葡萄酒中鏈脂肪酸乙酯產生機制與調控策略探討
2023-05-25 08:20:52胡文效姚彬彬陳明光朱宇彤沃曉妍邱磊
中外葡萄與葡萄酒 2023年3期
關鍵詞:酵母菌
胡文效,姚彬彬,陳明光,朱宇彤,沃曉妍,邱磊*
(1. 齊魯工業大學(山東省科學院),山東濟南 250353;2. 山東中煙工業有限責任公司,山東濟南 250013)
中鏈脂肪酸乙酯(MCFAEE),如己酸乙酯、辛酸乙酯和癸酸乙酯,具有漿果、花香香韻,對提高葡萄酒感官品質發揮著關鍵作用。該類物質是葡萄酒香氣的重要組成部分,普遍存在于葡萄酒、白蘭地、威士忌、朗姆酒等飲料酒中[1-2]。湯曉宏等[3]的研究發現,‘赤霞珠’果實中未檢出C8、C10的MCFAEE化合物,而對其葡萄酒的香氣化合物分析顯示,酯類的種類和含量最豐富,其中辛酸乙酯、癸酸乙酯相對含量顯著高于其它酯類;劉靜等[4]分析了3個年份‘維歐尼’葡萄酒的香氣成分,得出類似結果,即MCFAEE相對含量較高,且辛酸乙酯顯著高于其它酯類化合物;前蘇聯高格察西維里[5]從葡萄酒酵母沉淀物中分離出的康釀克油,其主要成分是癸酸乙酯、辛酸乙酯、月桂酸乙酯等;李記明等[6]分析鑒定了XO級白蘭地揮發性成分,結果表明酯類是白蘭地中主要的揮發性成分,其中辛酸乙酯、癸酸乙酯、十二酸乙酯相對含量較高。
葡萄漿果中脂肪族揮發性化合物的組成主要為直鏈或支鏈的短鏈C6醛和醇[7],葡萄中存在的所能檢測到的酯類脂肪族化合物通常處于較低豐度,且對香氣的影響較小[8],說明葡萄酒中MCFAEE是在葡萄酒發酵過程中產生。有研究指出,在厭氧發酵過程中,釀酒酵母會釋放大量的中鏈脂肪酸(MCFA)和相關的乙酯,對于發酵飲料的芳香品質非常重要[9]。進一步說明發酵過程中乙酯是通過脂質代謝產生[10]。本文通過對已有文獻分析MCFAEE的生成機制,探討MCFAEE是來源于脂質合成代謝或者分解代謝,以及調控環境變量對MCFAEE產量的影響。
1 釀酒酵母脂肪酸代謝
葡萄酒發酵過程中,酵母菌生長經遲緩期、對數期、穩定期、衰亡期,直至發酵停止,一般需要7~14 d,期間持續進行復雜的物質代謝和能量代謝。物質代謝的結果除產生酵母菌體和主產物乙醇外,還有眾多副產物,共同構成葡萄酒發酵產物。葡萄酒發酵中酵母菌生長對數后期至穩定期,生物量一般達到108cfu·mL-1。酵母菌生長繁殖需要相應量的長鏈脂肪酸(LCFA)合成特定的磷酸甘油酯(1, 2-二酯酰-L-甘油-3-磷酸),構建細胞膜系統。釀酒酵母菌的脂肪酸主要由飽和脂肪酸棕櫚酸(C16)、硬脂酸(C18)和不飽和脂肪酸油酸(C18:1)、棕櫚油酸(C16:1)組成[11]。因菌株和發酵基質、有氧或厭氧等變量差異,脂肪酸組成有所不同。陳聲明等[12]對釀酒酵母菌脂肪酸組成分析結果顯示,C16:1占59.6%、C18:1占24.73%、C16:0占6.9%。
1.1 脂肪酸分解代謝
葡萄汁中含有一定量的脂質。段亮亮[13]測得‘赤霞珠’葡萄果漿總脂肪酸含量為755.33 mg·kg-1,其中,亞油酸316.37 mg·kg-1、軟脂酸127.90 mg·kg-1、α-亞麻酸81.60 mg·kg-1、油酸64.62 mg·kg-1、硬脂酸44.45 mg·kg-1。在葡萄酒發酵過程中,葡萄汁中的脂肪酸經過相應機制進入酵母細胞被用于脂質合成或作為能量來源,如以三酰基甘油形式存在的脂肪酸,在釀酒酵母胞外酰基甘油酯酶作用下水解產生游離脂肪酸,并通過擴散或轉運蛋白被吸收進入酵母細胞。在定位于內質網、脂質滴的酰基輔酶A合成酶作用下,進入細胞質的外源性脂肪酸與輔酶A結合生成脂酰輔酶A,產物脂酰輔酶A可以用于細胞復雜的脂質合成或在過氧化物酶體中進行β-氧化[14]。釀酒酵母中脂肪酸的β-氧化作用僅限于細胞的過氧化物酶體區室[15-16],如圖1。
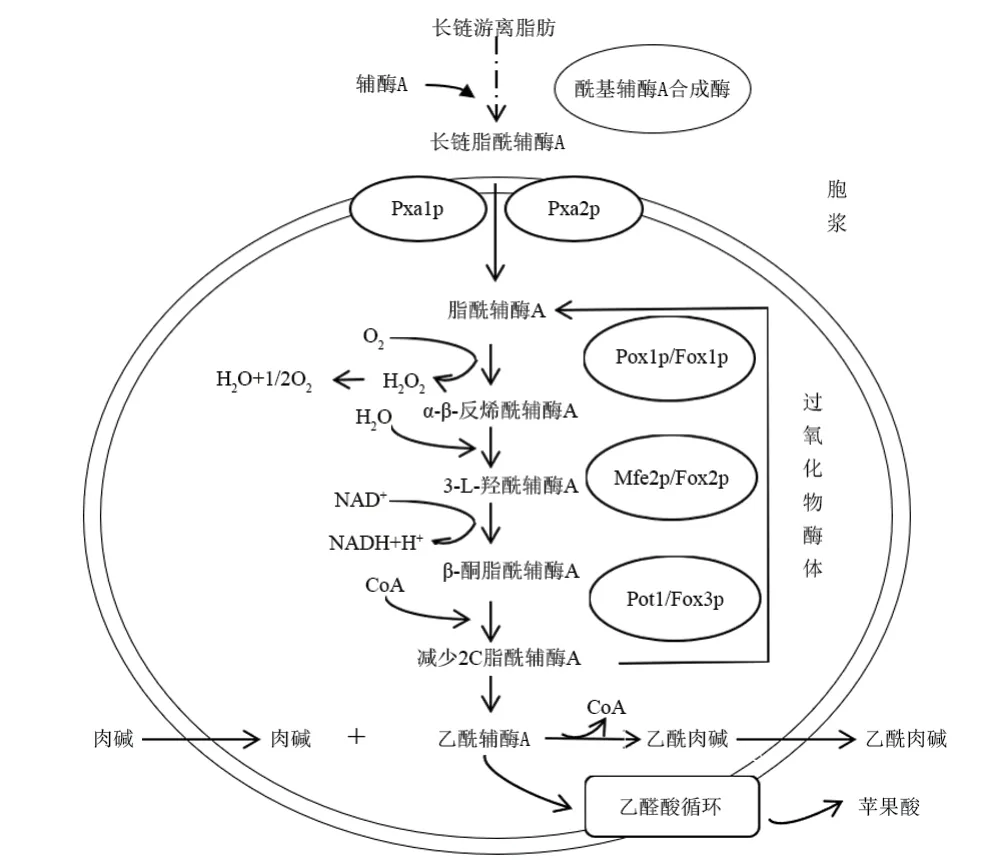
圖1 葡萄酒發酵釀酒酵母過氧化物酶體長鏈脂肪酸氧化示意圖Figure 1 Schematic diagram of long-chain fatty acid oxidation in wine fermentation by Saccharomyces cerevisiae
長鏈脂酰輔酶A(LCFA-CoA)通過過氧化物酶體膜轉運蛋白Pxa1p和Pxa2p導入過氧化物酶體[17]。β-氧化過程首先是酰基輔酶A氧化酶Fox1p(Pox1p)將脂酰輔酶A氧化為反式2-烯酰輔酶A;反式-2-烯酰輔酶A在Fox2p(Mfe2p)作用下,經水化、脫氫轉化為β-酮脂酰輔酶A;硫解酶Fox3p(Pot1p)催化β-酮脂酰輔酶A裂解為乙酰輔酶A和減少2個碳的脂酰基-CoA[18]。過氧化物酶體內產生的乙酰輔酶A進入胞質或線粒體有兩條途徑,即通過轉化為乙醛酸循環中間體或通過肉堿乙酰轉移酶轉化為乙酰肉堿[19]。肉堿乙酰基轉移酶催化肉堿與乙酰輔酶A之間的可逆反應,從而形成乙酰肉堿和游離輔酶A,肉堿乙酰基轉移酶的表達可以成功地用于調節酒的風味[20]。
葡萄酒發酵前菌種活化以及葡萄除梗、破碎,或者為滿足酵母菌快速生長的需要,葡萄汁中自然溶入一定量的氧,這使得葡萄酒發酵初期部分脂肪酸可以發生β-氧化。脂肪酸β-氧化主要意義在于釋放能量和產生乙酰輔酶A。乙酰輔酶A不僅是初級代謝中間體,還在酯類物質的合成中起關鍵作用。
1.2 脂肪酸合成代謝
根據釀酒酵母脂肪酸組成,葡萄酒發酵過程中釀酒酵母生長繁殖需要相應量的C16:1、C18∶1和C16:0等脂肪酸,菌體所需脂肪酸來自自身合成和環境基質提供,當葡萄汁中外源性脂肪酸不能滿足生長繁殖需要時,酵母菌必須自身合成。釀酒酵母脂肪酸初始合成在細胞漿和線粒體中發生,由乙酰輔酶A羧化為丙二單酰輔酶A(圖2)。

圖2 釀酒酵母細胞漿中脂肪酸合成及中鏈脂肪酸釋放示意圖Figure 2 Fatty acid synthesis and medium chain fatty acid release in Saccharomyces cerevisiae cytoplasm
胞漿中的乙酰輔酶A在乙酰輔酶A羧化酶作用下,生成丙二單酰輔酶A。乙酰輔酶A、丙二單酰輔酶A分別在乙酰輔酶A-ACP轉酰基酶、丙二單酰輔酶A-ACP轉酰基酶作用下,轉酰基為乙酰-ACP、丙二酰-ACP(ACP為酰基載體蛋白),之后在脂肪酸合成酶多酶復合體作用下,經過“縮合-還原-水解-還原”循環,每完成一次循環碳鏈增加2個碳原子,碳鏈延長過程經丁酰-ACP、己酰-ACP、辛酰-ACP、癸酰-ACP、十二酰-ACP等,直至目的產物,終止于硫酯酶作用,并在轉酰基酶作用下轉脂酰基到輔酶A上,生成LCFA-CoA,進入磷酸甘油酯合成或脂肪合成[21]。在脂質合成過程中,乙酰輔酶A羧化酶是脂肪酸合成的限速酶,該酶的催化反應是脂質合成代謝中的一個關鍵調控環節,已知的激活劑有檸檬酸和異檸檬酸;長鏈乙酰輔酶A(如棕櫚酸)則是強反饋抑制劑,乙酰輔酶A的活化與抑制作用非常協調[22]。
酵母菌脂肪酸合成過程中,在乙酰輔酶A羧化酶活化啟動到被抑制之間,釋放出LCFA-CoA(圖2)。乙酰輔酶A羧化酶啟動脂肪酸合成,并在缺氧條件下被長鏈飽和酰基輔酶A抑制,導致脂肪酸合成酶復合物中釋放出MCFA-CoA;在有氧的情況下,長鏈飽和酰基輔酶A轉化為不飽和酰基輔酶A,不抑制乙酰輔酶A羧化酶,不引起脂肪酸合酶復合物中MCA-CoA的釋放[23]。所以說,發酵過程中排出的MCFA是通過脂肪酸合成代謝而不是通過分解代謝產生[24]。MCACoA作為LCFA合成代謝中間體,又作為MCFAEE合成前體物,在酰基轉移酶作用下生成MCFAEE。
2 葡萄酒發酵中鏈脂肪酸乙酯合成機制與調控策略分析
2.1 中鏈脂肪酸乙酯合成酶的特異性
根據MCFAEE合成前體物MCFA來自脂肪酸合成中間體。對具體一款葡萄酒來說,各MCFA產生的量基本相同,但前述葡萄酒、白蘭地香氣分析中辛酸乙酯相對含量出現高于其它酯類化合物的結果,可能是由酰基轉移酶特異性導致。Saerens等[25]研究顯示,釀酒酵母中Eht1p和Eeb1p具有合成MCFAEE的能力和酯酶活性,是形成MCFAEE最重要的酶,EHT1和EEB1雙重缺失使辛酸乙酯的生成減少70%,Eht1p偏愛辛酰輔酶A做底物。Knight等[26]從動力學角度研究了體外Eht1對己酰輔酶A、辛酰輔酶A、癸酰輔酶A、十二酰輔酶A的親和性和催化效率,結果顯示,Eht1p的米氏常數(Km,mol·L-1)值順序為:辛酰輔酶A<癸酰輔酶A<十二酰輔酶A<己酰輔酶A,對辛酰基親和力最大;Eht1的催化常數(Kcat,S-1)值順序為:辛酰輔酶A>癸酰輔酶A>己酰輔酶A>十二酰輔酶A,對辛酸酰基催化轉化最快;Eht1的Kcat/Km(L·mol-1S-1)值順序為:辛酰輔酶A>癸酰輔酶A>十二酰輔酶A>己酰輔酶A,對辛酰基催圖化效率最高。Lilly等[27]的研究進一步證實,Eht1p催化了MCFAEE的合成,用商業葡萄酒酵母菌株VIN13(pEHT1-s)發酵的佐餐葡萄酒中己酸乙酯、辛酸乙酯和癸酸乙酯的濃度顯著增加,感官分析中癸酸乙酯、己酸乙酯(蘋果香氣)和辛酸乙酯的影響也明顯增強。這些研究結果表明,Eht1p具有催化辛酸乙酯等MCFAEE合成的特異性。
Byrne等[28]研究發現,EHT1和EEB1密切相關,堿基序列有58%的同一性、73%的相似性,是基因組重復復制的旁系同源蛋白。釀酒酵母Eht1p和Eeb1p是在底物誘導下產生的,辛酸(C8)能夠誘導EEB1和EHT1的表達,且EEB1表達上調顯著[29]。熊國通等[30]研究了乙醇脅迫條件下EEB1相對表達,3%、6%、10%乙醇處理的基因表達量分別是對照組的22倍、407倍和5113倍,差異顯著。
目前,人們已發現并鑒定了釀酒酵母中5種與酯類化合物合成相關的醇酰基轉移酶,即Atf1p、atf2p、Eht1p、Eeb1p和與Atf1p同源的Lg-Atf1p;分別由ATF1、ATF2、EHT1、EEB1編碼;1種與分解有關的酯酶Iah1p,由IAH1編碼。Eht1p和Eeb1p在發酵中催化MCFAEE合成;Atf1p、atf2p催化乙酸乙酯和乙酸異戊酯生產,Lg-Atf1p參與乙酸乙酯和乙酸異戊酯的生產;Iah1p酶水解乙酸異戊酯、乙酸乙酯和乙酸-2-苯乙酯,并在很大程度上水解乙酸己酯[24]。
Nancolas等[31]在研究Atf1p生化性質時發現,Atf1p對一系列脂酰輔酶A具有活性,并且對C8(辛酰基)底物具有最大的催化效率。Eht1p與Atf1p兩種酶之間的主要區別在于Eht1p的底物范圍更窄、效率更高,對辛酰基輔酶A的偏愛性更強。Eht1p和Atf1p相對于辛酰基輔酶A的Kcat/Km分別為15×104和8×104L·mol-1·S-1,二者均具有硫酯酶活性。Eht1p兼有硫酯酶功能,即使在高乙醇濃度下也能水解MCA-CoA生成MCFA,與Saerens、Knight等[25-26]的研究結果一致。
2.2 中鏈脂肪酸乙酯合成機制
Eht1p是一種位于酵母脂質顆粒和線粒體外膜的脂蛋白[32-33],具有轉移酶(乙醇-O-酰基轉移酶)和水解酶(硫酯酶)功能,與酶蛋白質一級和三級結構及底物誘導蛋白構像發生變化有關。Knight等[26]多序列比對預測,在Eht1p C末端的α/β水解酶結構域具有絲氨酸、天冬氨酸、組氨酸殘基(Ser-Asp-His)催化三聯體,核心催化殘基Ser247、Asp395和His423的作用。釀酒酵母可產生多種硫酯酶,Cantu等[34]預測了釀酒酵母硫酯酶可催化殘基Ser161、Asp241和His276。
葡萄酒發酵過程中,釀酒細胞內MCFA-CoA與乙醇在酰基轉移酶Eht1p、Eeb1p作用下生成MCFAEE(圖3)。根據Knight等[26]和Saerens等[24]對Eht1p核心催化殘基預測結果,結合催化三聯體親核催化和廣義酸堿催化協調機制,做Eht1p催化合成MCFAEE機制圖(圖4)。
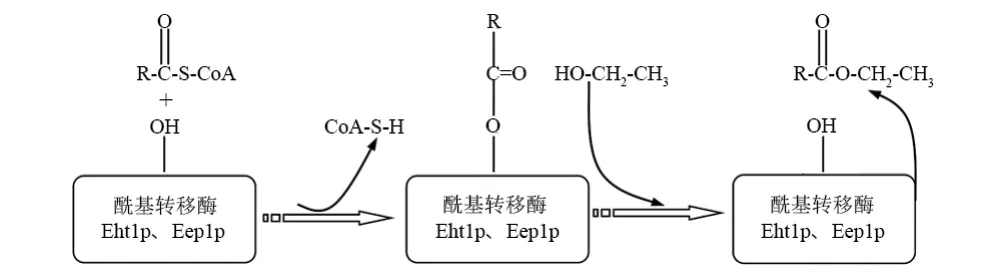
圖3 酰基轉移酶催化中鏈脂肪酸形成示意Figure 3 Acyltransferase catalyzes the formation of mediumchain fatty acids
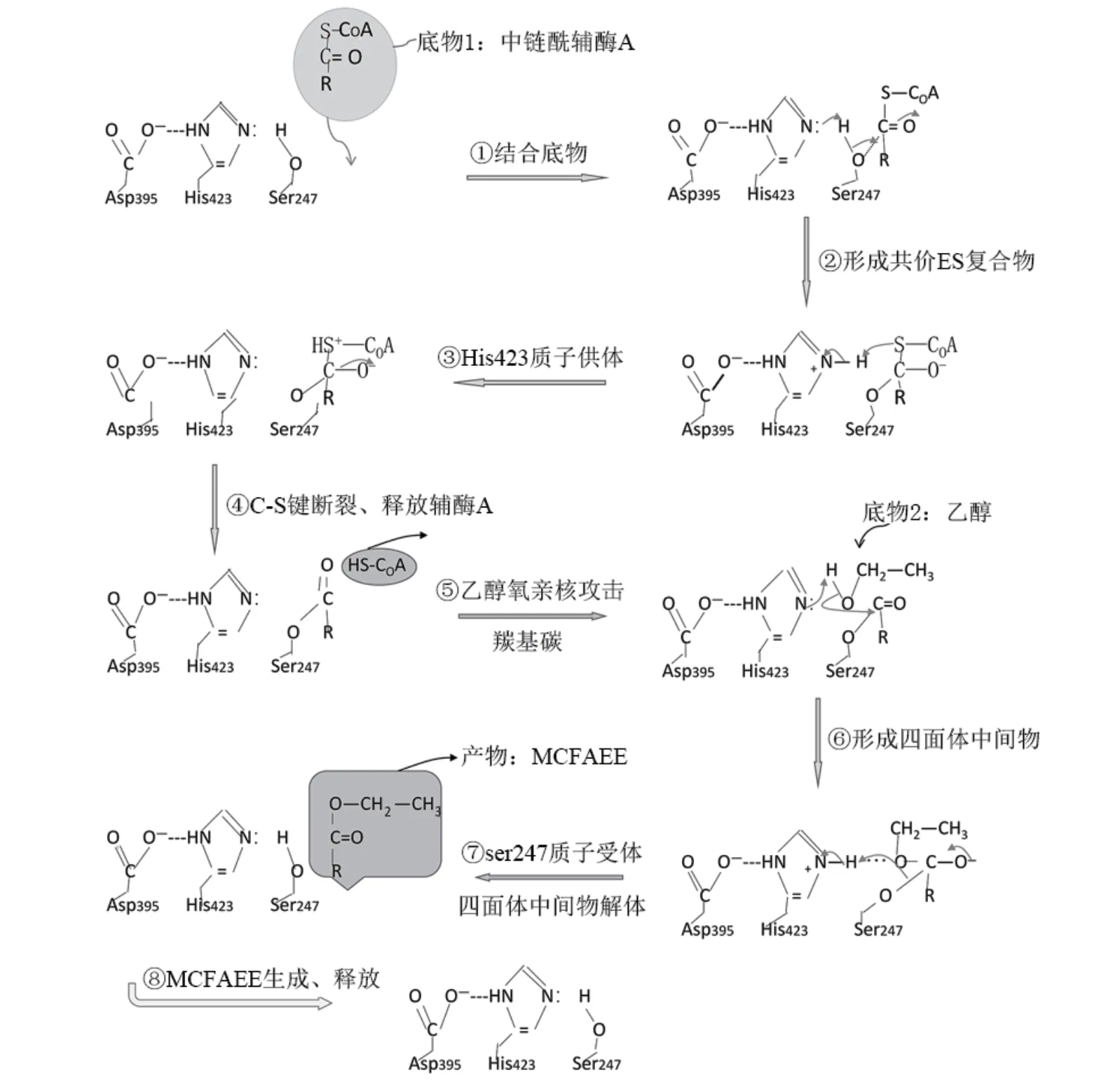
圖4 酰基轉移酶催化中鏈脂肪酸乙酯合成機制Figure 4 Acetyltransferase-catalyzed synthesis mechanism of medium-chain fatty acid ethyl esters
首先MCFA-CoA與酶活結合,誘導酶構象契合變化,在活性中心催化三聯體(Ser-Asp-His)絲氨酸殘基羥基質子轉移到組氨酸殘基咪唑基,帶負電荷氧原子攻擊底物酰基輔酶A羰基碳原子,形成底物與絲氨酸殘基共價過渡態,繼而組氨酸殘基咪唑基脫去質子,酰基輔酶A硫酯鍵斷裂,絲氨酸殘基羥基氧與底物羰基碳形成酯共價鍵,完成第一次親核反應;接著乙醇進入,在組氨酸殘基作用下脫去羥基質子,帶負電荷氧原子攻擊酯鍵羰基碳原子,再一次發生親核反應,將酰基轉移與乙醇中的氧結合形成MCFAEE,同時絲氨酸殘基獲得質子被還原。
2.3 葡萄酒發酵中鏈脂肪酸乙酯調控策略分析
葡萄酒中已鑒定出800多種香氣化合物[35-36],每款葡萄酒的香氣都是眾多香氣化合物整體組合的嗅覺表現,且每一香氣組分增量都會引起香氣整體嗅覺變化,特別是嗅覺閾值較低的化合物引起的整體變化更為顯著。葡萄酒發酵酵母菌代謝產生的酯類化合物乙酸酯類(乙酸乙酯、乙酸異戊酯、乙酸苯乙酯、乙酸己酯等)和乙酯類(乙酸乙酯、丁酸乙酯、己酸乙酯、辛酸乙酯、癸酸乙酯、十二酸乙酯等)對葡萄酒香氣具有重要貢獻,MCFAEE是白蘭地酒特征香氣成分,所以MCFAEE的代謝積累是葡萄酒、白蘭地釀造關注的重點。
基于MCFAEE源于脂肪酸合成代謝,并且催化MCFAEE合成的酰基轉移酶Eht1p、Eeb1p受底物MCFA-CoA誘導,以及EHT1或EEB1過表達不明顯影響乙酯的生產[25,27],即:MCFAEE的產量取決于底物的濃度,酵母細胞內MCFA-CoA濃度是MCFAEE產量的限制因素[24]。所以,MCFAEE發酵調控對象是脂肪酸合成,調控目標是MCFA-CoA釋放量。脂肪酸合成過程中,乙酰輔酶A是基礎原料,它的生成量、流向脂肪酸合成方向的量決定著脂肪酸合成水平。乙酰輔酶A的生成量與酵母菌生物量呈正相關,厭氧條件下乙酰輔酶A向脂肪酸合成的流量與細胞生命力(時序壽命、比生長率)相關。所以,通過調控葡萄酒發酵過程中釀酒酵母生物量、比生長率和時序壽命,可以達到調控脂肪酸合成水平的目的。
發酵溫度影響釀酒酵母生長速率、生物量,能夠調整釀酒酵母次生代謝產物通量,強烈影響脂質代謝。Beltran等[37]分析了13 ℃和25 ℃發酵條件下釀酒酵母顯著差異表達的535個基因,發現在25 ℃下的發酵中,與胞質脂肪酸合成有關的幾個基因下調;在13 ℃下的發酵中,與線粒體短鏈脂肪酸合成有關的基因上調。Schwinn等[38]研究了‘雷司令’發酵過程中溫度對酵母生長、揮發物等的影響,結果顯示,在19 ℃下發酵的后半段,除丁酸乙酯外,所有乙酯的最大濃度均高于14 ℃的發酵處理;在19 ℃下葡萄酒中酵母總數也最高,為1.5×108cfu·mL-1,而在14 ℃和16-11-17 ℃梯度發酵的葡萄酒中,酵母總數分別為1.3×108、1.2×108cfu·mL-1,差異約為20%。所以,溫度因素通過影響脂肪酸合成途徑相關基因的表達來影響MCFAEE產量。
氮是構成蛋白質和核酸的元素,氮素是常見的微生物生長限制因素。對于確定的釀酒酵母菌株,葡萄汁可同化氮影響葡萄酒發酵生物量、發酵速率和發酵產物。劉沛通等[39]比較了不同菌株150、250 mg·L-1氮濃度下發酵動力學和產物揮發性成分,結果顯示,氮的利用具有菌株特異性,但都表現出低濃度可同化氮的甘油產量高的特點,不同菌株表現出不同的酯和脂肪酸生成模式,商業菌株酯產量與初始可同化氮濃度正相關。
在葡萄酒發酵過程中,不飽和脂肪酸(UFAs)是釀酒酵母生長繁殖的必需營養物質,釀酒酵母主要從葡萄汁中獲得UFAs[39],因為UFAs不能在缺氧的條件下合成。Beltran等[40]研究了發酵培養基脂質組成對酒中揮發性成分的影響,發現葡萄汁與合成葡萄汁相比,前者比后者完成發酵快,分別為14、25 d;由葡萄汁獲得的葡萄酒中揮發性化合物的濃度較高,辛酸乙酯的含量是合成葡萄汁發酵的兩倍。這與合成葡萄汁中缺乏UFAs等生長因子,厭氧條件下限制了酵母菌生長有關。
氧氣是葡萄酒發酵中不可忽視的重要因素,葡萄酒發酵并非絕對厭氧發酵,酵母菌脂肪酸代謝是在有氧條件下進行的,微量氧氣對于酵母菌緩慢生長和提高次生代謝產物產量具有重要作用。有研究比較了釀酒酵母需氧培養和厭氧培養,結果顯示有500個基因表達存在顯著差異,只有23個基因是厭氧菌生長所特有,對于這些基因在無氧條件下表達量的差異性或其關鍵作用機制還需要進一步探索[41]。
綜上所述,葡萄酒發酵過程中釀酒酵母生物量、細胞生命活力(比生長率、時序壽命)狀態參量決定著MCFAEE代謝產量,環境變量溫度、發酵基質和氧氣等,顯著影響細胞總量、細胞生命活力。葡萄酒發酵采用適度低溫、適當溶氧和控制葡萄汁脂質、適量可同化氮,提高細胞群體峰高和細胞時序壽命,降低細胞比生長率,可以達到提高MCFAEE產量目的。這符合一般發酵規律,次級代謝作用只有當菌體在低的比生長率條件下才可發生[22]。
3 總結與展望
本文以釀酒酵母脂肪酸代謝、中鏈脂肪酸乙酯合成酶系的研究進展為基礎,探討了葡萄酒發酵中鏈脂肪酸乙酯的生成機理及調控策略,理清了基于釀酒酵母脂肪酸合成代謝的中鏈脂肪酸乙酯合成途徑,討論了脂肪酸合成限速酶(乙酰輔酶A羧化酶)、脂肪酸合成酶復合體、硫酯酶、硫激酶在中鏈酯酰輔酶A形成中的作用。并指出中鏈酯酰輔酶A為中鏈脂肪酸合成的限制因子,重點分析了酰基轉移酶Eht1p和Eeb1p的特異性及催化機制,結合實踐,圍繞提高葡萄酒發酵中鏈脂肪酸乙酯產量,提出了以發酵環境變量(溫度、溶氧、葡萄汁脂質、可同化氮)為手段,以釀酒酵母細胞總數、細胞生命活力為靶標的調控策略,為優化葡萄酒、白蘭地酒香氣質量提供了參考。
酒類風味是風味科學的重要領域,香氣在飲料酒風味中占主要地位,中鏈脂肪酸乙酯是飲料酒產品果香香韻特征香氣成分,研究其合成機制與調控策略,不但是釀酒風味科學向縱深發展的選項,而且對于促進飲料酒產品品質升級和飲料酒產業高質量發展具有重要意義。
中鏈脂肪酸及其衍生物在能源、醫藥領域有著廣闊的應用前景。基于釀酒酵母模式生物探索中鏈脂肪酸生物合成途徑與機制,進而利用合成生物學手段創新生物資源,從頭合成中鏈脂肪酸及其衍生物,符合經濟建設、人類健康需求。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
