二氫卟吩鐵對小麥漬水脅迫耐性的影響
2023-05-17 06:52:36楚燕蒙毛穎超蔡劍周琴戴廷波王笑姜東
中國農業科學 2023年10期
關鍵詞:產量
楚燕蒙,毛穎超,蔡劍,周琴,戴廷波,王笑,姜東
二氫卟吩鐵對小麥漬水脅迫耐性的影響
楚燕蒙,毛穎超,蔡劍,周琴,戴廷波,王笑,姜東
南京農業大學農學院/農業農村部作物生理生態與生產管理重點實驗室,南京 210095
【目的】漬水脅迫是影響長江中下游麥區小麥產量提高的主要逆境因子之一。提高小麥耐漬性是實現該區域小麥穩產和增產的重要目標。本研究從植株光合色素含量、光合機構穩定、植株抗氧化能力等角度,明確二氫卟吩鐵提高小麥對開花期漬水脅迫耐性的生理機制,為小麥抗漬栽培提供理論和技術支撐。【方法】以揚麥16為材料,通過設置開花期、灌漿期噴施3個濃度(0.0875、0.126、0.194 mmol·L-1)的二氫卟吩鐵,篩選能夠顯著提高小麥產量的二氫卟吩鐵適宜使用時期和濃度,在此基礎上研究二氫卟吩鐵施用對開花期漬水脅迫下小麥耐性的影響。【結果】與對照相比,不同濃度二氫卟吩鐵在開花期噴施對小麥的增產幅度高于灌漿期噴施處理。研究發現開花期噴施濃度為0.126 mmol·L-1二氫卟吩鐵處理(A2)可顯著提高小麥花后干物質積累量,通過提高千粒重,增加籽粒產量。基于此探究二氫卟吩鐵對小麥耐漬性的影響。開花期漬水脅迫顯著降低了小麥葉片葉綠素含量、凈光合速率和花后干物質積累量與轉運率,導致籽粒產量下降。但在漬水脅迫下與未噴施處理相比,開花期噴施濃度為0.126 mmol·L-1二氫卟吩鐵(AW2)處理表現出較高的光合色素含量、光系統Ⅱ穩定性、凈光合速率,并且提高了抗氧化酶活性,降低了O2-產生速率、H2O2含量,降低了丙二醛含量積累,減輕了漬水脅迫導致的細胞膜脂過氧化傷害,有效緩解了漬水脅迫導致的小麥減產。【結論】開花期噴施0.126 mmol·L-1二氫卟吩鐵可顯著提高小麥產量,并通過減緩開花期漬水脅迫下植株衰老進程,減輕對光合機構損傷、增強抗氧化酶活性,減輕細胞膜脂過氧化傷害,提高小麥葉片光合能力,降低了減產幅度,增強小麥對漬水脅迫的耐性。
小麥;漬水脅迫;二氫卟吩鐵;產量;葉綠素;光合速率
0 引言
【研究意義】漬害是小麥生長發育的重要限制因子,小麥生長發育的不同時期均會受到漬害威脅[1]。長江中下游地區是我國小麥主產區之一,也是冬小麥漬害頻發區[2]。因此,研究如何增強小麥的耐漬性,對實現小麥穩產增產及國家糧食安全具有重要的意義。【前人研究進展】漬水脅迫會導致小麥葉片光合速率降低,植株早衰,產量降低[3-4]。漬水脅迫抑制電子進行光化學反應,過量的電子會與氧氣結合形成大量活性氧[5-6],當抗氧化系統無法清除過量活性氧時,會導致細胞膜脂過氧化傷害[1]。因此,提高小麥對漬水脅迫耐性是生產上亟待解決的重要問題。綜合前人的研究發現,增強小麥耐漬性主要從耐漬品種選育以及配套栽培措施調控兩個方面。耐漬小麥品種選育是解決問題的關鍵[7],但由于植物對漬水脅迫響應的復雜性以及育種的周期較長,目前應用在小麥抗漬生產上的耐漬品種較少。外源調節物質在實現作物抗逆穩產中更能快速發揮緩解作用[8]。外源噴施油菜素內酯會促進黃瓜幼苗根部糖酵解代謝能力,提高抗氧化酶的活性,進而增強黃瓜幼苗對缺氧脅迫的適應性[9]。外源施用生長素和細胞分裂素可以改善漬水條件下植株生長[10],兩種激素協同作用可以提高漬水脅迫下植株葉片的氣孔導度,增強光合能力[11]。二氫卟吩鐵是一種新型的化學調控物質,在油菜越冬期和返青期噴施二氫卟吩鐵均有較明顯的增產效果[12],但在小麥上的應用尚無研究,其對漬水脅迫耐性的影響及其響應機制需要深入探討。【本研究切入點】前期研究發現逆境鍛煉,即在小麥生長前期進行適度漬水鍛煉可顯著增強小麥對關鍵生育期的漬水脅迫耐性,并發現經過鍛煉植株耐漬性增強主要是通過緩解漬水脅迫對小麥葉片光合能力下降以及花后光合產物對籽粒貢獻率的降低,來減緩小麥產量降低幅度,增強耐漬性[13]。但鍛煉模式直接在生產上大規模應用時存在局限性。因此,如果能找到一種外源物質可以實現和漬水鍛煉類似的效果,則可應用在大規模小麥生產上實現抗漬能力增強,穩產增產。二氫卟吩鐵分子結構與葉綠素、血紅蛋白相似,都有卟啉環結構,因此推測二氫卟吩鐵可能通過延緩葉綠素降解,維持光合光合作用及物質轉運來緩解漬水脅迫下小麥產量降低。【擬解決的關鍵問題】以揚麥16為材料,在篩選二氫卟吩鐵適宜施用時期和濃度的基礎上,從葉片葉綠素含量、光合特性、抗氧化能力、物質轉運能力等探究二氫卟吩鐵緩解開花期漬水脅迫,提高小麥耐性的生理機制,研究結果可為小麥抗漬栽培提供理論和技術支撐。
1 材料與方法
1.1 試驗材料與設計
本試驗于2020—2021年在南京農業大學句容基地試驗站進行(32.14°E,119.04°N),農田土壤屬黃棕壤,含有機質12.9 g·kg-1、全氮1.2 g·kg-1、速效氮73.5 mg·kg-1、速效磷44.1 mg·kg-1、速效鉀138.2 mg·kg-1。供試品種為揚麥16,基本苗為16萬/667 m2,11月11日播種,復合肥187.5 kg·hm-2(N﹕P﹕K配比為15﹕6﹕9)為基肥,拔節期追施氮肥120 kg·hm-2。
試驗一:二氫卟吩鐵使用方式的優化方案篩選。選擇3個二氫卟吩鐵噴施濃度(0.0875、0.126和0.194 mmol·L-1),分別在開花期、灌漿期(花后14 d)噴施,試驗處理詳見表1。
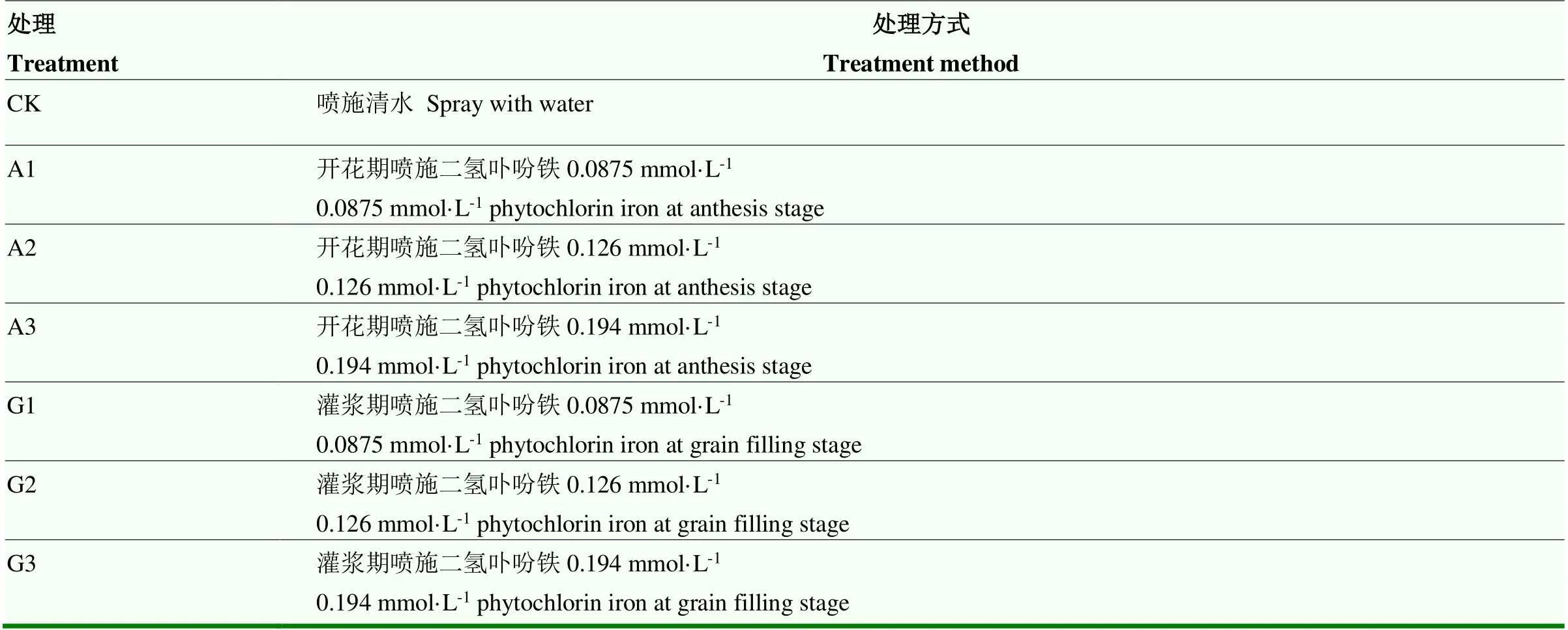
表1 試驗一的處理
試驗二:采用0.126 mmol·L-1濃度的二氫卟吩鐵在開花期噴施,連續噴施2 d。恢復2 d后進行10 d的漬水脅迫,漬水脅迫通過維持2 cm土表水層,試驗處理詳見表2。

表2 試驗二的處理
1.2 測定方法
1.2.1 產量及構成因素 成熟期每小區統計1 m2內小麥的穗數,并進行實收脫粒測產。同時每小區隨機連續取20穗用于測定穗粒數和千粒重。
1.2.2 干物質積累與轉運 開花期選取花期一致的植株進行標花,每個小區分別于開花期和成熟期選取標花的植株20株,分別將各器官于105 ℃殺青30 min,80 ℃烘箱烘干至恒重,稱重并計算各器官的干物質分配比例[14]。
1.2.3 葉綠素含量、光合參數與熒光參數測定 每個小區選取長勢一致的植株同一部位葉片,在漬水脅迫4、10 d時,依次使用葉綠素計(Minolta SPAD-502 Chlorophyll Meter,日本)測定旗葉、倒二葉、倒三葉、倒四葉葉綠素相對含量。
漬水脅迫10 d后,選取長勢一致的植株頂展葉采用便攜式光合速率測定儀LI-6800(Li-Cor公司,美國),于上午9:00—12:00測定旗葉凈光合速率(n)、氣孔導度(s)[15]。
采用FluorPen手持式葉綠素熒光儀(Hansatech公司,中國),用系統設置的固定程序NPQ2測定非光化學淬滅系數(NPQ)、光化學淬滅系數(qP)、PSII最大光化學效率(v/m)、PSII實際光化學效率(ΦPSII)[16]。
1.2.4 活性氧含量及抗氧化酶活性測定 樣品提取參照Tan等[17]的提取方法,在0.5 g鮮樣中加入預冷的PBS(pH 7.8),4 ℃研磨成勻漿,離心20 min(4 ℃,10 000 r/min),收集上清液。
過氧化氫(H2O2)含量使用試劑盒測定。在空白管、標準管、測定管中均加入1 mL試劑一(37 ℃預溫),之后分別加入0.1 mL雙蒸水、0.1 mL標準品應用液、0.1 mL上清液,再共同加入1 mL試劑二。混勻之后用雙蒸水調零,于OD405nm處測定各管吸光度。
超氧陰離子(O2-)產生速率測定參照Tan等[17]的方法,反應體系為0.5 mL上清液,0.5 mL 50 mmol·L-1PBS緩沖液,1 mL 1 mmol·L-1鹽酸羥胺,25 ℃保溫1 h,1 mL 17 mmol·L-1對氨基苯磺酸,1 mL 7 mmol·L-1α-萘胺,25 ℃保溫20 min,測定OD530 nm吸光值。
丙二醛(MDA)含量測定參考趙世杰等[18]的方法,反應體系為4 mL TCA-TBA混合液,2 mL上清液,沸水浴20 min,4 000 r/min離心10 min得上清液,測定OD450 nm、OD532 nm、OD600 nm吸光值。
抗氧化酶活性測定參考Tan等[17]改良的方法,測定超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)和抗壞血酸過氧化物酶(APX)活性。
1.3 數據分析
數據采用SPSS 20.0進行方差分析和顯著性分析,并用Origin 2021進行作圖。
2 結果
2.1 二氫卟吩鐵處理對小麥籽粒產量及其構成因素的影響
產量及其構成要素分析結果表明,與對照(CK)相比,開花期噴施二氫卟吩鐵濃度0.126 mmol·L-1(A2)增產9.8%,其中千粒重與對照相比增加了10.8%(表3)。因此,二氫卟吩鐵處理對小麥產量的提高有促進作用,且開花期噴施二氫卟吩鐵濃度0.126 mmol·L-1處理增產效果最好。
2.2 二氫卟吩鐵處理對小麥干物質積累與轉運的影響
與CK相比,二氫卟吩鐵不同處理均提高了花后光合同化量(PAA)以及光合同化產物對籽粒的貢獻率(CPA),降低了花前貯藏物質轉運量(RAP)及其向籽粒的貢獻率(CTA)。其中,A2處理降低了RAP和CTA,降幅分別為13.1%和18.0%;A2處理提高了PAA和CPA,分別增加了18.2%和11.2%(表4)。
2.3 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥產量及其構成因素的影響
基于試驗一結果,篩選出二氫卟吩鐵對小麥增產效果顯著的處理,即在開花期施用二氫卟吩鐵濃度為0.126 mmol·L-1(A2)。開花期漬水脅迫下,AW2處理的理論產量與實際產量均顯著高于W處理,其中AW2處理千粒重增加8.1%(表5)。
2.4 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥干物質積累與轉運的影響
與CK相比,W、AW2處理的PAA和CPA顯著降低,而提高了RAP、REP、CTA。與W處理相比,二氫卟吩鐵AW2處理降低了RAP及CTA,降幅分別為13.0%和12.1%;且提高了PAA和CPA,增幅分別為9.9%和11.2%(表6)。
同一列不同字母表示品種內處理間在<0.05水平差異顯著。下同
Different small letters in the same column mean significant difference at<0.05 level. the same as below
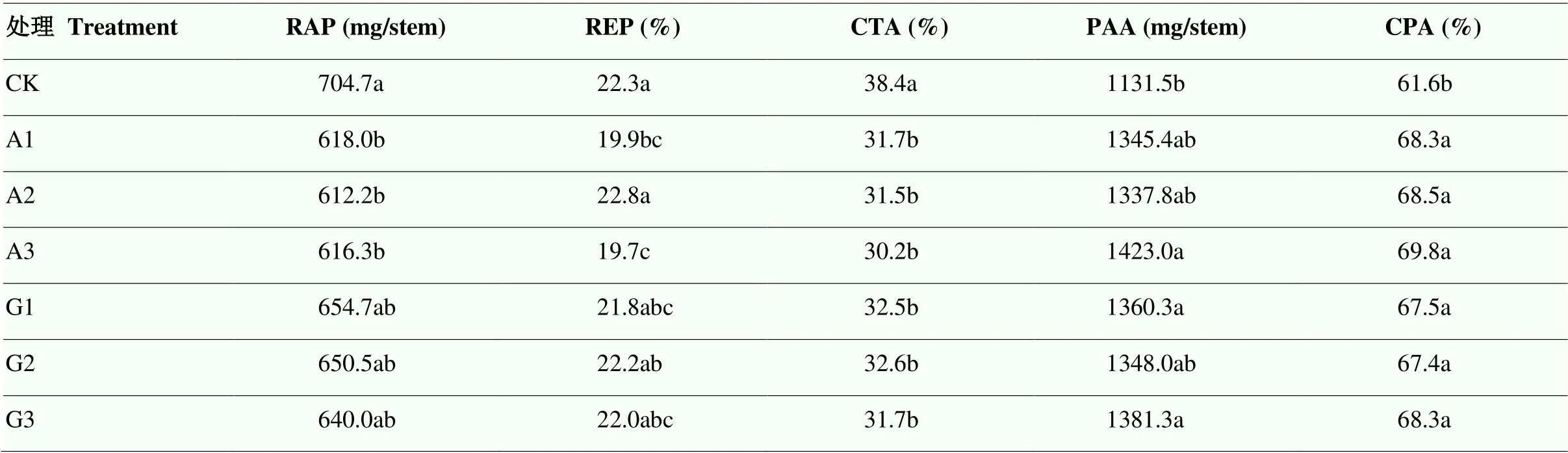
表4 二氫卟吩鐵處理對小麥干物質積累與轉運的影響
RAP:花前貯藏物質運轉量(mg/stem);REP:花前貯藏物質運轉率(%);CTA:花前貯藏物質對籽粒的貢獻率(%);PAA:花后光合同化量(mg/stem);CPA:花后光合同化物對籽粒的貢獻率(%)
RAP: Remobilization amount of pre-anthesis stored dry matter (mg/stem); REP: Remobilization efficiency of pre-anthesis stored dry matter (%); CTA: Contribution rate of pre-anthesis storage substances to grains; PAA: Post-anthesis assimilates accumulation (mg/stem); CPA: Contribution of post-anthesis assimilate to grain (%)

表5 二氫卟吩鐵處理對開花期漬水脅迫下小麥產量及其構成因素的影響
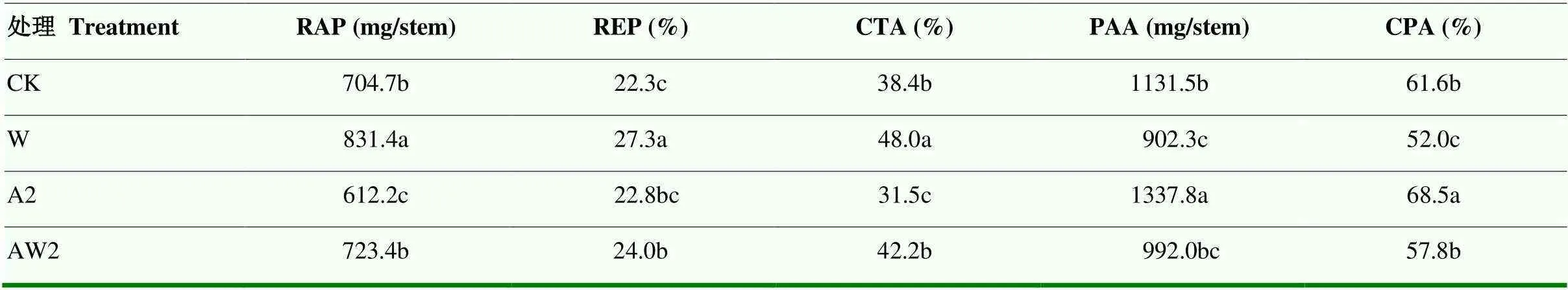
表6 二氫卟吩鐵處理對開花期漬水脅迫下小麥干物質積累和轉運的影響
2.5 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥葉綠素含量的影響
正常生長條件下,二氫卟吩鐵處理提高了下部葉片SPAD,對旗葉SPAD影響不顯著。漬水脅迫第4天,與CK相比,不同二氫卟吩鐵處理均在一定程度上提高了倒2葉、倒3葉、倒4葉的SPAD值,其中A2處理的倒2葉和倒4葉的SPAD值分別顯著提高了1.2%和6.1%。二氫卟吩鐵處理能提高漬水脅迫下的SPAD值,與W處理相比,AW2處理顯著提高了倒2葉、倒3葉和倒4葉的SPAD值(圖1-A)。
漬水脅迫第10天,各處理的小麥旗葉SPAD差異不顯著,但植株下部葉片SPAD值較脅迫第4天均大幅下降,且漬水脅迫下的小麥下部葉片SPAD值下降幅度更為顯著。其中,正常條件下,A2處理的倒4葉SPAD值顯著高于CK;漬水脅迫下,AW2處理的倒4葉、倒3葉和倒2葉SPAD值均顯著高于漬水脅迫W(圖1-B)。
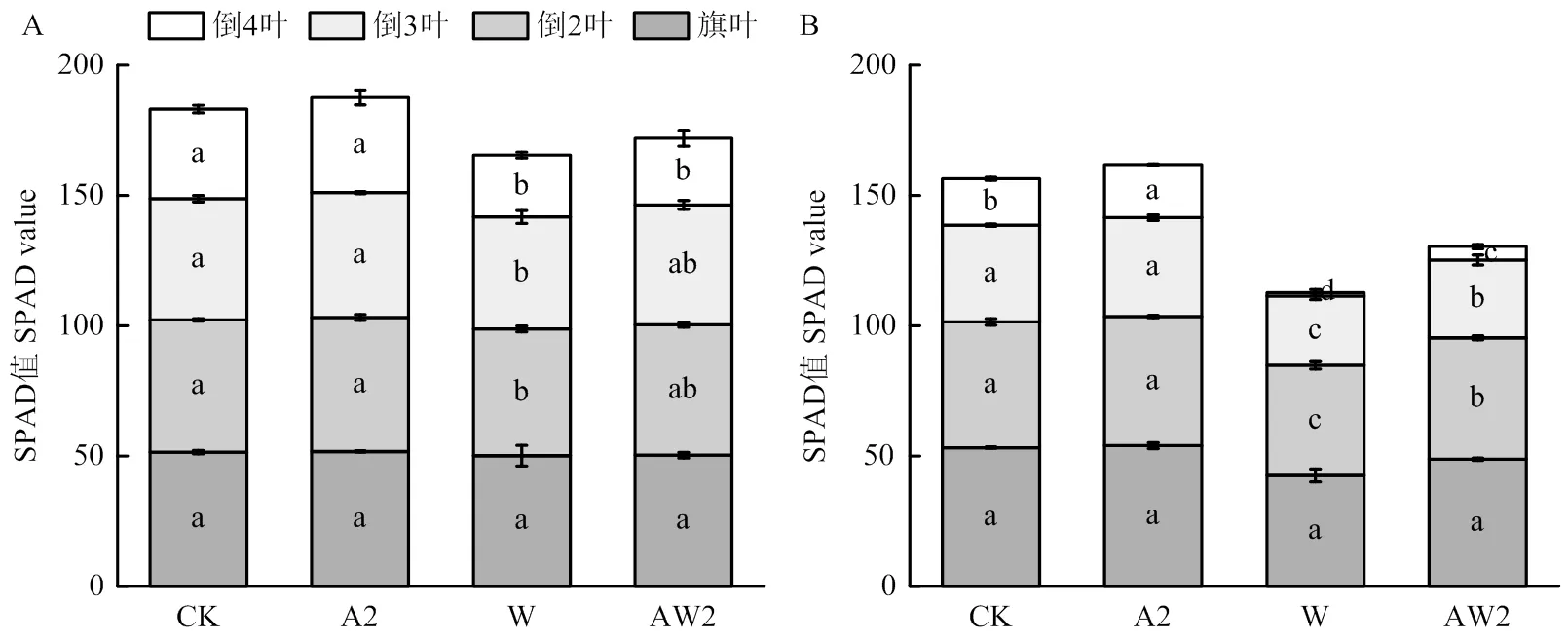
A:漬水脅迫4 d;B:漬水脅迫10 d。圖中不同字母表示處理間在P<0.05水平差異顯著。下同
2.6 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥旗葉光合特性的影響
正常生長條件下,與CK相比,A2處理顯著提高了旗葉凈光合速率(n),增加的幅度為5.6%。A2處理顯著提高了旗葉的氣孔導度(s)23.3%。漬水脅迫下,與W處理相比,AW2處理顯著提高了旗葉的n和s,增幅分別為42.5%和11.3%(圖2)。
2.7 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥旗葉熒光特性的影響
正常生長條件下,與CK相比,A2處理顯著提高了光化學淬滅(qP)。開花期漬水脅迫顯著降低了小麥旗葉v/m、ФPSII和qP,NPQ顯著升高;與W處理相比,AW2處理分別提高了旗葉v/m(3.1%)、ФPSII (14.9%)、qP(26.9%)并顯著降低了NPQ 9.6%(圖3)。
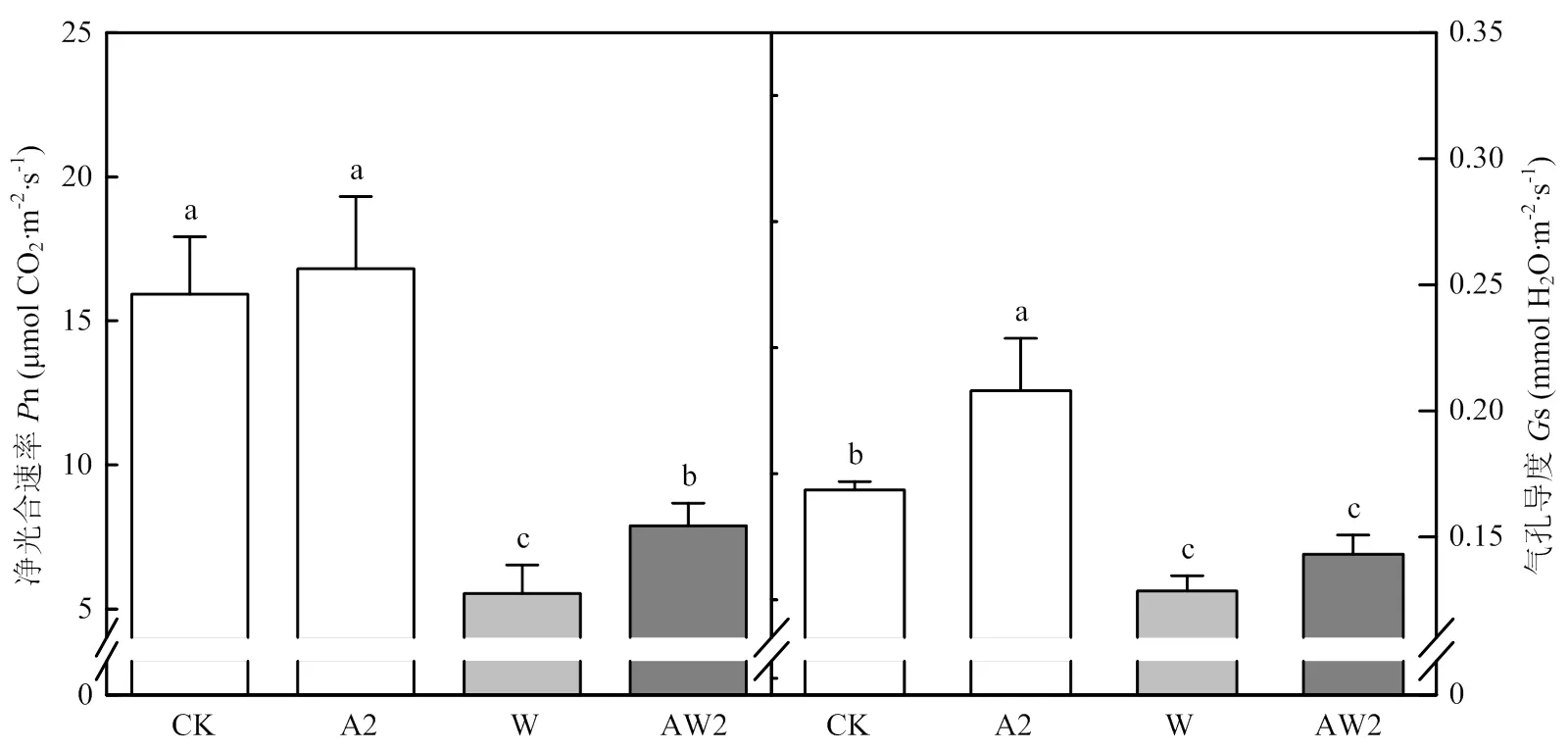
圖2 開花期二氫卟吩鐵處理對其后漬水脅迫后旗葉光合特性的影響
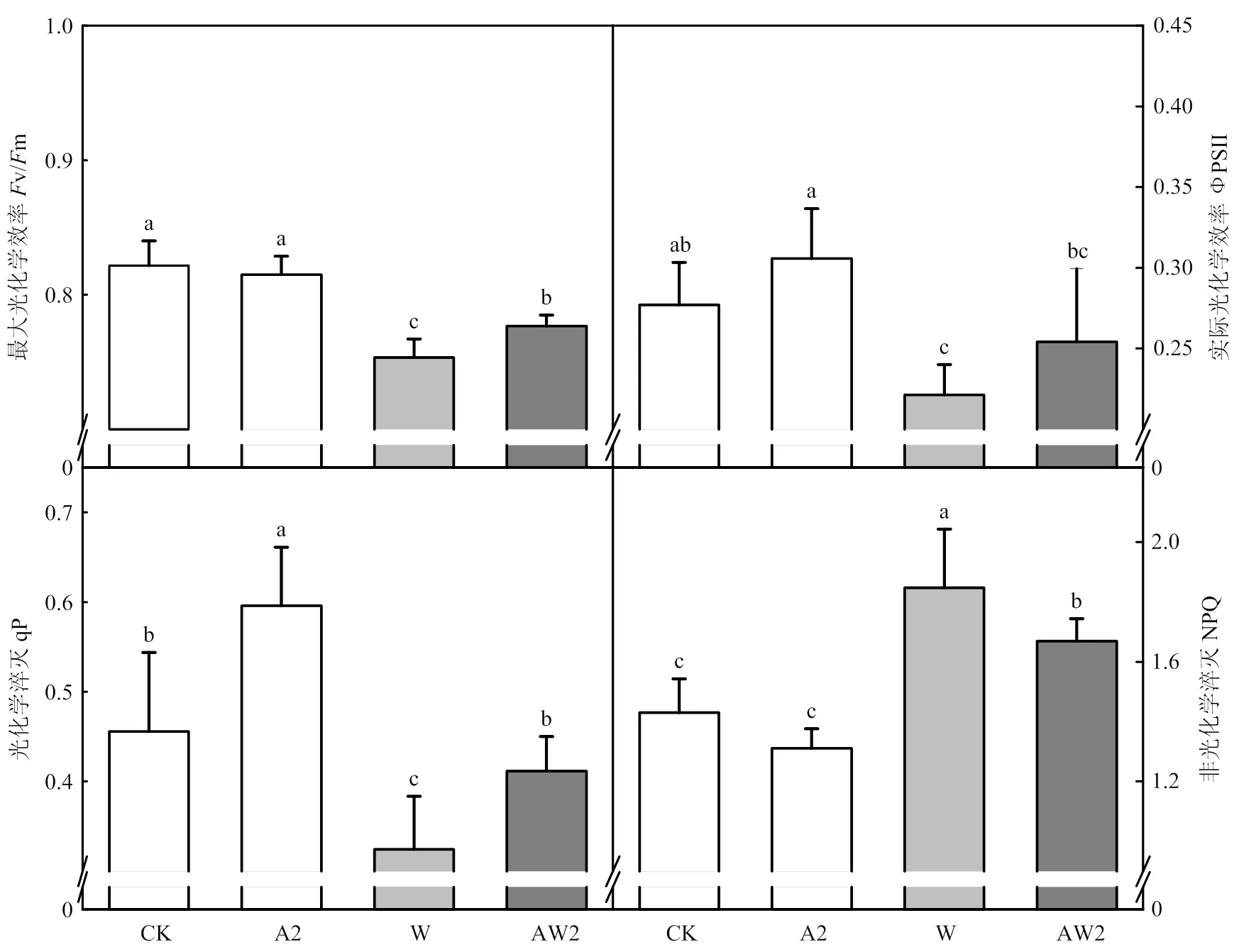
圖3 開花期二氫卟吩鐵處理對其后漬水脅迫后旗葉葉綠素熒光的影響
2.8 開花期二氫卟吩鐵處理對其后漬水脅迫下小麥旗葉活性氧及抗氧化酶活性的影響
正常生長條件下,A2對旗葉O2-產生速率、H2O2含量、MDA含量無顯著影響。漬水脅迫下,與W處理相比,AW2處理的O2-產生速率和H2O2含量分別降低24.4%和24.4%。此外,AW2處理的MDA含量顯著低于W處理,降低了21.9%(圖4)。
正常生長條件下,與CK相比,A2處理對旗葉SOD、POD、APX、CAT無顯著影響。漬水脅迫顯著降低了SOD、APX活性,顯著提高了POD、CAT的活性。與W處理相比,AW2處理使得SOD活性提高24.7%,POD活性提高19.6%,APX活性提高12.6%,CAT活性提高11.0%(圖5)。
2.9 各生理指標與產量相關性分析
產量與千粒重、n、v/m、SPAD、SOD活性呈顯著正相關,與MDA的含量呈顯著負相關(圖6)。SOD酶活性與MDA含量呈現顯著負相關,說明SOD在漬水脅迫下活性氧清除過程中扮演重要作用,從而緩解了細胞膜脂過氧化傷害。
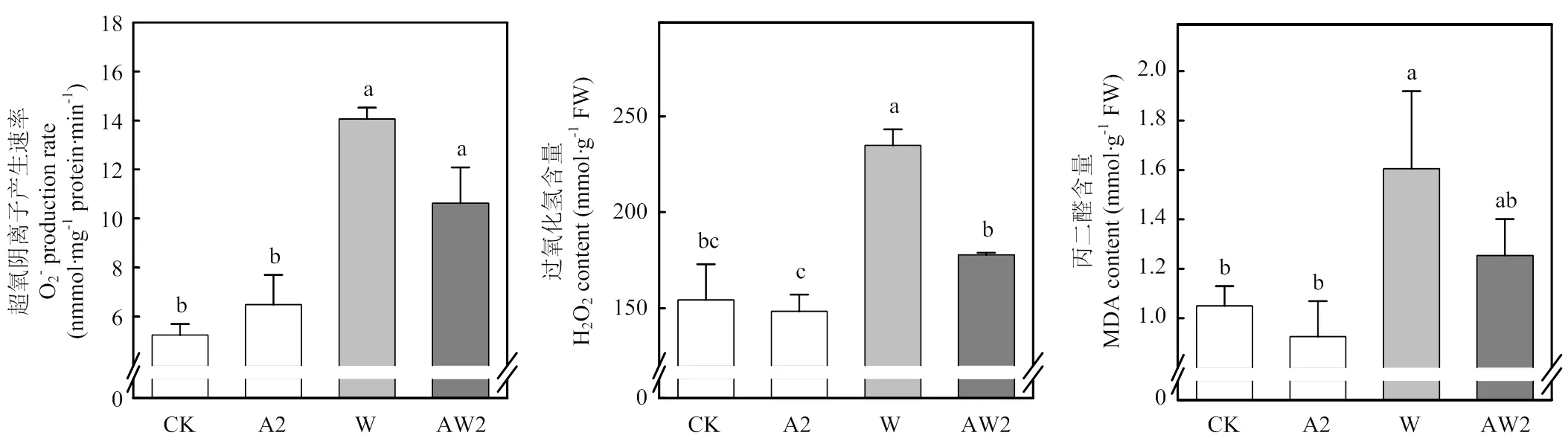
圖4 開花期二氫卟吩鐵處理對其后漬水脅迫后小麥旗葉活性氧的影響
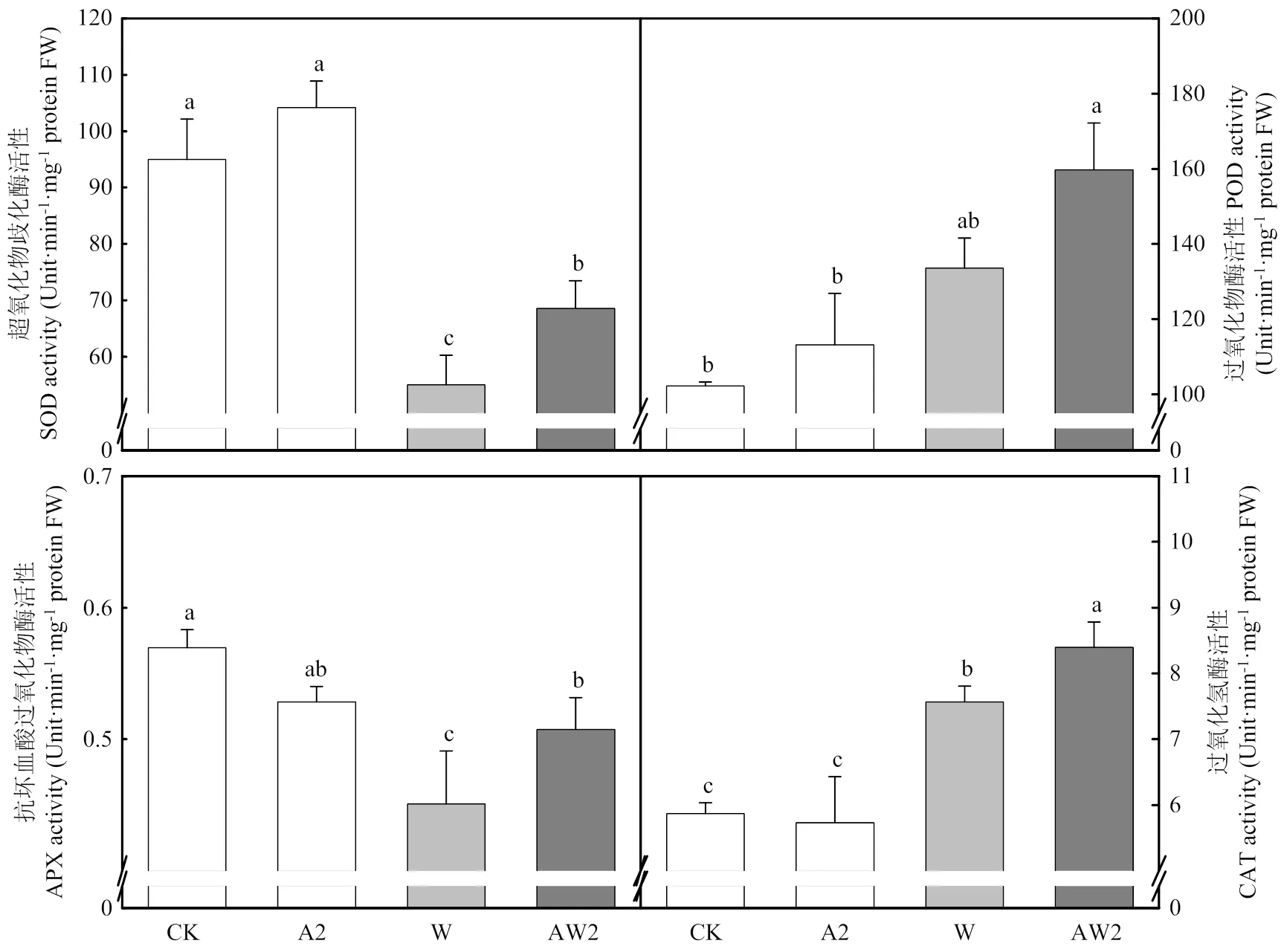
圖5 開花期二氫卟吩鐵處理對其后漬水脅迫后小麥旗葉抗氧化酶的影響
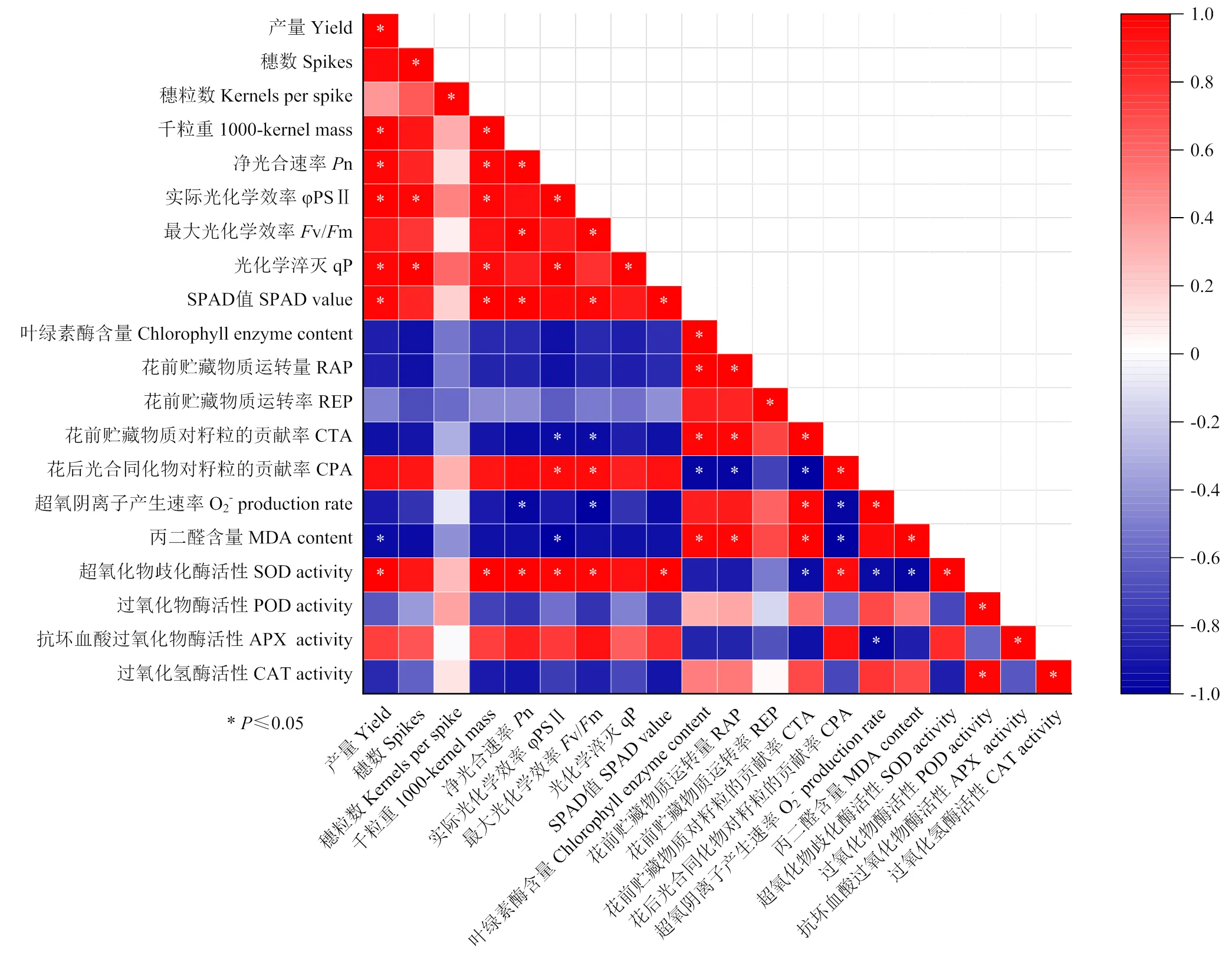
*表示處理間在P<0.05水平差異顯著 * means significant difference at P<0.05 level among the treatments
3 討論
3.1 二氫卟吩鐵可顯著提高小麥對開花期漬水脅迫的耐受性
長江中下游地區是我國小麥主產區之一,小麥生長中后期的大范圍降雨是該區域小麥產量提高的主要限制因子[19]。開花期漬水脅迫顯著降低了小麥的穗粒數和千粒重,縮短籽粒灌漿時間,降低花后光合同化物的積累及轉運,導致產量降低[20]。因此,增強小麥耐漬性是該區域小麥穩定生產迫切需要解決的問題。本研究結果表明,在開花期噴施0.126 mmol·L-1的二氫卟吩鐵可顯著提高花后光合同化產物對籽粒貢獻率,提高小麥產量。漬水脅迫前噴施二氫卟吩鐵可顯著提高千粒重,從而緩解漬水脅迫下產量損失。
前人研究發現,拔節期和花后漬水處理條件下,隨漬水持續時間延長,花后同化物積累對小麥產量的貢獻率逐漸降低[21]。本研究表明,開花期漬水脅迫顯著降低了花后光合同化物向籽粒的轉運,提高了花前貯藏物質的轉運,但二氫卟吩鐵處理提高了花后光合同化量及光合同化產物對籽粒的貢獻率,降低了漬水脅迫導致的產量損失。本研究中,開花期漬水脅迫并沒有顯著影響穗粒數,原因一方面可能與二氫卟吩鐵處理的時間有關,二氫卟吩鐵在花期噴施2 d,然后恢復2 d后,進行漬水脅迫,田間漬水脅迫在早期并沒有影響到穗粒數形成,而主要是隨著漬水時間延長后導致了籽粒灌漿不足,千粒重降低;另一方面可能與品種對漬水脅迫的敏感性差異有關,李霞等[22]研究發現不同品種在開花期漬水脅迫下,有些品種是影響穗粒數形成,有些品種是影響千粒重,導致減產。
3.2 二氫卟吩鐵緩解了其后漬水脅迫下小麥葉片光合能力的降低
小麥葉片在漬水脅迫下易出現黃化,影響植株光合作用[23-24],本研究發現隨著漬水脅迫處理時間延長,植株葉綠素含量顯著降低,在漬水10 d后,小麥除旗葉外其他部位葉片葉綠素含量降低較快,且與直接漬水處理相比,經過二氫卟吩鐵處理的植株主要是通過緩解了倒2葉至倒4葉的葉綠素降低,延緩了植株衰老,維持較高的葉片光合能力。葉綠素熒光參數可表征作物對環境脅迫的響應[25]。花后漬水脅迫使小麥旗葉光反應中心PSⅡ的開放程度下降,實際光化學效率(ΦPSⅡ)降低,且耐漬品種的v/m值顯著高于不耐漬品種[26-27]。本研究表明,漬水脅迫處理下,二氫卟吩鐵處理顯著提高了小麥旗葉v/m、ΦPSII,減小了NPQ,說明可緩解漬水脅迫對光系統II的破壞效應,這與二氫卟吩鐵處理植株保持較高的光合速率結果一致。綜上,二氫卟吩鐵處理后植株在漬水脅迫下通過維持較高的光合機構穩定,從而維持較高的光合能力。
3.3 二氫卟吩鐵減輕了其后漬水脅迫下小麥細胞膜脂過氧化傷害
漬水脅迫誘使植物體內活性氧類物質(O2-、H2O2等)大量積累[28-29],破壞活性氧清除機制,加劇膜脂過氧化過程[30],對植物細胞膜、DNA和蛋白質造成嚴重損傷[1]。過量的ROS會導致葉綠體質膜降解,PSII反應中心受損,破壞光合系統[31],從而限制作物生長發育[32-33]。漬水脅迫顯著提高了小麥葉片O2-產生速率、H2O2含量,MDA積累,加劇膜脂過氧化傷害,而漬水脅迫前噴施二氫卟吩鐵處理植株旗葉中的MDA、H2O2含量和O2-產生速率明顯低于未噴施處理。
作物可以通過啟動抗氧化系統來維持活性氧的動態平衡,從而降低氧化損傷的程度[34-35],SOD是抵抗活性氧損傷的第一道防線,通過催化O2-生成H2O2,減少O2-含量,生成的H2O2通過CAT、APX等酶進一步轉化為H2O[36]。APX在ASA-GSH循環中發揮了關鍵作用,能最大限度地減少H2O2含量和維持氧化還原穩態[37]。在本研究中,漬水脅迫下小麥葉片CAT和POD活性顯著增強以應對O2-和H2O2含量的過量增加,漬水脅迫前噴施二氫卟吩鐵處理顯著提高了SOD、CAT、POD和APX的活性,其中SOD酶活性與MDA含量呈現顯著負相關。說明二氫卟吩鐵通過上調抗氧化酶活性,提高了葉片細胞對O2-和H2O2的清除效率,減輕膜脂過氧化物積累,最終緩解了其后漬水脅迫對小麥細胞膜脂過氧化傷害。
4 結論
開花期噴施二氫卟吩鐵顯著提高了小麥對其后發生漬水脅迫的耐性。耐性增強的主要生理基礎是通過顯著減緩植株衰老,減輕對光合機構損傷,同時提高抗氧化酶活性,清除產生的過量活性氧,進而減輕了漬水脅迫對細胞膜脂過氧化傷害,從而維持較高的光化學效率,促進了花后光合同化物向籽粒的轉運,最終降低了漬水脅迫下小麥減產幅度,提高植株的耐漬性。研究結果為小麥抗漬栽培技術提供了重要的理論和技術支撐。
[1] ANNALISA P, SOFIA C, ANTONELLA L, ANNA B, ROSSELLA N, LAURA D G. ROS production and scavenging under anoxia and re- oxygenation incells: a balance between redox signaling and impairment. Frontiers in Plant Science, 2016, 7(1): 1803-1814.
[2] MIN Y, ZHUQING Z, XIANGYI D, JIWEI L, FANGZHU M, YUANHONG Q. Physiological mechanism of programmed cell death aggravation and acceleration in wheat endosperm cells caused by waterlogging. Acta Physiologiae Plantarum, 2017, 39(1): 23-34.
[3] 蔡永萍, 陶漢之, 張玉瓊. 土壤漬水對小麥開花后葉片幾種生理特性的影響. 植物生理學通訊, 2000, 36(2): 110-113.
CAI Y P, TAO H Z, ZHANG Y Q. The effect of soil waterlogging on several physiological characteristics of wheat leaves after flowering. Plant Physiology Communications, 2000, 36(2): 110-113. (in Chinese)
[4] 范雪梅, 姜東, 戴廷波, 荊奇, 曹衛星. 花后干旱和漬水下氮素供應對小麥旗葉衰老和粒重的影響. 土壤學報, 2005, 42(5): 173-177.
FAN X M, JIANG D, DAI T B, JING Q, CAO W X. Effects of nitrogen supply on flag leaf senescence and grain weight in wheat grown under drought or waterlogging from anthesis to maturity. Acta Pedologica Sinica, 2005, 42(5): 173-177. (in Chinese)
[5] YAN K, ZHAO S, CUI M, HAN G, WEN P. Vulnerability of photosynthesis and photosystem I in Jerusalem artichoke (L.) exposed to waterlogging. Plant Physiology & Biochemistry, 2018, 125(1): 239-246.
[6] 吳曉麗, 湯永祿, 李朝蘇, 吳春, 黃鋼. 不同生育時期漬水對冬小麥旗葉葉綠素熒光及籽粒灌漿特性的影響. 中國生態農業學報, 2015, 23(3): 309-318.
WU X L, TANG Y L, LI C S, WU C, HUANG G. Effect of waterlogging at different growth stages on flag leaf chlorophyll fluorescence and grain-filling properties of winter wheat. Chinese Journal of Eco-Agriculture, 2015, 23(3): 309-318. (in Chinese)
[7] 曹陽, 蔡士賓, 方先文, 朱偉. 利用馬卡小麥的強耐濕性創造新的耐濕種質. 江蘇農業科學, 1998(1): 20-21.
CAO Y, CAI S B, FANG X W, ZHU W. Using the strong moisture tolerance of Tr macha wheat to create new moisture tolerant germplasm. Jiangsu Agricultural Sciences, 1998(1): 20-21. (in Chinese)
[8] 王飛, 彭少兵. 水稻綠色高產栽培技術研究進展. 生命科學, 2018, 30(10): 1129-1136.
WANG F, PENG S B. Research progress on green and high-yield cultivation techniques of rice. Chinese Bulletin of Life Sciences, 2018, 30(10): 1129-1136. (in Chinese)
[9] NAZIR F, FARIDUDDIN Q, HUSSAIN A. Brassinosteroid and hydrogen peroxide improve photosynthetic machinery, stomatal movement, root morphology and cell viability and reduce Cu- triggered oxidative burst in tomato. Ecotoxicology and Environmental Safety, 2020, 207(1): 81-111.
[10] PANG J, CUINuin T, SHABALA L. Effect of secondary metabolites associated with anaerobic soil conditions on ion fluxes and electrophysiology in barley roots. Plant Physiology, 2007, 145(1): 266-276.
[11] DREW C M, LYNCH M J. Soil anaerobiosis, microorganisms, and root function. Annual Review of Phytopathology, 1980, 18(1): 37-66.
[12] 郭麗華, 唐為愛, 李萬梅. 0.02%二氫卟吩鐵DP調節油菜生長的藥效試驗. 上海蔬菜, 2016(2): 53-54.
GUO L H, TANG W A, LI W M. Efficacy test of 0.02% phytochlorin iron DP regulating rapeseed growth. Shanghai Vegetables, 2016(2): 53-54. (in Chinese)
[13] 馬博聞, 李慶, 蔡劍, 周琴, 黃梅, 戴廷波, 王笑, 姜東. 花前漬水鍛煉調控花后小麥耐漬性的生理機制研究. 作物學報, 2022, 48(1): 151-164.
MA B W, LI Q, CAI J, ZHOU Q, HUANG M, DAI T G, WANG X, JIANG D. Study on the physiological mechanism of waterlogging exercise before anthesis regulating waterlogging tolerance of post anthesis wheat. Acta Agronomica Sinica, 2022, 48(1): 151-164. (in Chinese)
[14] 丁錦峰, 蘇盛楠, 梁鵬, 江孟孟, 鄭麗潔, 汪先鵬, 李春燕, 朱新開, 郭文善. 拔節期和花后漬水對小麥產量、干物質及氮素積累和轉運的影響. 麥類作物學報, 2017, 37(11): 1473-1479.
DING J F, SU S N, LIANG P, JIANG M M, ZHENG L J, WANG X P, LI C Y, ZHU X K, GUO W S. Effect of waterlogging at elongation or after anthesis on grain yield and accumulation and remobilization of dry matter and nitrogen in wheat. Journal of Triticeae Crops, 2017, 37(11): 1473-1479. (in Chinese)
[15] GAO J, WANG F, SUN J, TIAN Z, HU H, JIANG S, LUO Q, XU Y, JIANG D, CAO W, DAI T. Enhanced Rubisco activation associated with maintenance of electron transport alleviates inhibition of photosynthesis under low nitrogen conditions in winter wheat seedlings. Journal of Experimental Botany, 2018, 69(22): 5477-5488.
[16] YAKUN C, ZHONGWEI T, XU Z, ABID M, HUIMIN H, DONG J, WEIXING C, TINGBO D. Effect of water deficit during vegetative growth periods on post-anthesis photosynthetic capacity and grain yield in winter wheat (L.). Acta Physiologiae Plantarum, 2015, 37(10): 1-10.
[17] TAN W, LIU J, DAI T, JING Q, CAO W, JIANG D. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis water-logging. Photosynthetica, 2008, 46(1): 21-27.
[18] 趙世杰, 許長成, 鄒琦, 孟慶偉. 植物組織中丙二醛測定方法的改進. 植物生理學通訊, 1994, 30(3): 207-210.
ZHAO S J, XU Z C, ZOU Q, MENG Q W. Improvement of method for measurement malondialdehyde in plant tissue. Plant Physiology Communications, 1994, 30(3): 207-210. (in Chinese)
[19] SHAO G C, LAN J J, YU S E, LIU N, GUO R Q, SHE D L. Photosynthesis and growth of winter wheat in response to waterlogging at different growth stages. Photosynthetica, 2013, 51(3): 429-437.
[20] 李赟, 李霞, 李慕嶸, 楊蕊, 王小燕. 開花期漬水對小麥產量及氮素分配的影響. 麥類作物學報, 2022, 42(6): 1-9.
LI Y, LI X, LI M R, YANG R, WANG X Y. Effects of waterlogging during flowering on wheat yield and nitrogen distribution. Journal of Triticeae crops, 2022, 42(6): 1-9. (in Chinese)
[21] XIAO W, MEI H, QIN Z, JIAN C, TINGBO D, WEIXING C, DONG J. Physiological and proteomic mechanisms of waterlogging priming improves tolerance to waterlogging stress in wheat (L.). Environmental and Experimental Botany, 2016, 132: 175-182.
[22] 李霞, 李赟, 李慕嶸, 尹立俊, 王小燕, 漆棟良. 開花期漬水對土壤不同形態氮素含量及小麥氮素積累運轉和產量的影響. 南方農業學報, 2022, 53(7): 1883-1892.
LI X, LI Y, LI M R, YI L J, WANG X Y, QI D L. Effects of waterlogging at anthesis stage on nitrogen content of different forms in soil and nitrogen accumulation and translocation of wheat and yield. Journal of Southern Agriculture, 2022, 53(7): 1883-1892. (in Chinese)
[23] ARAKI H, HAMADA A, HOSSAIN M A, TAKAHASHI T. Waterlogging at jointing and/or after anthesis in wheat induces early leaf senescence and impairs grain filling. Field Crops Research, 2012, 137(9): 27-36.
[24] ROSSI S, BURGESS P, JESPERSEN D, HUANG B R. Heat-induced leaf senescence associated with chlorophyll metabolism in bentgrass lines differing in heat tolerance. Crop Science, 2017, 57: 169-178.
[25] LIU L Z, WU J J, ZHOU H K, LI X H, WANG Q F, AN X L, LI R T. Chlorophyll fluorescence and its progress in detecting water stress. Spectroscopy and Spectral Analysis, 2017, 37(9): 2780-2787.
[26] 譚維娜, 戴廷波, 荊奇, 曹衛星, 姜東. 花后漬水對小麥旗葉光合特性及產量的影響. 麥類作物學報, 2007, 27(2): 314-317.
TAN W N, DAI T B, JING Q, CAO W X, JIANG D. Effects of post anthesis waterlogging on photosynthetic characteristics and yield of flag leaves of wheat. Journal of Triticeae Crops, 2007, 27(2): 314-317. (in Chinese)
[27] WU X L, TANG Y L, LI C S, MCHUGH A D, LI Z, WU C. Individual and combined effects of soil waterlogging and compaction on physiological characteristic of wheat in southwestern China. Field Crops Research, 2018, 215: 163-172.
[28] ASADA K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant physiology, 2006, 141(2): 391-396.
[29] RAPPAPORT F, GUERGOVA-KURAS M, NIXION P J. Kinetics and pathways of charge recombination in photosystem II. Biochemistry, 2002, 41(26): 8518-8527.
[30] SLESAK I, LIBIK M, KARPINSKA B. The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses. Acta biochimica Polonica, 2007, 54(1): 39-50.
[31] YAMAUCHI T, WATANABE K, FUKAZAWA A. Ethylene and reactive oxygen species are involved in root aerenchyma formation and adaptation of wheat seedlings to oxygen-deficient conditions. Journal of experimental botany, 2014, 65(1): 261-273.
[32] DE SAN C R P, ABELEDO L G, MIRALLES D J. Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant & Soil, 2014, 378(12): 265-277.
[33] ZHANG R, YUE Z, CHEN X. Foliar applications of urea and melatonin to alleviate waterlogging stress on photosynthesis and antioxidant metabolism in sorghum seedlings. Plant Growth Regulation, 2022, 97: 429-438.
[34] GUOPING Z, KOJI T, JUN A, SHIGENORI M. Influence of waterlogging on some anti-oxidative enzymatic activities of two barley genotypes differing in anoxia tolerance. Acta Physiologiae Plantarum, 2007, 29(2): 171-176.
[35] DOUPIS G, KAVROULAKIS N, PSARRAS G, PAPADAKIS I. Growth, photosynthetic performance and antioxidative response of ‘Hass’ and ‘Fuerte’ avocado (Mill.) plants grown under high soil moisture. Photosynthetica: International Journal for Photosynthesis Research, 2017, 55(4): 655-663.
[36] RUCHI B, JAI P S. Antioxidative defense system in pigeonpea roots under waterlogging stress. Acta Physiologiae Plantarum, 2012, 34(4): 515-522.
[37] MIRZA H, BHUYAN M H, TAUFIKA I A, KHURSHEDA P, KAMRUN N, JUBAYER A M, MASAYUKI F. Regulation of ascorbate- glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants, 2019, 8(9): 384-396.
Effect of Phytochlorin Iron on Stress Tolerance to Waterlogging in Wheat
CHU YanMeng, MAO YingChao, CAI Jian, ZHOU Qin, DAI TingBo, WANG Xiao, JIANG Dong
College of Agriculture, Nanjing Agricultural University/Key Laboratory of Crop Physiology, Ecology and Production Management, Ministry of Agriculture and Rural Affairs, Nanjing 210095
【Objective】Waterlogging stress is one of the main limiting factors for wheat production, especially in the middle and lower reaches of the Yangtze River in China. Improving the waterlogging tolerance of wheat is an important goal to achieve stable and increased yield in this region. In this study, by exploring the suitable use period and concentration of phytochlorin iron, its role in improving waterlogging stress tolerance was further evaluated mainly from the perspectives of plant photosynthesis and plant antioxidant capacity. The research results could provide the theoretical and technical support for waterlogging-resistant cultivation of wheat. 【Method】Using Yangmai 16 as material, three concentrations (0.0875, 0.126, and 0.194 mmol·L-1) of phytochlorin iron were set at anthesis and grain filling stages to screen the appropriate period and concentration for achieving a significant increase in wheat yield. Based on this, the effect of phytochlorin iron on wheat tolerance to waterlogging stress at anthesis stage was further evaluated.【Result】Compared with control, treatment with a concentration of 0.126 mmol·L-1phytochlorin iron at anthesis stage (A2) could significantly increase wheat grain yield by increasing the grain weight. Waterlogging stress at anthesis stage significantly reduced the chlorophyll content, net photosynthesis rate, and post-flowering dry matter accumulation and translocation to grain, resulting in grain yield reduction. However, compared with non-spraying treatment, AW2 treatment showed a higher photosynthetic pigment content, photosystem II stability, net photosynthetic rate. Meantime, the raised activities of antioxidant enzymes, reduced O2-production rate and H2O2content, which showed correspondence with the reduced accumulation of malondialdehyde content, thus alleviated the damage of cell membrane lipid peroxidation and the yield reduction caused by waterlogging stress.【Conclusion】Spraying a concentration of 0.126 mmol·L-1phytochlorin iron at anthesis stage could significantly increase wheat yield. Phytochlorin iron could alleviate the plant senescence, reduce damage to PSII, enhance the activity of antioxidant enzymes, reduce the damage of cell membrane lipid peroxidation, maintain higher photosynthetic rate, reduce the degree of yield reduction, and enhance wheat tolerance to waterlogging stress.
wheat; waterlogging stress; phytochlorin iron; yield; chlorophyll; photosynthesis rate

10.3864/j.issn.0578-1752.2023.10.003
2022-08-24;
2023-01-09
國家自然科學基金(32272213,31771693,U1803235)、國家現代小麥產業技術體系(CARS-03)、江蘇省協同創新中心(JCIC-MCP)
楚燕蒙,E-mail:2020801194@stu.njau.edu.cn。通信作者王笑,E-mail:xiaowang@njau.edu.cn
(責任編輯 楊鑫浩,岳梅)
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
