx3基因的部分序列,設計中間片段的擴增引物Prx-F和Prx-R(表1)。以凡納濱對蝦肝胰腺cDNA為模板,經過第1 輪PCR 擴增,擴增程序:94 ℃3 min;94 ℃30 s,58 ℃30 s,72 ℃1 min,35 個循環;72 ℃10 min。PCR 產物經瓊脂糖凝膠電泳檢驗后,用SanPrep 柱式DNA 膠回收試劑盒(生工生物工程(上海)股份有限公司)回收、純化,連接至pMD18-T 載體(TaKaRa,大連),轉化到大腸埃希菌(Escherichia coli)DH5α 感受態細胞(TaKaRa,大連)中。挑選陽性單克隆質粒,送至深圳華大基因公司測序。
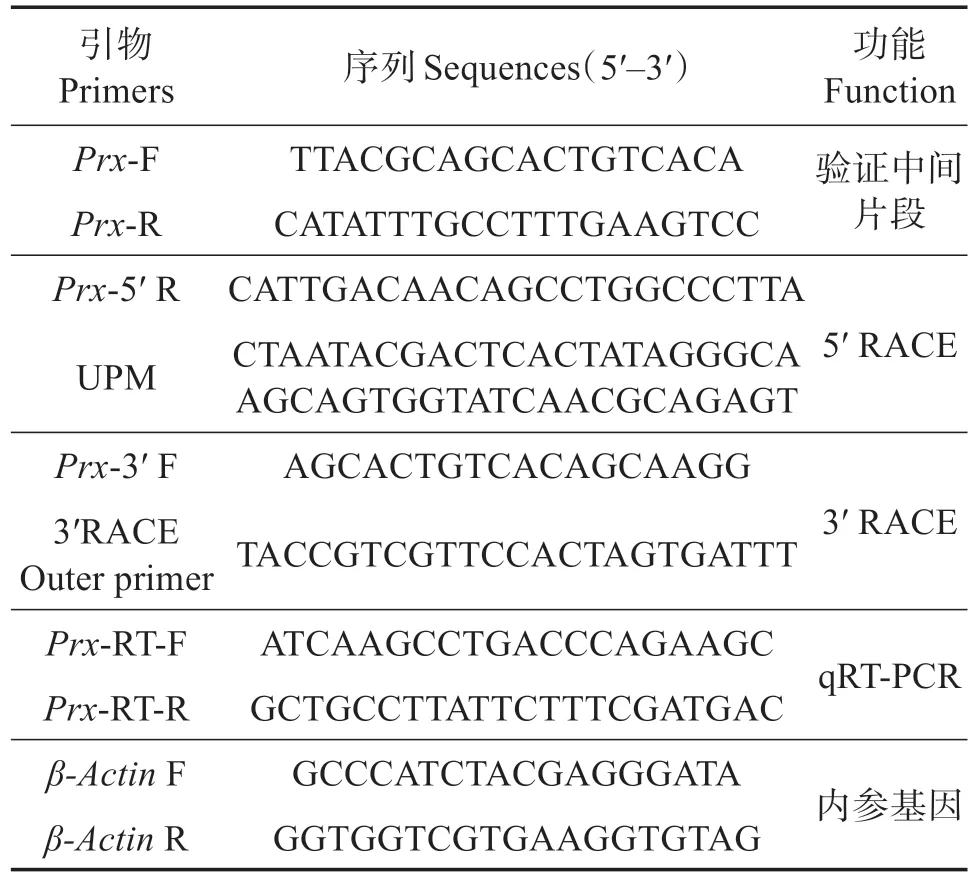
表1 實驗中所用引物序列Table 1 Oligonucleotide primers used in experiments
LvPrx3基因5' 末端片段按照SMART RACE cDNAAmplification Kit試劑盒(Clontech,USA)說明書進行擴增,用Primer Premier 6.0 設計基因下游引物Prx-5'R 和Universal Primer Mix(UPM)進行配對擴增,擴增程序:94 ℃30 s,68 ℃30 s,72 ℃3 min,30 個循環。Prx3'末端,使用3'-Full RACE Core Set with PrimeScript RTase 試劑盒(Clontech,USA),根據已知中間片段設計基因上游引物Prx-3'F,然后與3' RACE Outer Primer 進行PCR 擴增,擴增程序:94 ℃3 min;94 ℃30 s,58 ℃30 s,72 ℃1 min,35個循環;72 ℃10 min。5'-RACE 和3'-RACE 產物經凝膠回收純化,與克隆載體連接、轉化感受態細胞,測序。將目的基因5'和3'端測序結果聯合中間片段進行拼接,取得最終的LvPrx3基因cDNA 全長序列。
1.4 LvPrx3基因序列分析
通過DNAStar 中的EditSeq 軟件分析LvPrx3的開放閱讀框(ORF)及其推導的氨基酸序列,利用NCBI 數據庫 的BLAST(http://www.ncbi.nlm.nih.gov/blast)查找LvPrx3的同源核酸序列及其氨基酸序列并進行比對。用ExPASy(http://au.expasy.org/tools/)程序預測LvPrx3的蛋白理化性質,以SignalP 5.0(http://www.cbs.dtu.dk/services/SignalP/)預測其信號肽,運用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM/)分析蛋白跨膜結構域;Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)預測蛋白的亞細胞定位;再通過NCBI 的保守結構域(CDD)(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)預測其保守區和特殊位點;運用Clustal X 和MEGA 6.0 對不同物種來源的Prx 氨基酸序列進行多重比對及聚類分析,并以鄰接法(NJ)構建系統發育樹(置信度:Bootstrap=1000)。
1.5 LvPrx3基因組織分布特征分析
隨機選取15 尾健康蝦,取其血細胞以及眼柄、鰓、肝胰腺、腸道、肌肉組織,提取其總RNA并進行半定量PCR。以內參基因的β-ActinF和β-ActinR為引物,各組織cDNA為模板進行半定量PCR擴增,程序:94 ℃3 min;94 ℃30 s,58 ℃30 s,72 ℃1 min,30個循環;72 ℃10 min。擴增產物用瓊脂糖凝膠電泳評估。根據條帶亮度調整各組織cDNA模板使用量,使內參基因擴增的片段亮度基本一致,最終確定各組織cDNA用量。在相同狀態下,以Prx-RT-F和Prx-RT-R為引物擴增LvPrx3,產物用瓊脂糖凝膠電泳檢測。
1.6 脂多糖(LPS)注射和4-壬基酚(4-NP)脅迫實驗
將150尾健康對蝦隨機分成LPS組和對照組,每組3 個重復組,每重復組25 尾。將LPS(2 mg·mL-1,E.coliL2880,Sigma)溶解于無菌生理鹽水,配制成2 μg·μL-1的LPS 注 射液。根據Xian 等[16]的研究,LPS 組按每g 蝦8 μg 的用量注射LPS 溶液,對照組注射等量的無菌生理鹽水。在注射后的0、3、6、12、24、48 h 時,實驗組和對照組各隨機采樣9 尾,取其眼柄和腸道在液氮速凍,置于-80 ℃下保存備用。
根據本課題組的前期研究確定4-NP 脅迫質量濃度500 μg·L-1[17]。用4-NP(標準品,純度最低為99%)和無水乙醇制備1 mg·mL-1的4-NP 母液。在海水中加入4-NP 母液,使4-NP 最終質量濃度為500 μg·L-1。對照組中加入等量的無水乙醇。無水乙醇在4-NP實驗組中的最終體積分數小于0.002%。設置實驗組和對照組,每組設3個平行,每個平行組隨機放置健康蝦30 尾。為保證水體的實驗濃度穩定,每24 h換水1次,每次換水量為50%,加入4-NP工作溶液,保持4-NP 最終質量濃度為500 μg·L-1,在水體中的實際質量濃度為(467.18 ± 28.56)μg·L-1。在對照組中未檢測到4-NP。4-NP暴露0、24和48 h后,每個平行分別隨機采集9尾蝦,取其眼柄和腸道在液氮速凍保存。
1.7 LvPrx3基因相對表達量分析
用Bio-Rad T100(Bio-Rad,USA)儀器進行qRTPCR檢測,以Prx-RT-F、Prx-RT-R和β-ActinF、β-ActinR為引物,PCR 反應體系設置為20 μL。擴增程序:94 ℃3 min,95 ℃15 s,58 ℃15 s,72 ℃20 s,40個循環。采用2-ΔΔCt法計算表達量,數據以平均值±標準差表示,用SPSS 19.0進行t檢驗,分析LPS刺激和4-NP脅迫下,LvPrx3基因在相應組織中的表達變化。
2 結果與分析
2.1 LvPrx3基因序列分析
通過5'/3'-RACE 技術分別克隆得到585 bp 的5'末端和682 bp 的3'末端,經拼 接獲得962 bp 的LvPrx3cDNA 全長序列。該序列含有45 bp 的5'端非編碼區(5' UTR)、233 bp 的3' 端非編碼區(3' UTR)和684 bp 的ORF,可編碼227 個氨基酸。其3'末端處有1 個終止密碼子TAA,多聚腺苷酸加尾信號AATAAA 和poly A 尾(圖1)。LvPrx3 蛋白的分子質量為25.09 ku,pI 為6.22。LvPrx3具有“FYPLDFTFVCPTE”的N 端和“GEVCPA”的C 端,為典型2-Cys Prx的特殊結構域。此外,LvPrx3還有以下的特殊結構域:催化三聯體結構(T77-C80-R156),二聚體界面(V37-F78-V79-T82-G145-L155-K168-H169-M170-V172-N173-D174-G178-R179-R186-C201-P202-A203-N204),十聚體(五個二聚體)界面(F76-F110-S111-F136-K138-D150),過氧化和可溶性半胱氨酸(C80-C201)(圖2)。SignalP 5.0預測結果表明,LvPrx3蛋白為跨膜信號肽。Cell-PLoc 2.0預測結果顯示,LvPrx3蛋白主要定位于線粒體中。

圖1 凡納濱對蝦Prx3及其推導的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of Prx3 gene of Litopenaeus vannamei
2.2 LvPrx3基因同源性及進化樹分析
LvPrx3基因與凡納濱對蝦基因組對應核苷酸序列(GenBank 登錄號XR_003477980.1)的相似性達99%;LvPrx3基因可編碼227個氨基酸,而基因組對應核苷酸序列編碼的氨基酸(ROT83170.1)僅為186個,兩者的覆蓋率(Coverage)為72%。LvPrx3與報道的凡納濱對蝦Prx 蛋白序列(ACX53642)的相似性為65%;與Prx4(AET36895)的相似性為57%。同其他物種相比,LvPrx3 與斑節對蝦(P.monodon,XP_037784347.1)的Prx 相似性最高,為98%;與日本對蝦(Penaeus japonicus,XP_042874112.1)和美國龍蝦(Homarus americanus,XP_042239359.1)的相似性次之,分別為94%和86%;與擬穴青蟹(Scylla paramamosain,ASS34530.1)的相似性為82%。與無脊椎動物日本扇貝(Mizuhopecten yessoensis,OWF43851.1)、蚤狀溞(Daphnia pulex,EFX63892.1)和大型溞(Daphnia magna,KZS17058.1)的相似性分別為75%、75%和70%,與紅樹林鳉(Kryptolebias marmoratus,XP_017259545.1)、虎尾海馬(Hippocampus comes,XP_019737549.1)、長鰭叉尾鮰(Ictalurus furcatus,ADO27790.1)、斑點叉尾鮰(I.punctatus,NP_001188127.1)、黃顙魚(Tachysurus fulvidrac,XP_027030673.1)的相似性分別為74%、72%、72%、72%和68%。
多重序列比對結果表明,催化三聯體與過氧化和可溶性Cys結構在所比對物種的Prxs序列中高度保守,但二聚體界面結構和十聚體界面結構在不同物種的序列中有差異。在典型的2-Cys型Prx中,包含兩個保守且有還原活性的Cys:過氧化Cys和可溶性Cys,且為結構域交疊的同源二聚體。可溶性Cys位于C 端,由五個二聚體按特定規則形成一個環狀十聚體[18-20]。由此推斷,本研究克隆的LvPrx3基因屬于典型的2-Cys 型Prx;同時結合Cell-PLoc 2.0 預測結果,該基因定位于線粒體中,屬于Prx3亞型(圖2)。

圖2 LvPrx3蛋白的多序列比對Fig.2 Multiple sequence alignment of LvPrx3 protein
由Prx 序列相似性構建的系統進化樹顯示,凡納濱對蝦先與同為節肢動物門甲殼綱的斑節對蝦聚為一支,再與軟甲綱的擬穴青蟹聚成一小分支;而節肢動物門切甲亞綱的兩種水溞和軟體動物門雙殼綱日本扇貝聚為一小分支后再與凡納濱對蝦所在分支形成無脊椎動物的兩大分支。Prxs的變化趨勢符合從低等無脊椎動物發展到高等脊椎動物的進化歷程(圖3)。
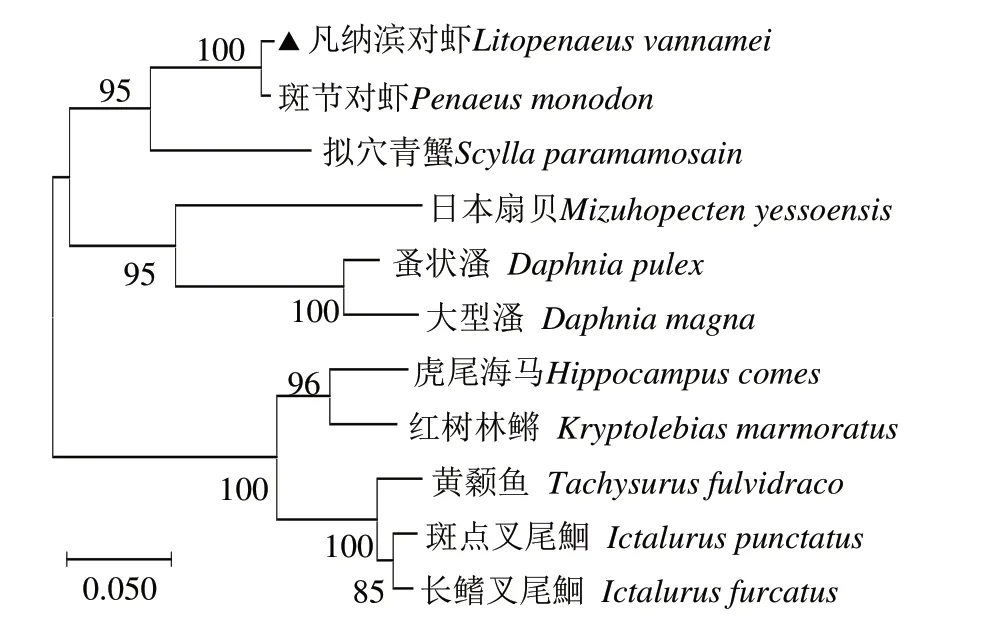
圖3 LvPrx3基因的系統進化分析Fig.3 Phylogenetic analysis of LvPrx3 gene
2.3 LvPrx3基因在不同組織中的表達量
圖4 可見,LvPrx3基因在所有檢測組織中均有不同程度的表達,在眼柄中表達量最高,腸道次之,其他4種組織中的表達量無明顯區別。
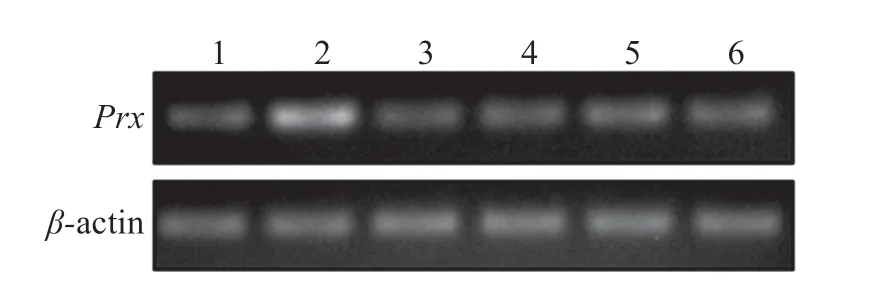
圖4 LvPrx3在不同組織中的相對表達Fig.4 Expression levels of LvPrx3 gene in different tissues
2.4 LPS刺激及4-NP脅迫后LvPrx基因的表達量
如圖5(A)所示,LPS 刺激3 h 時,對蝦眼柄中的LvPrx3表達量顯著上升(P<0.05),6 h時恢復到對照組水平(P>0.05),12~48 h 時出現長時間上升(P<0.05)。在刺激12 h 時表達量最大,是對照組的1.79倍。圖5(B)表明,LPS 刺激后凡納濱對蝦腸道中的LvPrx表達量在3~24 h時持續性顯著下調(P<0.05),而在LPS刺激48 h時升至對照組水平(P>0.05)。
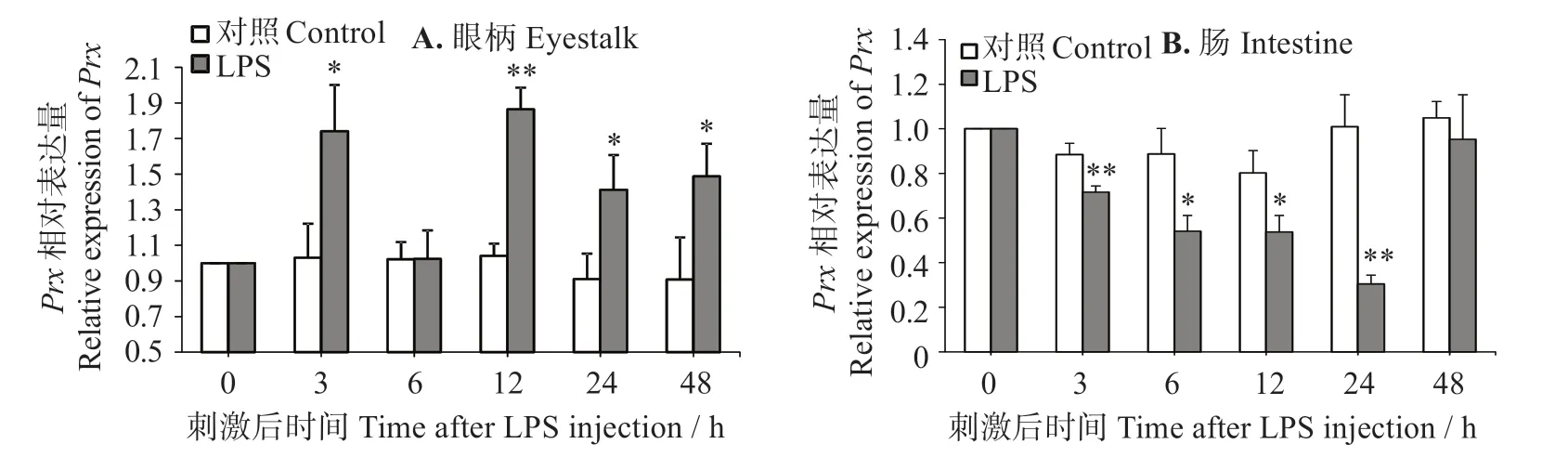
圖5 LPS注射后LvPrx3在蝦眼柄和腸道中的表達量Fig.5 Expression levels of LvPrx3 gene in eyestalk and intestine after LPS injection
圖6(A)可見,凡納濱對蝦眼柄中LvPrx3表達量在4-NP 脅迫后6~24 h 逐漸上調(P<0.01),24 h 時最大,為對照組的4.11倍;脅迫至48 h時LvPrx3表達水平的增加量減少,但仍顯著高于對照組(P<0.05)。圖6(B)表明,凡納濱對蝦腸道中LvPrx3表達量在脅迫6 h 時與對照組無明顯區別(P>0.05),脅迫12 h時顯著上升(P<0.05),24 h 時降低至對照組水平(P>0.05),脅迫48 h時出現明顯下調(P<0.05)。
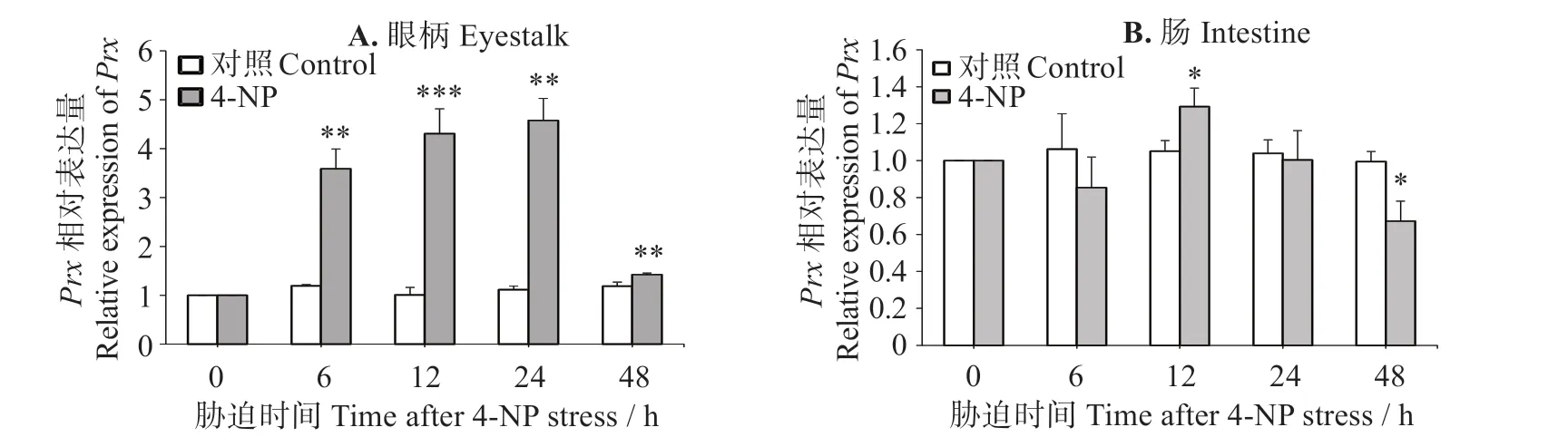
圖6 4-NP脅迫下LvPrx3在眼柄和腸道中的表達量Fig.6 Expression levels of LvPrx3 gene in eyestalk and intestine under 4-NP stress
3 討論
從凡納濱對蝦中克隆到962 bp 的LvPrx3基因cDNA 全長序列,通過蛋白特異性結構域分析和BLAST 比對,確定其屬于典型的2-Cys Prx,有特異的“FYPLDFTFVCPTE”N 端和“GEVCPA”C 端,與多種物種的Prx 氨基酸序列相似性較高。LvPrx3與先前報道的凡納濱對蝦Prx(ACX53642)[21-22]和Prx4(AET36895)[13]的相似性分別為65%、57%。這3 個不同亞型Prx 的序列差異較大,表明它們在對蝦中可能發揮了不同的生理作用。
通過對LvPrx3的蛋白結構進行預測,發現其不僅有典型的2-Cys Prx 的C 端和N 端,還包含催化三聯體結構,二聚體、十聚體界面,過氧化、可溶性半Cys。其中催化三聯體與LvPrx3的蛋白酶促反應相關,而過氧化和可溶性Cys 是使LvPrx3蛋白更易與氧化還原信號分子反應的調節開關。目前特殊的蛋白結構二聚體界面與十聚體界面的功能尚不明確,但蛋白質的聚合狀態對蛋白質的活性有影響,對其表達和激活有決定性作用。Jang等[23-24]研究發現,當生物體內過氧化物濃度處于低水平時,Prx會以單體或二聚體狀態存在;當氧化物濃度處于高水平時,Prx則折疊成穩定的環狀十聚體結構,作為分子伴侶行使抗氧化功能。另外,對LvPrx3進行亞細胞定位分析,發現它主要存在于易聚集活性氧的線粒體中,這與Prx的功能相對應,與中華絨螯蟹EsPrx3[12]的結果一致,表明LvPrx3可能具有抗氧化功能。
基因的組織分布和病理相關表達量變化與其行使的功能密切相關[25]。中國明對蝦(F.chinensis)Prx主要在肝胰腺、卵巢和腸道中有較高的表達量[10],Prx4則在性腺和肝胰腺中高表達[26]。脊尾白蝦(E.carinicauda)Prx5在肝胰腺和卵巢中出現高表達,在眼柄、胃和腸道中低表達[9]。在斑節對蝦(P.monodon)中,Prx1主要在腦、腸道、鰓和肌肉中高表達,而Prx5在肌肉中高表達,在胃、心臟和鰓中的表達量逐漸降低[11]。在日本沼蝦(Macrobrachium nipponense)中,Prx表達量最高的組織是肝胰腺和鰓,最低的是腸道[27]。中華絨螯蟹(E.sinensis)的4種Prx(Prx、Prx3、Prx4和Prx5)均在肝胰腺中有較高的表達水平[12]。在擬穴青蟹(Scylla paramamosain)和遠海梭子蟹(Portunus pelagicus)中,Prx3在肝胰腺中的表達量最高,肌肉(擬穴青蟹)和性腺(遠海梭子蟹)中次之[28-29]。中華新米蝦(Neocaridina denticulata sinensis)Prx3在鰓組織中表達量最高,眼柄中次之,在肝胰腺、性腺和腸道中幾乎無表達[30]。本研究中,LvPrx3在所檢測的6 個組織中均有表達,其中在眼柄中表達量最高,腸道中次之。可見,不同物種及不同亞型Prx的表達具有一定的組織特異性,不同物種各種亞型的Prx主要在特定組織中發揮關鍵作用。本研究的LvPrx3主要在眼柄和腸道的抗氧化過程中發揮作用。
Prx是一類重要的過氧化物酶,主要在清除過氧化物,防止過量活性氧(ROS)引起的氧化損傷中起關鍵作用[7]。甲殼動物Prx在大多數環境脅迫和病原體感染過程中均被誘導并發揮作用[31]。斑節對蝦(P.monodon)在病原菌[哈維弧菌(Vibrio harveyi)、無乳鏈球菌(Streptococcus agalactiae)]感染、鹽度脅迫、pH 脅迫狀態下,鰓中Prx5表達量均一定程度上調,在重金屬銅和鎘脅迫時則被顯著抑制[11]。中國明對蝦(F.chinensis)在受到弧菌(Vibrio anguillarum)刺激時,肝胰腺和血細胞中的Prx轉錄水平均明顯上調[32];在對蝦感染白斑綜合征病毒(WSSV)后,Prx4在這兩種組織中mRNA 表達量顯著升高[26]。在日本囊對蝦(Marsupenaeus japonicus)中,Prx6在感染鰻弧菌(V.anguillarum)后被誘導上調表達,通過調節過氧化物還原酶活性,提高了對蝦的抗細菌能力[33];在感染WSSV后,Prx4被誘導分泌到血淋巴中,促進轉錄因子Dorsal 入核,后誘導抗菌肽基因的表達,抑制WSSV的復制[34]。脊尾白蝦(E.carinicauda)被鰻弧菌(V.anguillarum)和WSSV 脅迫后,肝胰腺和血細胞的Prx5表達量出現先上調后下調趨勢[9]。擬穴青蟹(S.paramamosain)在受到重金屬鎘脅迫時Prx表達量在刺激前期顯著上調[28]。中華絨螯蟹(E.sinensis)在感染嗜水氣單胞菌(Aeromonas hydrophila)的狀態下,Prx、Prx3、Prx4和Prx5表達水平均顯著升高[12]。本研究中,經LPS刺激后,眼柄和腸道的LvPrx3表達量表現出相反的表達模式,眼柄LvPrx3的表達量被顯著誘導上調,腸道LvPrx3表達水平則被顯著抑制,表明LvPrx3在革蘭陰性菌感染時,在眼柄中起重要的抗氧化保護作用,而在腸道中可能不是主要免疫防御因子。4-NP 脅迫結果顯示,眼柄LvPrx3的表達量在所有檢測的時間點被顯著誘導上調,腸道LvPrx3表達量在脅迫中期略有上升,在后期被抑制。這些結果表明LvPrx3在4-NP脅迫時,在眼柄的抗氧化防御中發揮重要作用,也參與了腸道的免疫防御。LPS 刺激和4-NP 脅迫后,均可發現LvPrx3在眼柄中有高度的表達量,表明LvPrx3主要在眼柄中發揮抗氧化保護功能,與LvPrx3在眼柄中基礎表達量最高的結果相吻合。
4 結語
LvPrx3全長cDNA 序列962 bp,ORF 為684 bp,能編碼227個氨基酸。氨基酸序列推導的蛋白分子質量為25.09 ku,pI 為6.22。LvPrx3基因屬于典型的2-Cys Prx,主要定位于線粒體中,包含高度保守的“FYPLDFTDVCPTE”N 端和“GEVCPA”C 端。還具有催化三聯體、二聚體界面、十聚體界面、過氧化和可溶性半胱氨酸等結構域。LvPrx3 與斑節對蝦P.monodon的Prx 氨基酸序列相似性最高。基因LvPrx3在凡納濱對蝦所有檢測的組織中均有表達,在眼柄中表達量最高,腸道次之。LPS 刺激和4-NP脅迫后,LvPrx3在眼柄的表達量均顯著上調,表明LvPrx3主要在眼柄中發揮抗氧化保護功能。
