鹽磷脅迫對木麻黃和臺灣相思種子萌發及幼苗生長的影響
2023-05-22 06:19:52袁鋒羅倩夏心慧潘國營李茂瑾江傳陽蘇親桂陳燦
廣西植物 2023年4期
袁鋒 羅倩 夏心慧 潘國營 李茂瑾 江傳陽 蘇親桂 陳燦
摘 要:惡劣環境下,人工海防林因面臨養分脅迫而經營困難。為探討鹽、磷脅迫對主要海防林樹種木麻黃和臺灣相思種子萌發及生長的影響,該研究分別用不同濃度的NaCl(鹽)和KH2PO4(磷)溶液處理種子和澆灌幼苗,測定種子萌發和幼苗生長指標。結果表明:(1)高鹽脅迫顯著抑制種子萌發,對幼苗生長有一定影響,但兩種植物影響程度不同;臺灣相思種子萌發耐鹽性高于木麻黃,前者相對鹽害率最大值為23.03%,后者為89.15%;隨著鹽濃度增加,木麻黃和臺灣相思種子的發芽率、發芽勢、發芽指數和活力指數均降低,對應最大值分別為38.70%、34.67%、18.70、0.055和76.67%、62.22%、48.46、6.11。(2)兩種植物的株高和根長隨鹽濃度增加而降低,木麻黃和臺灣相思株高分別為12.29~6.01 mm和48.27~17.33 mm,根長分別為8.57~1.45 mm和33.41~5.88 mm;臺灣相思根、莖、葉生物量及根冠比均隨鹽濃度的增加逐漸減小,木麻黃各處理差異較小。 (3) 臺灣相思的種子和幼苗較木麻黃更耐低磷環境,二者最適磷濃度存在差異;木麻黃種子發芽率(49.33%)、發芽指數(23.12 %)、活力指數(0.093)及根莖葉生物量最大值均在0.1 mmol·L-1磷處理;臺灣相思種子發芽率(81.11%)、發芽勢(62.22%)、發芽指數(38.23)、活力指數(5.07)、株高(54.48 mm)及根長(37.16 mm)最大值均在5 mmol·L-1磷處理,10 mmol·L-1磷處理抑制其萌發。因此,播種海防林種子或種植幼苗時須根據土壤鹽和磷的含量選擇播種或種植方式。
關鍵詞: 養分脅迫, 木麻黃, 臺灣相思, 種子萌發, 幼苗生長
中圖分類號:Q945
文獻標識碼:A
文章編號:1000-3142(2023)04-0587-09
Abstract:Artificial coastal protection forests are difficult to manage because ofnutrient stress in harsh environment. To explore the effects of salt and phosphorus stress on the seed germination and seedling growth of Casuarina equisetifolia and Acacia confusa, we respectively watered the seeds and seedlings of the two kinds of plants with the solutions of NaCl (salt) and KH2PO4 (phosphorus) in different concentrations, and then determined their growth indicators, respectively. The results were as follows: (1) High salt concentration solutions significantly inhibited seed germination and had a certain effect on the growth of seedlings, however, the effects of salt stress on seed gemination and growth were different. The salt tolerance of Acacia confusa seed germination was higher than that of Casuarina equisetifolia. The biggest relative salt damage rate of Acacia confusa was 23.03% and that of Casuarina equisetifoliawas 89.15%. As the concentration of NaCl increased, the germination rate, germination potential, germination index and vitality index of Casuarina equisetifolia and Acacia confusa seeds decreased. And the maximums of the four germination indexes of Casuarina equisetifolia seed were 38.70%, 34.67%, 18.70 and 0.055, and of Acacia confusa seeds were 76.67%, 62.22%, 48.46 and 6.11, respectively. (2) The height and root length of the plants decreased with the increase of salt concentration. The plant heights of Casuarina equisetifolia and Acacia confusa were 12.29 to 6.01 mm and 48.27 to 17.33 mm, and the root lengths were 8.57 to 1.45 mm and 33.41 to 5.88 mm, respectively. The biomass of root, stem and leaf as well as rootshoot ratio of Acacia confusa gradually decreased with the increase of salt concentration, but the differences between the treatments of Casuarina equisetifolia were not obvious. 0.6% and 0.4% salt stress were the thresholds for seed germination and seedling growth of the two plants, respectively. (3)The seed and seedling of Acacia confusa were more tolerant to low-phosphorus environment than Casuarina equisetifolia, and there was a difference in the optimum phosphorus concentration between the two. When the phosphorus concentration was 0.1 mmol·L-1, the seed germination rate(49.33%), germination index (23.12%) and vitality index(0.093) of Casuarina equisetifolia, as well as the biomass of root, stem and leaf, all reached the maximum. In the 5 mmol·L-1 treatment, the maximums of germination rate(81.11%), germination potential(62.22%), gemination index(38.23), vitality index(5.07), plant height (54.48 mm) and root length (37.16 mm) of Acacia confusa seeds all appeared, while their germination was inhibited in the treatment of 10 mmol·L-1. Therefore, when sowing the seed of coastal defense forest or planting its seedling, the sowing or planting mode must be selected according to the soil salt and phosphorus contents.
Key words: nutrient stress, Casuarina equisetifolia, Acacia confusa, seed germination, seedling growth
種子順利萌發、成苗是植物能否完成生活史的前提,除受種子自身大小、質量等因素影響外,還受制于土壤養分等環境因素(Rajjou et al.,2012)。鹽漬化土中的植物種子萌發和正常生長均十分困難(齊琪等,2020),在鹽漬化和低養分惡劣生境中,種子能夠萌發成苗是植物適應此類惡劣環境的表現(張建鋒等,2003;沈振榮等,2006)。然而,鹽脅迫在一定程度上抑制了植物種子的萌發及生長,植物種子的活力指數、發芽指數、發芽勢等萌發指標及幼苗生長均隨著鹽濃度的升高呈降低趨勢
(李云等,1997;沈振榮等,2006; 韓廣軒等,2009; 盧艷敏,2012)。此外,磷含量也是影響種子萌發和幼苗生長的重要因素。農業上重視抗低磷基因和品種等研究(呂春雨和沙愛華,2017),林業上有學者發現低濃度磷添加可以促進種子萌發及幼苗生長,而高濃度磷有一定的抑制作用(唐敏,2007),不同磷源會對種子萌發等產生影響(戴開結等,2009)。中國鹽堿地總面積超過 9 913 萬 hm2,其中濱海鹽堿地約占 842.6萬 hm2(朱建峰等,2018),中國東部及南部沿海地區土壤除鹽漬化外,全磷和有效磷含量均較低(汪濤等,2008),只有少數樹種能存活下來,由它們組成的沿海海防林構成了沿海地區的重要生態安全屏障。因此,研究海防林主干植物種子和幼苗對土壤鹽堿化和低磷等惡劣環境的適應性具有重要意義。雖然抗鹽植物可以通過形態、生理和生化調節提高自身對單一惡劣環境的適應能力,但多因素的疊加效應可能大幅度限制或降低這種能力。因此,開展海防林樹種抵抗鹽漬和低磷的閾值以及二者同時作用對海防林植物種子萌發和幼苗生長影響程度的研究,將有助于探索和揭示海防林天然更新的機制和提高海防林經營管理水平,特別是對海防林的主要樹種更是如此。
木麻黃(Casuarina equisetifolia)是木麻黃科(Casuarinaceae)木麻黃屬(Casuarina Adans.)植物,20世紀50年代大規模引種到中國東南沿海地區,成為主要防護林樹種(袁鋒等,2020)。臺灣相思(Acacia confusa)是豆科(Leguminosae)金合歡屬(Acacia)速生樹種,具有根系發達、抗風、萌芽力強、耐干旱貧瘠等特點,與木麻黃相似,均能適應惡劣的海岸環境。因此,生產上常將二者進行混植(陳青霞,2011)。木麻黃種子萌發研究表明水分(王玉等,2020)、高濃度鹽脅迫(武沖等,2010)、鉻脅迫(周希群和李裕紅,2004)、鎘脅迫及鉛脅迫(陳懷宇等,2007)均會抑制種子的萌發及生長,而磷脅迫對木麻黃種子萌發影響及鹽、磷脅迫對臺灣相思種子萌發和生長影響的研究未見報道,同時鹽、磷脅迫對兩種植物種子萌發的影響閾值還未確定。因此,為探討海防林植物在種子萌發和萌發后的幼苗生長這兩個脆弱期對高鹽低磷的適應能力,本研究開展不同濃度鹽磷脅迫對木麻黃、臺灣相思的種子萌發及幼苗生長影響實驗,確定影響萌發閾值以及影響幼苗生長的指標,以期為海防護林植被更新及可持續經營提供理論參考。
1 材料與方法
1.1 材料
木麻黃、臺灣相思的種子由福建省惠安赤湖國有防護林場提供,晾曬后選取顆粒飽滿的種子于2020年11月進行萌發實驗,木麻黃的千粒重、縱徑、橫徑分別為(0.43±0.07) g、(3.74±0.13) mm、(1.58±0.08) mm,臺灣相思的分別為(32.3±1.31) g、(5.86±0.21) mm、(4.21±0.22) mm。采用TTC法(劉有軍等,2019)測定種子活力,木麻黃、臺灣相思的種子活力分別在60%、80%以上。
1.2 實驗設計
實驗在人工氣候箱(RTOP-500Y)中進行,設置恒溫25 ℃、濕度75%、光照強度2 000 μmol·m-2·s-1(晝夜各12 h)。采用砂培法進行種子實驗,在培養皿(直徑14.5 cm)內放置石英砂(粒徑 ≤ 0.5 mm、厚度1 cm),使用前用鹽酸(5%)浸泡1 h后用清水沖洗干凈,恒溫烘干。為確定鹽、磷影響閾值,分別用NaCl和KH2PO4配置成0%(CK)、0.2%、0.4%、0.6%、0.8%、1 %和0(CK)、0.1、0.5、1、5、10 mmol·L-1濃度的溶液(曲東等,1998;韓廣軒等,2009)。對種子用KMnO4(濃度1%)浸泡20 min消毒,用蒸餾水洗凈后,木麻黃用45 ℃溫水,臺灣相思用70 ℃熱水浸種,自然冷卻后用相應濃度的溶液浸種24 h后播種。每個培養皿中均勻放木麻黃種子50粒、臺灣相思種子30粒,每個處理3個重復,用相應濃度的溶液浸透石英砂,后續定期測定質量,并用蒸餾水補充流失的水分以保持恒重。以培根漏出種皮超過種子長度作為種子萌發的標準(王玉等,2020),每天記錄種子的萌發數,連續5 d無種子萌發后結束實驗,木麻黃、臺灣相思分別于14、10 d結束實驗。
1.3 指標測定
木麻黃、臺灣相思每個處理分別選取5、10棵測定其株高、根長及根莖葉鮮重。種子萌發及生長指標計算公式(張大鵬等,2008;劉有軍等,2019):發芽率=(發芽種子數/供試種子數)×100%;發芽勢=(種子萌發高峰期種子數/供試種子數)×100%;發芽指數=∑Gt/Dt,式中,Gt為在t天的發芽數,Dt為發芽天數;活力指數=鮮重×發芽指數;相對鹽害率=(對照發芽率-處理發芽率)/對照發芽率×100%;根冠比=地下鮮重/地上鮮重。
1.4 數據處理
采用Excel、SPSS 25軟件進行數據處理,在單因素方差分析的基礎上,利用最小顯著差異法進行處理間差異顯著性檢驗,利用雙因素方差分析鹽、磷交互作用對種子萌發及生長的影響,采用Origin 2018軟件進行作圖。
2 結果與分析
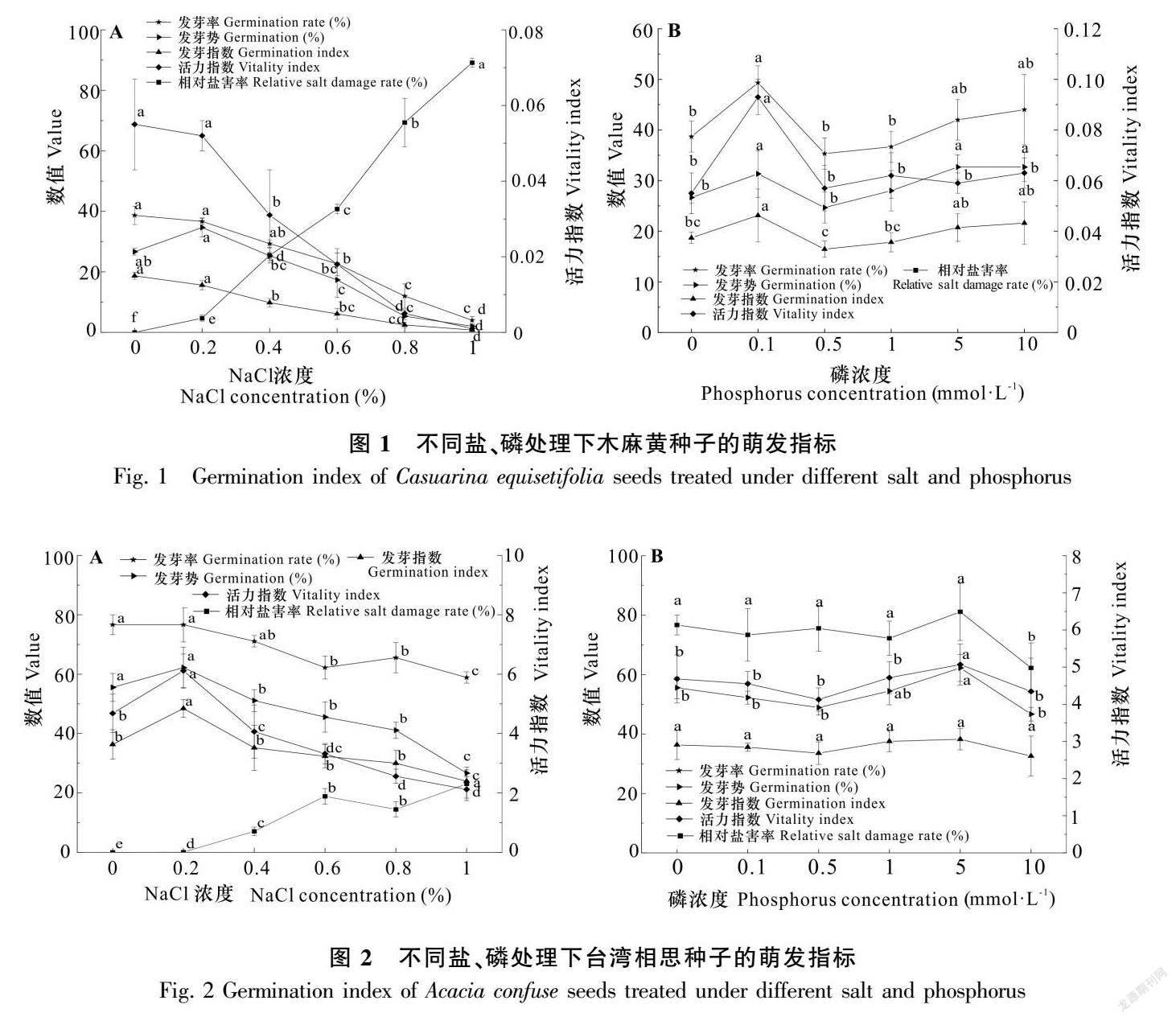
2.1 鹽、磷脅迫對種子萌發及存活的影響
NaCl 處理下木麻黃種子由0%~1%濃度首次萌發時間逐漸延長,由第3天延遲到第6天,萌發高峰期相應由第3天延遲到第9天,1%處理萌發率極低。臺灣相思首次萌發時間均在第1天,萌發高峰期集中在第3天。磷處理下木麻黃種子均在第3天萌發,高峰期均在第4天,臺灣相思首次萌發均在第1天,高峰期在第2天或第3天。
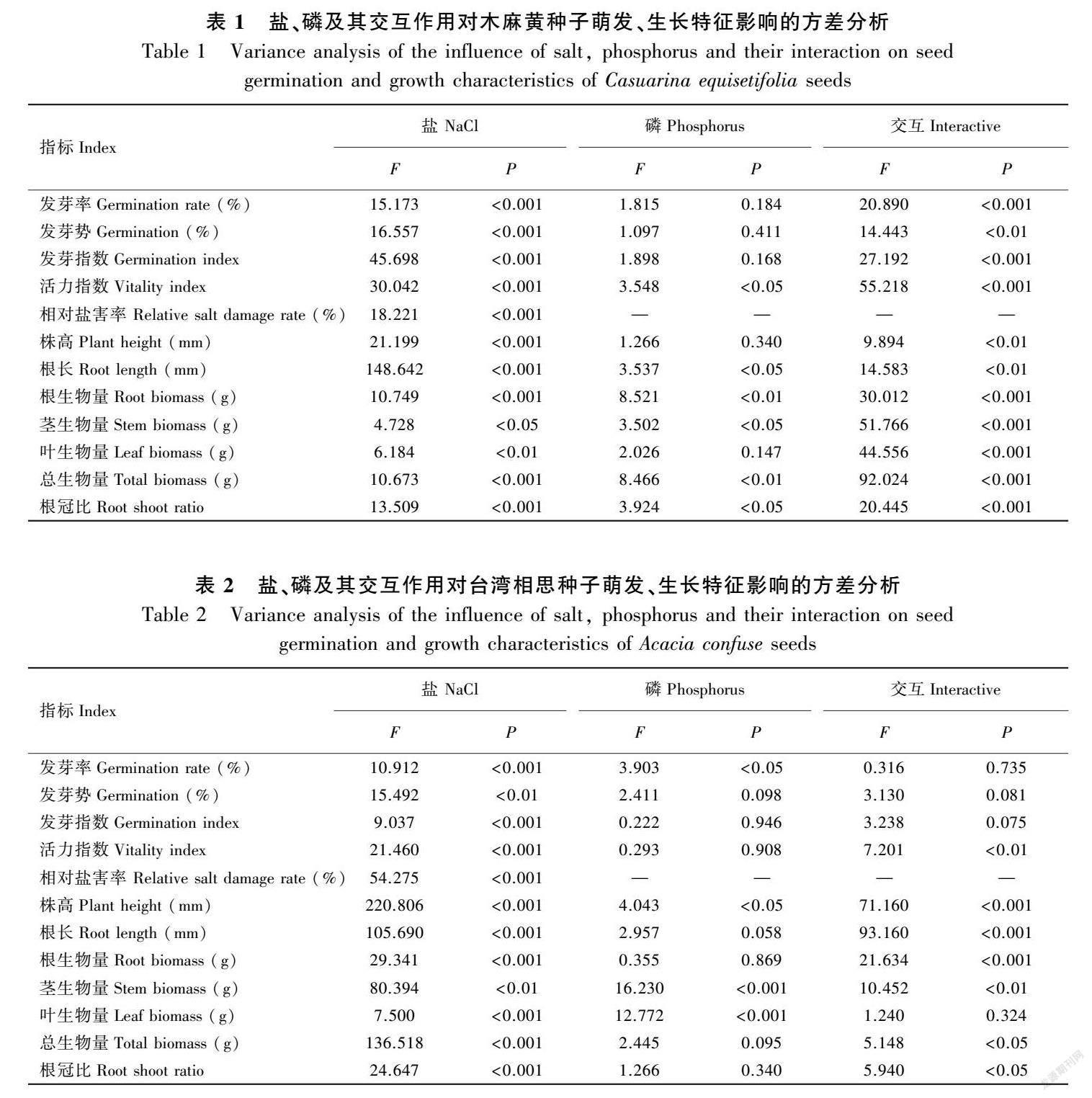
高NaCl濃度對種子萌發產生顯著抑制作用(圖1,圖2,表1,表2)。木麻黃種子隨著NaCl濃度增加,發芽率由38.67%降到4.00 %,發芽指數由18.70降到0.78,活力指數由0.055降到0.001,相對鹽害率由4.66%增加到89.15%,發芽勢由0%(26.67%)增加到0.2%(34.67%)后逐漸降到2.00%。總體上,0%和0.2%處理間差異不顯著,其他處理間差異顯著(P<0.01)。0.2%NaCl濃度對臺灣相思種子萌發具有促進作用,萌發指標均為最大值且顯著高于0%,發芽率、發芽勢、發芽指數、活力指數分別為76.67%、62.22%、48.46、6.11;NaCl濃度大于0.2%后,萌發指標開始下降,相對鹽害率不斷增加,最大鹽害率為23.03%。
不同磷濃度處理對種子萌發影響存在差異, 適度磷添加具有促進作用(圖1,圖2)。隨著磷濃度增加,萌發指標表現為先增加后降低再增加的趨勢,濃度為0.1 mmol·L-1時,木麻黃種子發芽率(49.33%)、發芽指數(23.12%)和活力指數(0.093)均最大,發芽勢最大值(32.67%)位于5、10 mmol·L-1處理。臺灣相思種子發芽率(81.11%)、發芽勢(62.22%)、發芽指數(38.23)和活力指數(5.07)最大值均位于5 mmol·L-1處理。從總體來看,前5個處理差異不顯著,10 mmol·L-1處理萌發指標明顯低于0 mmol·L-1(圖2),與其他處理差異顯著(P<0.01),說明等于及大于此處理后對種子萌發具有抑制作用。方差分析表明,磷對臺灣相思種子萌發影響較小,鹽、磷雙因素對木麻黃種子萌發影響顯著,對臺灣相思種子影響較小(表1,表2)。
2.2 鹽、磷脅迫對幼苗株高、根長的影響
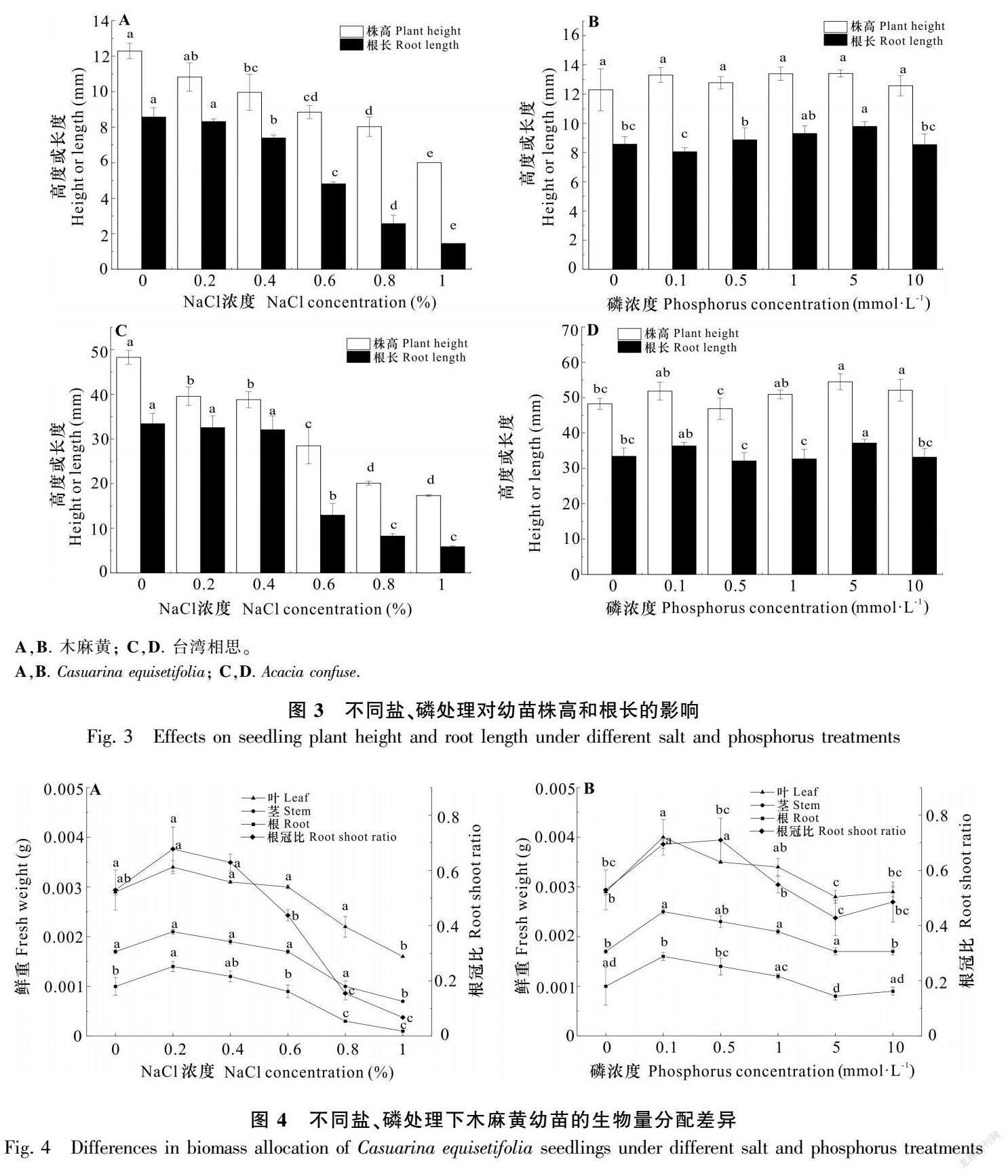
不同NaCl濃度處理對幼苗株高和根生長表現為顯著抑制作用。隨著濃度增加株高及根長均逐漸降低,木麻黃株高由12.29 mm降到6.01 mm,根長由8.57 mm降到1.45 mm,0%和0.2%處理間差異不顯著;臺灣相思株高由48.27 mm降到17.33 mm,根長由33.41 mm降到5.88 mm,根長0%、0.2%、0.4%處理間無顯著差異(圖3,表1,表2)。
磷處理對木麻黃株高影響不顯著,對根長影響顯著(圖3,表1),株高、根長最大值位于5 mmol·L-1處理,分別為13.41、9.79 mm。磷處理對臺灣相思株高影響顯著,對根長影響不顯著(圖3,表2),株高、根長最大值位于5 mmol·L-1處理,分別為54.48、37.16 mm。
2.3 鹽、磷脅迫對幼苗生物量分配的影響
不同NaCl濃度處理木麻黃根生物量0.2%(0.001 4 g)、0.4%(0.001 2 g)處理高于CK(0.001 0 g),大于0.4%后逐漸降低且均低于CK,莖、葉生物量各處理差異較小,莖生物量為0.000 6~0.000 8 g,葉生物量為0.000 9~0.001 3 g,總生物量大于0.6%后逐漸降低,根冠比0.2%(0.68)、0.4%(0.63)處理同樣高于0%(0.53),其他處理均顯著低于CK,1%處理為0.07,根、葉生物量和根冠比最大值位于0.2%處理(圖4)。臺灣相思的根、莖、葉生物量及根冠比均隨NaCl濃度增加逐漸減小,0%、0.2%、0.4%處理間差異較小,濃度大于0.4%時各指標明顯下降,表明0.4%為閾值(圖5)。
隨著磷濃度增加木麻黃根生物量和根冠比均呈先增加后降低的趨勢(圖4),最大值為0.001 6 g、0.70。莖生物量為0.000 7~0.000 9 g,葉生物量為0.001 1~0.001 5 g,莖、葉生物量各處理間差異不顯著,根、莖、葉生物量的最大值位于0.1 mmol·L-1處理。總體上,磷對臺灣相思幼苗生物量分配影響較小。臺灣相思根生物量為0.017~0.019 g,處理間差異不顯著;莖生物量為0.028~0.039 g,葉生物量為0.072~0.078 g,根冠比為0.15~0.18。方差分析發現,鹽、磷交互作用對根、莖生物量及根冠比的影響顯著(表1,表2)。
3 討論與結論
種子保持活力及幼苗正常生長是植物在高鹽環境下存活的關鍵。相關研究表明,滲透、離子效應是鹽脅迫下影響種子萌發及生長的主要因素,主要表現為種子的萌發速度變慢、萌發率降低以及延緩幼苗的生長等,高鹽甚至導致種子失活或幼苗死亡(齊琪等,2020)。本研究發現,高鹽環境對木麻黃、臺灣相思的種子萌發及生長抑制作用明顯,發芽率、發芽勢、發芽指數、活力指數、株高、根長、生物量及根冠比均隨NaCl濃度升高而下降,這與大多數研究結果一致(李云等,1997;韓廣軒等,2009;武沖等,2010),但鹽脅迫下種子發芽率也可能與其品種有關(武沖等,2010)。木麻黃發芽率、發芽指數、活力指數、株高、根長、根和葉生物量最大值均位于0%,根冠比、發芽勢均位于0.2%處理,各指標0%與0.2%之間差異不顯著,其他處理間差異顯著,表明低于0.2%處理對木麻黃種子萌發影響較小,1%處理下種子基本不萌發,王玉等(2020)對木麻黃種子進行0~0.1%NaCl濃度脅迫同樣得出無顯著影響的結論。本研究還發現木麻黃種子的耐鹽性低于成苗的耐鹽性,蘇祖榮(1999)研究表明1 a生幼苗在0.6%濃度鹽脅迫下造林成活率大于60%,耐鹽率為0.767%,成林最大耐鹽率為3.141%。本研究中,臺灣相思種子萌發指標最大值均為0.2%濃度且明顯大于CK,生長指標株高、根長、根冠比及根和莖生物量最大值均位于CK,但0%、0.2%、0.4%處理之間差異性較小,萌發及生長指標表明0.2%NaCl濃度有助于臺灣相思種子的萌發,原因可能為此濃度下離子效應為正效應(齊琪等,2020),當濃度大于0.4%后各指標下降,與CK間差異顯著,說明0.4%濃度為臺灣相思種子萌發的閾值。本研究從相對鹽害率來看,木麻黃種子在4.66%~89.15%之間,臺灣相思種子在0.07%~23.03%之間,鹽脅迫對臺灣相思種子萌發的影響明顯低于木麻黃,臺灣相思種子萌發的耐鹽性更強,原因可能為臺灣相思種子有一層堅硬的外殼。
在不同NaCl濃度處理下,根生物量變化幅度遠大于莖葉生物量,并且隨著濃度增加根冠比逐漸減小,根長同樣逐漸變短,表明鹽脅迫對根的生長影響大,這與韓廣軒等(2009)對黑松研究結果一致。Jose等(2017)研究認為耐NaCl的植物通過形態學、生理學和生化等方面的調節來適應鹽度,防止鹽離子毒性以維持水的狀態,從而達到限制水分流失等目的,如改變根冠比,但上述調節要在一定范圍內。
鹽影響海防林兩種植物種子萌發及生長的原因主要包括以下方面:第一,不同NaCl濃度造成種子周圍水勢及細胞膜損害程度差異,水勢影響種子吸水,濃度越大水勢越低,種子內外水勢差增大,種子吸水不足,進而抑制呼吸作用及物質運輸,導致萌發困難(王娟娟和張文輝,2011;齊琪等,2020)。第二,細胞膜損害改變通透性,造成離子(K+、Na+等)含量、植物體內的保護酶系統中各種酶含量(SOD、POD、CAT等)及MDA含量相應存在差異,進而影響種子萌發及生長(張大鵬等,2008;王娟娟和張文輝,2011;陳意蘭等,2021)。第三,離子應力和質膜電位改變影響種子萌發和生長。Assaha等(2017)研究認為離子應力是鹽度中最重要的組成部分之一,是由過量的Na+積累引起的,Na+干擾K+穩態,兩者失衡進而引起質膜(PM)電位變化等差異。
本研究結果表明,低、中濃度的磷添加對種子的萌發及生長影響較小或有促進作用,高磷有一定的抑制作用,這與前人(唐敏,2007;宋紅麗等,2012)研究結果基本一致。木麻黃種子萌發指標在0.1 mmol·L-1磷處理均最高,其次為5 mmol·L-1,生長指標中株高、根長的最大值均在5 mmol·L-1處理,根、莖、葉生物量及根冠比的最大值均在0.1 mmol·L-1磷處理,表明0.1、0.5 mmol·L-1磷處理對木麻黃種子萌發及生長具有促進作用。臺灣相思種子萌發指標在前5個處理差異不顯著,10 mmol·L-1磷處理萌發指標明顯低于CK且與其他處理差異顯著,最大值在5 mmol·L-1磷處理。株高、根長最大值均在5 mmol·L-1磷處理,根、莖、葉生物量以及根冠比各處理之間差異不顯著,表明磷對臺灣相思種子萌發及生長影響較小,5 mmol·L-1磷處理對其萌發和生長有一定的促進作用,同時10 mmol·L-1磷處理萌發指標明顯低于CK且與其他處理差異顯著,表明高磷對種子萌發及生長具有抑制作用,這與唐敏(2007)對馬尾松(Pinus massoniana )研究結果一致。
種子萌發受磷影響的原因可能為磷添加提高了淀粉酶活性(曲東等,1998),或者用低、中濃度磷浸種的種子電導率降低,有助于種子膜結構修復(唐敏,2007)。高磷脅迫下,種子電導率升高,破壞膜結構的完整性,對種子造成傷害,并且極低磷和高磷的逆境環境均使得SOD、POD及CAT等抗逆性酶活性及MDA含量升高。劉慶艷等(2013)研究表明隨著磷養分由低-中-高濃度的輸入,興凱湖沼澤土壤種子庫種子萌發數為減小-增加-顯著減小的變化趨勢,并且不同植物對磷養分適應性存在差異,適量的磷添加有助于植物種子萌發。植物有限的吸收表面積可能是高磷抵制生長的重要原因,過量的磷包圍、改變細胞內外磷離子濃度、占據膜表面電子位,極可能影響其他元素的吸收,進而影響幼苗的生長。因此,即使種子萌發對磷的依賴度低,幼苗生長也需要補充適量的磷元素,而過量則起反作用。
本研究發現,鹽、磷交互作用對種子萌發及生長可能產生一定的影響,而此類研究目前卻較少。宋紅麗等(2012)研究表明,鹽脅迫下添加磷對堿蓬(Suaeda glauca)種子萌發及生長影響復雜,但在高鹽脅迫下,添加低濃度磷卻有利于堿蓬種子萌發及生長,原因可能是鹽脅迫改變種子和根系細胞膜表面離子濃度和電子平衡環境,進而影響植物對磷元素的吸收,而磷元素是植物光合作用中光合磷酸化所必需的營養物質,阻礙或抵制了光合作用,進而影響植物的初級生產力及分配(宋紅麗等,2012;李玲,2015)。鹽磷互作對種子萌發及生長的報道較少,其對種子萌發、幼苗生長影響及生理機制還需進一步研究。
綜上所述,高鹽脅迫對木麻黃種子萌發表現為顯著抑制甚至完全抑制作用,0.2%NaCl處理促進臺灣相思種子萌發及生長,0.6%、0.4% NaCl分別為木麻黃、臺灣相思的種子萌發及生長的閾值,其中鹽脅迫對根的生長影響顯著,隨著NaCl濃度的增加,根生物量、長度、根冠比等指標逐漸降低。0.1 mmol·L-1磷處理對木麻黃種子萌發及生長存在促進作用,低、中濃度磷處理對臺灣相思種子萌發及生長影響不顯著,10 mmol·L-1磷處理對臺灣相思種子萌發及生長存在抑制作用。同時,木麻黃種子的耐鹽性低于成苗,在采用種子進行種植時應檢測土壤鹽、磷的含量,木麻黃、臺灣相思的鹽度應分別在0.6%、0.4%范圍內,木麻黃可施加適量的磷肥,臺灣相思不施加。
參考文獻:
ASSAHA DVM, UEDA A, SANEOKA H, et al., 2017. The Role of Na+ and K+ transporters in salt stress adaptation in glycophytes [J]. Front Physiol, 8: 509.
CHEN HY, LI YH, LIN YC, 2007. Effects of Cd2+ and Pb2+ on seed germination and fastness physiological characteristics of Casuarina equisetifolia [J]. Chin Agric Sci Bull, 23(8): 229-232. [陳懷宇, 李裕紅, 林元燦, 2007. Cd2+、Pb2+對木麻黃種子萌發及抗性生理特性的影響 [J]. 中國農學通報, 23(8): 229-232.]
CHEN QX, 2011. Study on mixed forest of Casuarina equisetifolia and Acacia taiwanensis in sandy land of Huian County [J]. Anhui Agri Sci Bull, 17(9): 158-159. [陳青霞, 2011. 惠安縣沙地木麻黃臺灣相思混交林研究 [J]. 安徽農學通報, 17(9): 158-159.]
CHEN YL, LI X, ZHAO WZ, et al., 2021. Physiological response of Thuarea involuta under salt stress [J]. Guihaia, 41(2): 225-232.[陳意蘭, 李昕, 趙文忠, 等, 2021. 蒭雷草對鹽脅迫的生理響應 [J]. 廣西植物, 41(2): 225-232.]
DAI KJ, HE F, SHEN YX, et al., 2009. Effects of different phosphorus on growth and P absorption of Pinus yunnanensis Franch. seedlings[J]. Acta Ecol Sin, 29(8):4078-4083.[戴開結, 何方, 沈有信, 等, 2009. 不同磷源對云南松幼苗生長和磷吸收量的影響 [J]. 生態學報, 29(8):4078-4083.]
HAN GX, MAO PL, LIU SJ, et al., 2009. Effects of sea water salinity and mother tree size on the seed germination and seedling early growth of Pinus thunbergii coastal protection forest [J]. Chin J Ecol, 28(11): 2171-2176.[韓廣軒, 毛培利, 劉蘇靜, 等, 2009. 鹽分和母樹大小對黑松海防林種子萌發和幼苗早期生長的影響 [J]. 生態學雜志, 28(11): 2171-2176.]
JOSE RA, MARIA FO, AGUSTINA B, et al., 2017. Plant responses to salt stress: adaptive mechanisms [J]. Agronomy, 7(1): 1-38.
LI Y, ZHENG DZ, LIAO BW, et al., 1997. Effect of salinity and temperature on seed germination of mangrove Sonneratia apetala Buch. Ham. [J]. For Res, 10(2): 30-35. [李云, 鄭德璋, 廖寶文, 等, 1997. 鹽度與溫度對紅樹植物無瓣海桑種子發芽的影響 [J]. 林業科學研究, 10(2): 30-35.]
L CY, SHA AH, 2017. Response to phosphorus deficiency regulated by microRNA168 in soybean plant [J]. Chin J Oil Crop Sci, 39(3): 321-325. [呂春雨, 沙愛華, 2017. 大豆microRNA168調控植物低磷脅迫響應 [J]. 中國油料作物學報, 39(3): 321-325.]
LU YM, 2012. Effects of different saline stress on seed germination of tall fescue [J]. Pratac Sci, 29(7): 1088-1093.[盧艷敏, 2012. 不同鹽脅迫對高羊茅種子萌發的影響 [J]. 草業科學, 29(7): 1088-1093.]
LIU YJ, LIU SZ, KANG CZ, et al., 2019. Comparative adaptation of seed germination and seedling growth to environmental factors in two Picea plant species [J]. Acta Ecol Sin, 39(2): 611-619. [劉有軍, 劉世增, 康才周, 等, 2019. 兩種云杉種子萌發和幼苗生長對環境因子的適應性 [J]. 生態學報, 39(2): 611-619.]
LIU QY, JIANG M, WANG GD, et al., 2013. Effect of exogenous phosphorus inputs on seed germination of soil seed bank in marshes in Xingkai Lake [J]. Wetl Sci, 11(1): 41-47. [劉慶艷, 姜明, 王國棟, 等, 2013. 外源磷輸入對興凱湖沼澤土壤種子庫種子萌發的影響 [J]. 濕地科學, 11(1): 41-47.]
LI L, 2015. Effects of different phosphorus levels on the growth of Kosteletzky virginica under NaCl stress [D].Jinan: Shandong Normal University: 31-41. [李玲, 2015. 不同磷素水平對鹽脅迫下海濱錦葵生長的影響 [D]. 濟南: 山東師范大學: 31-41.]
QI Q, MA SR, XU WD, 2020. Advances in the effects of salt stress on plant growth and physiological mechanisms of salt tolerance [J]. Mol Plant Breed, 18(8): 2741-2746. [齊琪, 馬書榮, 徐維東, 2020. 鹽脅迫對植物生長的影響及耐鹽生理機制研究進展 [J]. 分子植物育種, 18(8): 2741-2746.]
QU D, WANG BL, BAI HY, et al., 1998. Effect of seed pretreatment with phosphorus on seed germination and seedling growth [J]. Acta Univ Agric Boreal-occident, 26(1): 8-12. [曲東, 王保莉, 白紅英, 等, 1998. 磷浸種對種子萌發和生長的影響 [J]. 西北農業大學學報, 26(1): 8-12.]
RAJJOU L, DUVAL M, GALLARDO K, et al., 2012. Seed germination and vigor [J]. Ann Rev Plant Biol, 63: 507-533.
SHEN ZR, YANG WR, XU XM, 2006. Effect of salt stress on germination of alfalfa seeds Fengmei [J]. Seed, 25(4): 34-37.[沈振榮, 楊萬仁, 徐秀梅, 2006. 不同鹽分脅迫對苜蓿種子萌發的影響 [J]. 種子, 25(4): 34-37.]
SONG HL, SUN ZG, SUN JK, et al., 2012. Effects of nitrogen and phosphorus on seed germination and seedling growth of Suaeda salsa under different growth conditions of the Yellow River Estuary [J]. Acta Pratac Sin, 21(6): 30-41.[宋紅麗, 孫志高, 孫景寬, 等, 2012. 氮、磷輸入對黃河口潮灘濕地不同生境下堿蓬種子萌發與幼苗生長的影響 [J]. 草業學報, 21(6): 30-41.]
SU ZR, 1999. Experimental study on salt tolerance of Casuarina equisetifolia and salt reduction technology [J]. Prot For Sci Technol, 38(1): 17-19.[蘇祖榮, 1999. 木麻黃抗鹽能力及降鹽技術試驗研究 [J]. 防護林科技, 38(1): 17-19.]
TANG M, 2007. Study on effect of phosphorus to germination and seedling of different provenances [D]. Guiyang: Guizhou University: 17-25.[唐敏, 2007. 磷對不同種源馬尾松種子及幼苗影響的研究 [D]. 貴陽: 貴州大學: 17-25.]
WANG T, YANG YH, MA WH, 2008. Storage, patterns and environmental controls of soil phosphorus in China [J]. Acta Sci Nat Univ Pekin, 44(6): 945-952.[汪濤, 楊元合, 馬文紅, 2008. 中國土壤磷庫的大小、分布及其影響因素 [J].北京大學學報(自然科學版), 44(6): 945-952.]
WANG JJ, ZHANG WH, 2011. Effects of activities of protective enzymes and seed germination in Atriplex canescens under NaCl and Na2CO3 stress [J]. Sci Silv Sin,47(2):154-160. [王娟娟, 張文輝, 2011. NaCl和Na2CO3脅迫對四翅濱藜種子萌發及保護酶活性的影響 [J]. 林業科學, 47(2): 154-160.]
WU C, ZHANG Y, TANG SM, et al., 2010. Effect of NaCl stress on Casuarina seed germination [J]. Seed, 29(4): 30-33.[武沖, 張勇, 唐樹梅, 等, 2010. 鹽脅迫對木麻黃種子萌發的影響 [J]. 種子, 9(4): 30-33.]
WANG Y, YANG B, HAO QY, 2020. Limiting ecological factors for seed germination of Casuarina equisetifolia [J]. Guihaia, 40(3): 403-411.[王玉, 楊彬, 郝清玉, 2020. 木麻黃種子萌發的限制生態因子 [J]. 廣西植物, 40(3): 403-411.]
YUAN F, WANG YY, LI MJ, et al., 2020. Dynamic characteristics of metal element content and return of Casuarina equisetifolia litter at different distances to the coastline [J]. Chin J Plant Ecol, 44(8): 819-827.[袁鋒, 王艷艷, 李茂瑾, 等, 2020. 不同海岸距離上木麻黃凋落葉金屬元素含量及歸還量動態特征 [J]. 植物生態學報, 44(8): 819-827.]
ZHU JF, CUI ZR,WU CH, et al., 2018. Research advances and prospect of saline and alkali land greening in China [J]. World For Res, 31(4): 70-75. [朱建峰, 崔振榮, 吳春紅, 等, 2018. 我國鹽堿地綠化研究進展與展望 [J]. 世界林業研究, 31(4): 70-75.]
ZHANG DP, CAO BH, JIA B, et al., 2008. Germination and physiological response of Albizia julibrissin seeds under alkali-salt stress [J]. Sci Silv Sin, 44(9): 157-161.[張大鵬, 曹幫華, 賈波, 等, 2008. 鹽堿脅迫下合歡種子的抗逆萌發生理 [J]. 林業科學, 44(9): 157-161.]
ZHANG JF, LI JY, XING SJ, et al., 2003. Seed germination of Chionanthus retusa and Cedrela sinensis under salinity stress [J]. J Beijing For Univ, 25(4): 80-83.[張建鋒, 李吉躍, 邢尚軍, 等, 2003. 流蘇和香椿種子在鹽分脅迫下的發芽研究 [J]. 北京林業大學學報, 25(4): 80-83.]
ZHOU XQ, LI YH, 2004. The physiological and ecological responses of the seed germination of Casuarina equisetifolia to chromic stress [J]. Chin J Eco-Agric,12(1): 58-60.[周希琴, 李裕紅, 2004. 木麻黃種子萌發對鉻脅迫的生理生態響應研究 [J]. 中國生態農業學報, 12(1): 58-60.]
(責任編輯 蔣巧媛 鄧斯麗)
