根系分泌物主要作用及解析技術進展①
2023-05-26 02:51:52唐昊冶郭家歡王如海吳永紅俞元春
土壤 2023年2期
關鍵詞:植物
陳 虹,唐昊冶,郭家歡,潘 暢,王如海,吳永紅,俞元春*
根系分泌物主要作用及解析技術進展①
陳 虹1,2,唐昊冶2,郭家歡1,潘 暢1,王如海2,吳永紅2,俞元春1*
(1 南京林業大學南方現代林業協同創新中心,生物與環境學院,南京 210037;2 中國科學院南京土壤研究所土壤與環境分析測試中心,南京 210008)
根系分泌物是植物保持根際微生態系統活力的關鍵因素,也是根際物質循環的重要組成部分,對根際土壤生態環境中的物質循環具有重要的驅動作用。根系分泌物可以刺激微生物生長,增強其活性,加速根際養分循環,增加土壤養分利用率,并在小規模空間引起溫室氣體通量的變化。此外,它也是植物參與競爭的重要策略,植物通過根分泌物以獲取種間長期生存的養分,甚至分泌對自身有害的化感物質來排擠其他植物,實現自我生存,即使存在自毒作用或引起連作障礙等。植物的健康生長依賴于自身與土壤微生物復雜動態群落的相互作用,但是根際微生物群落結構和組成卻又受植物物種、植物生長期、土壤性質、功能基因等因素影響,這些因素的動態變化可能導致根系分泌物的多樣化,從而形成復雜多變的根系分泌物與植物的關系,進而影響植物的健康生長。目前,對植物根系分泌物的研究是土壤生態學、植物營養與代謝等領域的研究熱點,且隨著分析技術手段的快速發展,根系分泌物相關研究也逐漸深入,進一步揭示植物與微生物間的協同作用機理對農、林等行業生產具有重要的指導意義。
根系分泌物;分析技術;土壤;微生物
根系分泌物是植物生長過程中根系不同部位向生長介質中分泌或釋放的種類繁多的碳活性物質,包括相對低分子質量的有機物質、高分子質量的黏膠物質、根細胞脫落物及其分解產物以及氣體、質子和養分離子等[1]。迄今為止,已鑒定的根系分泌物有300多種,主要包括碳水化合物、氨基酸、有機酸、酶和其他化合物等,詳見表1[2–9]。根系分泌物不僅數量可觀,而且作用巨大,是保持根際微生態系統活力的關鍵因素,也是根際物質循環的重要組成部分。但是不透明的復雜土壤環境給分泌物研究帶來了重重困難,研究人員仍需進一步研究根系分泌物對農、林生態系統乃至全球生態系統的影響,這就強烈依賴于分析技術的不斷發展以提供更深入、便捷、準確的分析手段和途徑,來揭示根系分泌物對根際土壤生態環境的調控機制,為相關行業生產管控提供科學的理論支撐。
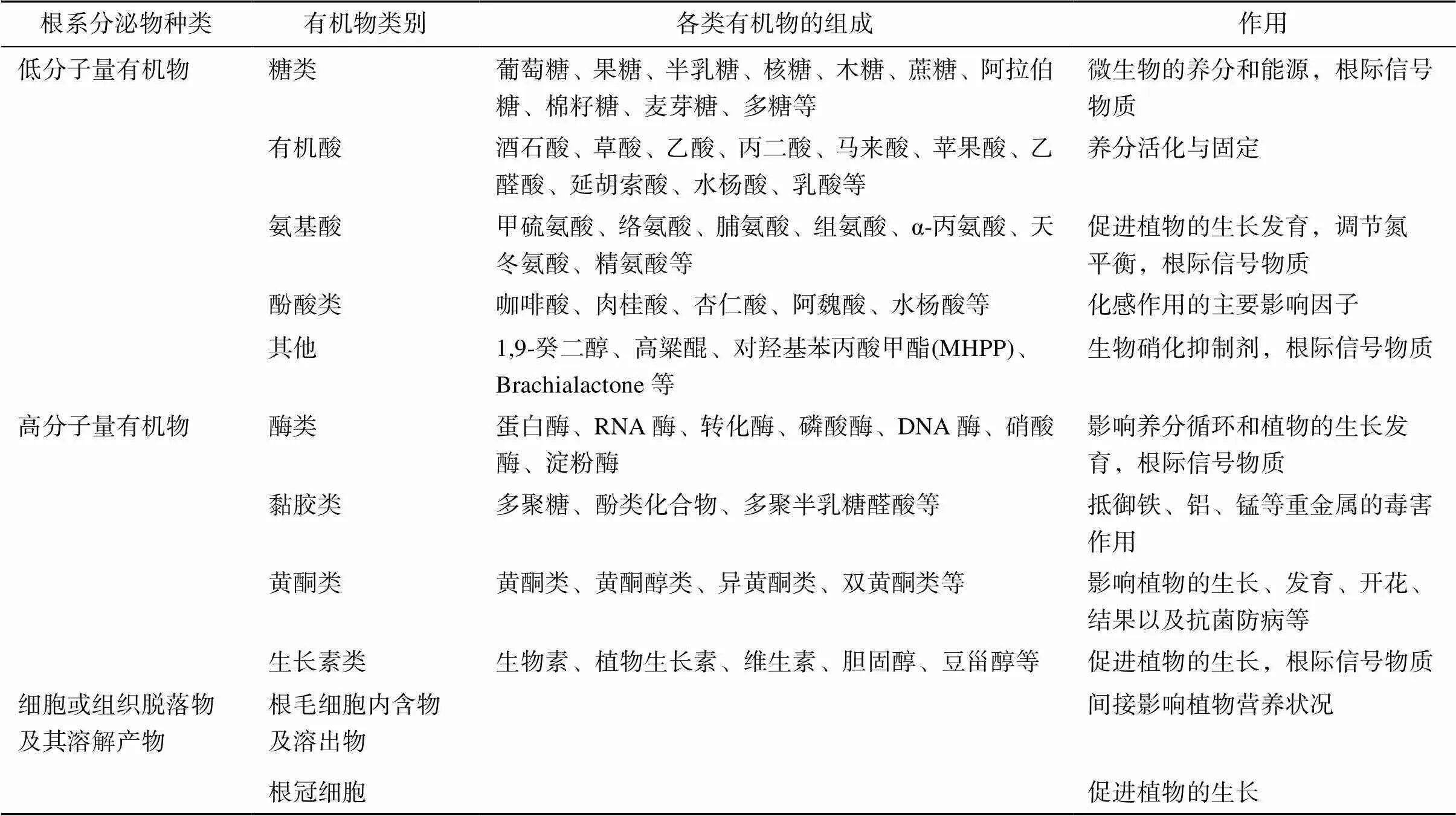
表1 根系分泌物組分及其作用
1 根系分泌物對土壤生態環境中物質循環的驅動作用
1.1 驅動土壤有機質轉化
土壤有機質(SOM)是指土壤中的有機碳化合物,來源于植物的凋落物和死根、死的土壤生物、土壤生物和植物的排泄物或分泌物、人為有機肥和廢物等。SOM對維持農業、林業等生態系統生產力具有重要意義[10],其中SOM礦化被認為是調節全球碳循環的關鍵過程[11],也是植物生長過程中主要養分來源。但植物只有在生物或非生物降解作用下才能直接吸收來自SOM的礦化營養物質。Nardi等[12]發現農林植物根系分泌物中琥珀酸具有較高的類生長素活性,脂肪酸C14COOH、C15COOH和C17COOH的同分異構體活性最高,結合之前Piccolo等[13–14]的研究,他們認為有機酸可進入腐殖質膠束內部并改變其立體疏水結構,生物分子是由根系分泌物中的有機酸從大部分土壤中驅動起來的,植物可以介導根際的發育和生理活動,并進一步驗證了根系分泌物對SOM的生物活性、打破腐殖質交聯和釋放激素類活性物質的重要作用。Nardi等[15]還通過有機酸提取物對玉米苗代謝特別是對氮同化的影響測試,發現農田土壤提取物比森林土壤提取物表現出更高的類激素活性,應是刺激了氮通路所致。Wang等[16]將3種分泌物模擬物(即葡萄糖、甘氨酸和草酸)添加到200年歷史的云杉、冷杉林土壤中,證實不同組分的根系分泌物通過不同的生物和非生物啟動機制深刻調節了土壤碳動態和土壤碳儲量。
1.2 提高土壤生態系統中養分利用率
SOM作為微生物底物在化學上難以降解或在能量上無法激活[17],但植物可以通過釋放刺激微生物生長、活動和周轉的根系分泌物來加速根際氮循環[18],如分泌物刺激微生物生長,誘導微生物釋放胞外酶進而促進氮循環,并通過啟動效應解聚SOM[19]。森林土壤中SOM含量遠高于農業土壤,在森林土壤中酚和蛋白復合物可驅動沉積在慢循環SOM庫中的可激活有機氮[20–21]。因此,植物通過釋放根系分泌物,為微生物提供能量,促使其產生胞外酶,從SOM中掘取氮[22–23]。如果大部分的氮可由緩慢循環的SOM啟動,那么森林生態系統就會有幾乎取之不盡的氮供應,而不再受氮的限制。Meier等[24]發現,在紅松人工林根際原位添加模擬分泌物增強了快速降解、循環氮庫酶的特定活性,增加了微生物對氮降解化合物的分配;并推測根系分泌物能刺激微生物分解不穩定的SOM并釋放氮,若在富氮土壤中植物能激發這種功能,其作用可能更大。經Yin等[25]推算,在兩種外生菌根和兩種叢枝菌根樹種的森林中,根系每年分泌物質量占凈初級生產力(NPP)總量的2.5%,而該分泌量引起的微生物氮循環變化可能占總凈氮礦化量的18% 左右。而陸玉芳和施衛明[6]及Lu等[26]確認根系還會分泌一些特定的化合物——生物硝化抑制劑,其能夠抑制硝化作用,提高根際氮循環中氮的利用率。Strickland等[27]發現在森林土壤有機碳和微生物碳中,分泌物葡萄糖(13C標記)迅速升高,表明根系分泌物可以刺激微生物和動物群的活動,以獲取營養。在缺氮條件下,玉米通過主動調節根系分泌物的組成,增加與根際細菌的相互作用,以刺激水解酶的產生和活性,并改變它們的根際范圍,在更大的土壤體積中調動氮和碳[28]。而根系分泌物中有機酸的分泌使得低肥力土壤中氮的礦化增加[29]。
磷(P)是植物生長最重要的營養物質之一,也是植物生長過程中最重要的限制養分之一。許多農業系統強烈地依賴于肥料磷的連續輸入;與此相反,在森林生態系統中,生物群具有高效驅動、吸收、回收和再利用磷的途徑[30]。因此,在森林土壤中,應充分研究植物從有機磷和礦物磷中獲取可用磷的策略,即無機磷增溶和有機磷礦化。33P同位素稀釋法被用于研究根系分泌物對微生物總有機磷礦化的促進作用,結果發現,添加丙氨酸和葡萄糖可以刺激微生物的總磷礦化和磷酸酶活性,根系分泌物的結構和化學計量比可以顯著影響微生物的刺激程度,因此根系分泌物對非菌根微生物的刺激可能是植物提高土壤中磷有效性的重要途徑[31]。Zhang等[32]研究發現,根系共生的叢枝菌根真菌分泌的果糖不僅是一種碳源,而且可作為信號分子觸發細菌介導的有機磷礦化過程,可使根系獲得可靠的磷。白羽扇豆() 等植物在低磷脅迫下會形成大量排根,排根能分泌檸檬酸等有機酸來活化土壤中的難溶性磷[33]。青藏高原東部亞高山森林根際土壤速效磷含量明顯高于非根際,并與檸檬酸含量高度相關,可能是檸檬酸通過解吸或配體交換有效調節了土壤中的磷[34]。
1.3 改變溫室氣體排放通量
根際土壤環境中活性炭、氮化合物通常來自根系分泌物,一般是低分子量的碳化合物,包括糖、有機酸和氨基酸等,可為大部分微生物的生長和活動提供能量,并占土壤呼吸總量的30%[35–36]。Strickland等[27]使用低分子量的葡萄糖(13C標記)追蹤位于美國南卡羅來納州3個牧場和3個森林土壤的主要碳軌跡,72 h后,葡萄糖中13C在微生物生物量碳和土壤有機碳中所占比例最大,其次是CO2、溶解性有機碳和微節肢動物;進入地下系統的低分子量碳化合物可能通過土壤食物鏈迅速傳播。因此,豐富的根系分泌物可以誘導更多的CO2排放。
濕地(包括泥炭地)在全球碳循環中發揮著至關重要的作用,其CH4(甲烷)排放量占全球甲烷排放總量的20% ~ 39%;泥炭地作為巨大的碳儲存地,是植物和土壤呼吸產生CO2的重要來源,也是缺氧條件下有機物降解產生CH4的重要來源[37]。將根模擬分泌物(糖和有機酸組成的混合物)加入泥炭中以評估CO2和CH4的排放,結果顯示,CO2和CH4排放量增加,其響應的幅度和持續時間取決于添加的不穩定碳混合物的成分而不是添加的碳的量或泥炭的源植物。所以,同樣證實了根系分泌物成分在調節溫室氣體通量中的重要性;且原位植物物種的變化,特別是那些與土地利用變化相關的變化,可能解釋了由于物種特有的根系分泌物組成而帶來的CO2和CH4通量的小規模空間變化[38]。
在農田生態系統中,不同C/N比的模擬根系分泌物添加對水稻土CO2的排放均有影響,增加分泌物C/N比會加劇CO2排放[39]。同樣,模擬根系分泌物(草酸、絲氨酸和葡萄糖)添加到培養的農田土壤中加劇了氧化亞氮(N2O)的排放,高濃度葡萄糖處理下N2O排放量最高,低濃度草酸處理下N2O排放量最高,且根系分泌物濃度越高,即土壤所提供的碳源越多,越促進異養微生物作用下的反硝化過程,對N2O的貢獻越大[40]。而根系分泌的1,9-癸二醇等生物硝化抑制劑還能通過抑制硝化作用而降低土壤N2O的排放[26]。
2 根系分泌物對植物種間競爭與共生的影響
根系分泌物的化學成分因物種和植物發育階段以及當時的環境條件而異,但應包括不穩定的糖、有機酸和氨基酸等。因此,植物可根據食物——糖的分類來選擇微生物群落和動物群,以達到植物與生物之間和諧共生,相互促進[41–42]。但是與植物的其他碳輸入(如凋落物等)相比,根系分泌物的碳濃度較低,微生物對其利用卻很快。不同的植物根系分泌物(可能是有機酸)可能是植物物種間長期共生的養分獲取途徑。
但有些植物會分泌有害的有機化合物(化感物質),這些化合物可以殺死對其敏感的植物,將其從小環境中排除,這種排他作用也可能通過影響土壤微生物群落來實現[43–44]。Fernandez等[45]用櫟樹或Molinia草的根系分泌物來澆灌橡樹苗,發現Molinia草根系分泌物顯著降低了橡樹的生物量增長,櫟根分泌物則有利于橡樹生長,因此種植橡樹時應減少橡樹幼苗附近的Molinia草,以幫助減少其化感作用和緩解資源競爭。Xia等[46]將闊葉樹種與杉木混交種植,發現與香樟混交后,香樟根系分泌的化感物質抑制杉木的生長,且土壤主要微生物類群發生變化;而與櫟樹混交后,杉木生長得以持續促進,主要表現在其促進杉木根的生長和改變根的行為,這為杉木人工林的管控提供了重要的啟示。而根據Vyvyan[47]對作物分泌物的研究,一些有害物質或化感物質甚至可以被認為是新型除草劑或農藥,植物可通過根系分泌、葉片揮發、凋落物降解淋濾等方式,釋放對其他植物有害的物質到周圍的環境中,以減少其他植物競爭生長的機會。
3 根系分泌物對土壤微生物的影響
植物在自然環境中的健康依賴于由宏觀微生物和微生物組成的復雜動態群落的相互作用,其生長量和產量取決于大量的與細菌和真菌的相互作用[48–49]。但是根際微生物群落結構和組成又受植物物種、土壤性質等影響,且在植物的不同發育階段具有不同的根系分泌物和根際效應[50–51],進而微生物群落也會隨著植物的年齡而變化,甚至會受到特定根系分泌物的調控而被選擇性修飾。Liu等[52]研究發現,刺槐造林后根際土壤和細根特性高度誘導了主要細菌菌屬和真菌菌屬的變異量,土壤可利用磷和細根總磷是調節根際土壤微生物群落的關鍵,根際微生物群落組成受細根和土壤性質的協同影響。肉桂酸是一種抑制土壤微生物、促進根系生長的化感物質,Mehmood等[53]等證實肉桂酸降低了玉米根系生長和次生代謝產物的分泌,還可能影響其吸引促進植物生長的內生真菌的能力。Yuan等[54]發現,香蕉根分泌物中的酚酸可能增強了淀粉芽孢桿菌njk-6 (一種高效的拮抗真菌)的根定植和抑菌能力。Shi等[55]發現,一年生草本植物始終抑制根凋落物的分解速率,通過建立概念模型確認了兩種主要機制:一是細菌和真菌的根際種群在遺傳上可能比不與根系相關的種群更能利用小分子量分泌物,而更不可能使用復雜的大分子;二是植物蒸散和土壤干燥速率高,可能會降低微生物的整體功能潛力,從而抑制分解活動。該機制解釋了植物誘導的分解主因的大小(甚至方向)是如何由根系對微生物功能基因的影響驅動的。同樣,Huang等[56]證實,擬南芥根三萜類代謝物主導了根內、根際微生物組的建成與維持,塑造了根際的微生物群落,以滿足其生長需求。Wen等[57]推測,易感品種的黃瓜傾向于通過根系分泌更多的有機酸(檸檬酸、丙酮酸、琥珀酸和富馬酸)來組裝有益微生物。
根分泌物在豆科植物與根瘤菌的共生關系中也起著重要作用。Sasse等[58]認為,植物遺傳因素特別是根的形態和根系分泌物是形成根瘤菌的主因,宿主基因型、馴化和植物發育明顯影響根瘤菌的組成。Wang等[59]證實,真菌內生菌接種至花生,改變了花生的根分泌物酚酸、黃酮類化合物、有機酸和氨基酸的濃度,并誘導了根瘤菌的趨化行為;其中有機酸(檸檬酸和草酸)和氨基酸(谷氨酸和氨基酸)起了誘導作用,甘氨酸和谷氨酰胺在較高濃度下可促進根瘤菌生長和生物膜形成:根分泌物的改變有助于介導的花生根瘤菌結瘤增強。
此外,根系代謝產物和根際微生物群落的共生結合也是植物克服重金屬脅迫、養分脅迫的關鍵適應機制,即拮抗作用以減輕環境脅迫帶來的生長不良影響[3–4,60];或通過改變根際土壤中細菌、真菌的種群數量及其群落結構所營造的根際微生態環境,以促進有機氯降解而緩解外源毒性[61]。
4 根系分泌物對土壤酶活性的影響
土壤酶是由生物體(包括土壤微生物、土壤動物、部分植物)產生的、具有高度催化作用的一類蛋白質,參與了土壤的發生、發育以及土壤肥力的形成和演化的全過程。不同酶活性可以反映微生物對土壤中不同底物的分解強度[62],常被作為表征SOM 中碳、氮分解的敏感指標[63]。
由于大量輸入易降解的低分子量有機物,即根系分泌物,微生物在根際更活躍,產生更多的酶[64]。在大多數植物中,低分子量有機物的主要形式是糖、氨基酸和羧酸[65],所以根際微生物活性取決于根系分泌物的組成,特別是糖、羧酸和氨基酸的比例,其同樣也影響土壤酶的活性。Zhang等[66]采用特殊的方法模擬根系分泌物釋放對分別含有碳、氮、磷、硫元素的5種微生物酶的影響,即含碳的β-葡糖苷酶和纖維二糖水解酶、含氮的亮氨酸氨基肽酶、含磷的磷酸酶和含硫的硫酸酯酶,發現含磷、含氮和含硫相關酶活性在人工根附近較高,隨著離根距離的增加而逐漸降低;碳循環酶的形態是均勻的,與根系分泌物的組成無關;磷酸酶是對根系分泌物成分最敏感的酶,任何影響根系分泌物成分的因素都會對磷循環產生強烈的影響。司鵬等[67]同樣證實小分子有機物(葡萄糖、甘油和甘氨酸)對土壤酶活性和土壤微生物的影響,發現葡萄糖對土壤過氧化氫酶活性影響顯著高于對照、甘油和甘氨酸處理,而對堿性磷酸酶活性影響顯著低于其他處理;且不同種類的小分子有機物對土壤酶活性的影響各不相同。張亮等[68]研究發現,半夏塊莖提取液顯著降低細菌和放線菌數量,顯著降低土壤微生物群落的多樣性、均勻性和優勢度指數,而對土壤酶活性的影響表現出多樣性,如提高蔗糖酶活性,降低脲酶和磷酸酶活性等。因此,根系分泌物可能改變土壤微生物種群結構,改變土壤酶活性,其中的化感物質甚至會造成連作障礙等。
Wang等[69]發現,不同林分密度下杉木人工林中土壤因子的變化可導致表層(0 ~ 20 cm)土壤酶活性及真菌群落豐富度和多樣性的變化,而微生物和胞外酶對土壤有機碳隨林齡變化較為敏感,5月和9月土壤有機碳組分、酶活性和微生物活性均高于1月,應是根系分泌物在土壤微環境中起了關鍵的調節作用。王小平等[70]通過原位收集連香樹根系分泌物的方式,同樣證實連香樹根系分泌物碳和氮分泌速率夏季最高,冬季最低,根際土壤微生物碳、氮及土壤酶活性顯著高于非根際,根系分泌物輸入是驅動根際微生物活性的重要因子。與純樟子松林相比,樟子松楊樹混交林分下樟子松根系分泌物使土壤有機碳、氮磷鉀、土壤酶活性和土壤微生物數量均得以顯著提升[71]。
5 根系分泌物解析技術進展
根系分泌物取樣多采用實驗室砂培、水培、土培植物收集[2,72–73],因這3種方式容易實現,故應用廣泛。原位收集[25,73]通常是將原位挖出的部分根系放入合適的容器中進行水培而獲得,因野外實施難度較大,對根系要求較高,故應用較少。但原位收集更能體現根系分泌物的實際代謝過程,所以多用于較大型木本植物的根系分泌物收集。
根系分泌物中發現的化合物已證實了植物與環境之間的相互作用[74]。但根系分泌物解析過程復雜,挑戰性極強,極大程度依賴于分析技術的發展。早期,對根系分泌物中總糖、有機酸等的分析主要依賴于分光光度法和滴定法,代謝物碳、氮等的分析則依賴于C/N分析儀、流動分析儀等,表征總量信息[27,72]。目前,隨著色譜、質譜等儀器的快速發展,收集到的分泌物經合適的方式進行富集或濃縮等預處理,可由GC (氣相色譜儀)、GC-MS (氣相色譜–質譜聯用儀)[72–73]、HPLC (液相色譜儀)[9,73]、IC (離子色譜儀)[65]、CE (毛細管電泳儀)[5]、LC-MS (液相色譜–質譜儀)[26,75]、NMR (核磁共振波譜儀)、FT-ICR MS (傅里葉變換–離子回旋共振質譜儀)等進行分析,由此可獲得準確的多種代謝物信息。隨著空間代謝組學研究的發展,近幾年質譜成像系統已用于植物中代謝物空間分析,如對根進行分析,可發現根橫切面上特異性代謝物的分泌位置;對莖、葉等不同部位進行質譜成像分析,可發現特異性代謝物的主要富集部位,結合LC-MS、FT-ICR MS等儀器解析結果,可為植物中代謝物合成、運輸和積累提供信息[76–77]。以上結果與土壤特性、微生物類群分析等綜合考量后,可揭示根系分泌物中特征物質在根際土壤生態環境中的調控作用,或與其他生物間的相互影響,為實際生產提供理論與實踐性支撐。圖1為根系分泌物解析技術參考圖。
以杉木的根系分泌物研究為例,學者們采用GC、GC-MS、HPLC、LC-MS等對杉木的根系分泌物、林下土壤或枝體提取物進行了分析,確認了環境或營養脅迫下根系分泌物組成特征[78–80]。熊德成等[81]在LC-TOFMS (液相色譜–飛行時間質譜聯用儀)幫助下具體分析了杉木的根系分泌物成分,發現增溫后杉木細根分泌物的抗逆境物質明顯增加,如用于抵御干旱環境的脫落酸、氨基酸類物質(胱氨酸)及酚酸類物質(山奈酚);用于活化土壤礦質養分的相關物質分泌也顯著增加,如檸檬酸、肉桂酸;有利于植物對抗外界傷害和病原菌侵染的一些物質也會增加,如茉莉酸、水楊酸及黃酮類物質(槲皮黃酮):證實植物在遭受根際高溫脅迫時會分泌一些特定的物質以有利于抵御環境的變化。環二肽是學者采用2D-NMR (二維–核磁共振波譜儀)和質譜技術在杉木凋落物、根系分泌物和連作土壤中發現的一種化感物質,并經證實其影響了相應的微生物群落組成,積累了潛在致病性真菌,阻礙了杉木根系的生長;而從杉木林下土壤中檢測到的酚酸類物質在自然濃度下并沒有抑制杉木幼苗的生長,反而有促進作用[82–83]。這些都為杉木的連栽障礙管控提供了寶貴的理論依據。
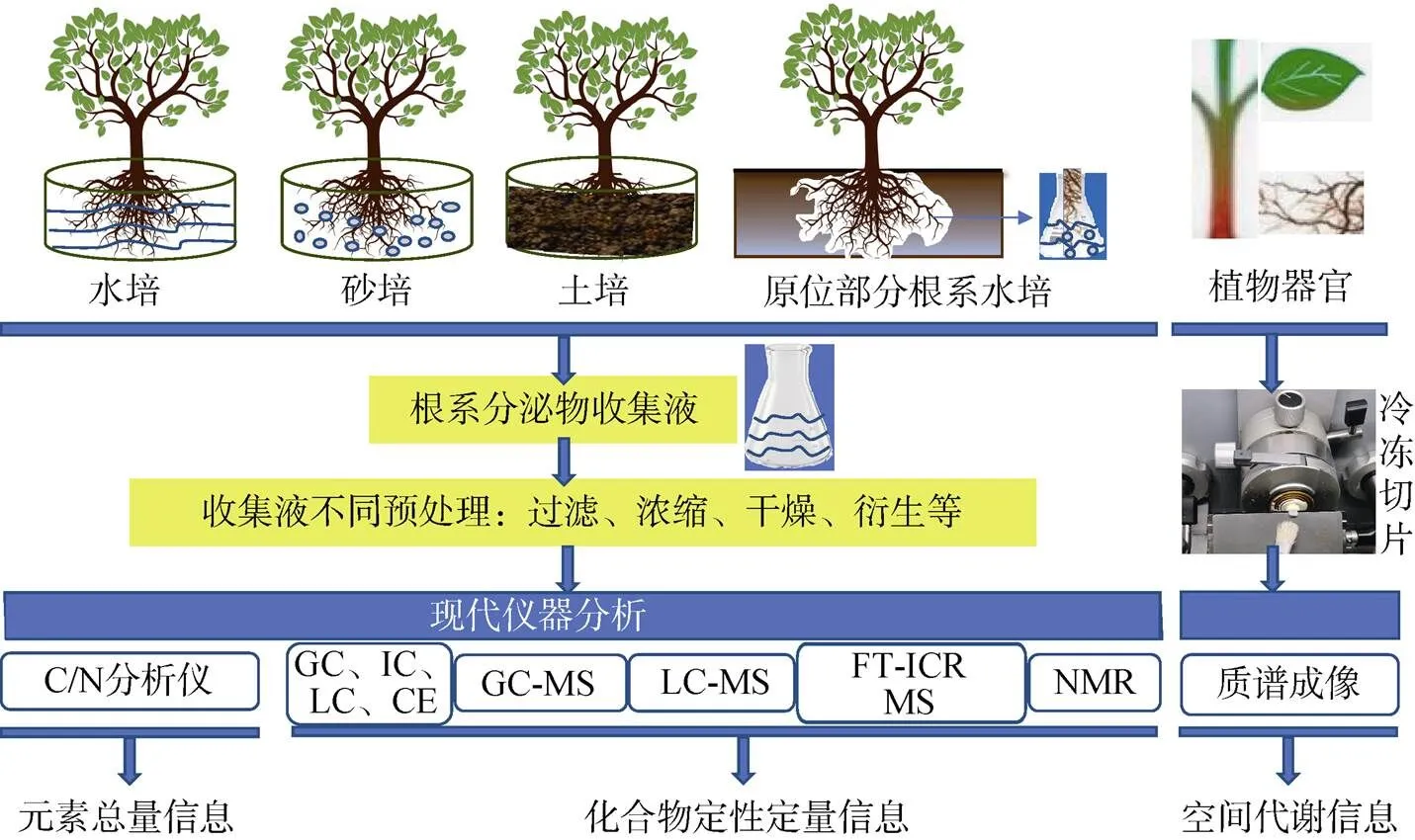
圖1 根系分泌物解析技術參考圖
6 研究展望
在全球范圍內,環境因素對微生物多樣性的影響大于植物多樣性,但在局域內,植物多樣性越高,凋落物和根系分泌物的差異越大,土壤微生物群落多樣性就越高,植物代謝產物是影響微生物群落的重要因素[84]。因此,在土壤這個“黑箱子”中,急需分析和識別多種根系化學信號物質[4],識別地下生物相互作用中最重要的化學特征,在此特征基礎上,探究代謝物對植物根際土壤生物學性質的影響機制。而實現這些目標的根本在于對根系分泌物的精準解析,更依賴于分泌物解析技術的發展,筆者認為主要需從3個方面切入,一是尋找最能代表植物根系真實代謝過程的根系分泌物收集方法,且須涵蓋草本類植物;二是確認可通用的精準的代謝物聯合分析技術;三是揭示根系信號物質[4]在土壤生態環境中對種間、生物間的相互影響與調控途徑,以為相關行業生產管控提供技術支撐。
[1] 周健民, 沈仁芳. 土壤學大辭典[Z]. 北京: 科學出版社, 2013.
[2] 劉芷宇, 李良謨, 施衛明. 根際研究法[M]. 南京: 江蘇科學技術出版社, 1997.
[3] 徐煒杰, 郭佳, 趙敏, 等. 重金屬污染土壤植物根系分泌物研究進展[J]. 浙江農林大學學報, 2017, 34(6): 1137–1148.
[4] 趙寬, 周葆華, 馬萬征, 等. 不同環境脅迫對根系分泌有機酸的影響研究進展[J]. 土壤, 2016, 48(2): 235–240.
[5] 陸玉芳, 施衛明. 根際化學信號物質與土壤養分轉化[J]. 生物技術通報, 2020, 36(9): 14–24.
[6] 陸玉芳, 施衛明. 生物硝化抑制劑的研究進展及其農業應用前景[J]. 土壤學報, 2021, 58(3): 545–557.
[7] Subbarao G V, Nakahara K, Ishikawa T, et al. Biological nitrification inhibition (BNI) activity in sorghum and its characterization[J]. Plant and Soil, 2013, 366(1): 243–259.
[8] Zakir H A K M, Subbarao G V, Pearse S J, et al. Detection, isolation and characterization of a root-exuded compound, methyl 3-(4-hydroxyphenyl) propionate, responsible for biological nitrification inhibition by()[J]. The New Phytologist, 2008, 180(2): 442–451.
[9] Subbarao G V, Nakahara K, Hurtado M P, et al. Evidence for biological nitrification inhibition inpastures[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(41): 17302–17307.
[10] Zhao S C, Li K J, Zhou W, et al. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China[J]. Agriculture, Ecosystems & Environment, 2016, 216: 82–88.
[11] Bellamy P H, Loveland P J, Bradley R I, et al. Carbon losses from all soils across England and Wales 1978–2003[J]. Nature, 2005, 437(7056): 245–248.
[12] Nardi S, Concheri G, Pizzeghello D, et al. Soil organic matter mobilization by root exudates[J]. Chemosphere, 2000, 41(5): 653–658.
[13] Piccolo A, Nardi S, Concheri G. Macromolecular changes of humic substances induced by interaction with organic acids[J]. European Journal of Soil Science, 1996, 47(3): 319–328.
[14] Piccolo A, Nardi S, Concheri G. Micelle-1ike conformation of humic substances as revealed by size exclusion chromatography[J]. Chemosphere, 1996, 33(4): 595–602.
[15] Nardi S, Sessi E, Pizzeghello D, et al. Biological activity of soil organic matter mobilized by root exudates[J]. Chemosphere, 2002, 46(7): 1075–1081.
[16] Wang Q T, Yuan Y S, Zhang Z L, et al. Exudate components mediate soil C dynamic through different priming mechanisms in forest soils[J]. Applied Soil Ecology, 2021, 160: 103855.
[17] Kemmitt S J, Lanyon C V, Waite I S, et al. Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass—a new perspective[J]. Soil Biology and Biochemistry, 2008, 40(1): 61–73.
[18] Jackson L E, Burger M, Cavagnaro T R. Roots, nitrogen transformations, and ecosystem services[J]. Annual Review of Plant Biology, 2008, 59: 341–363.
[19] Cheng W X, Parton W J, Gonzalez-Meler M A, et al. Synthesis and modeling perspectives of rhizosphere priming[J]. The New Phytologist, 2014, 201(1): 31–44.
[20] Rillig M C, Caldwell B A, W?sten H A B, et al. Role of proteins in soil carbon and nitrogen storage: Controls on persistence[J]. Biogeochemistry, 2007, 85(1): 25–44.
[21] Majuakim L, Kitayama K. Influence of polyphenols on soil nitrogen mineralization through the formation of bound protein in tropical montane forests of Mount Kinabalu, Borneo[J]. Soil Biology and Biochemistry, 2013, 57: 14–21.
[22] Phillips R P, Meier I C, Bernhardt E S, et al. Roots and fungi accelerate carbon and nitrogen cycling in forests exposed to elevated CO2[J]. Ecology Letters, 2012, 15(9): 1042–1049.
[23] Kuzyakov Y, Xu X L. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance[J]. The New Phytologist, 2013, 198(3): 656–669.
[24] Meier I C, Finzi A C, Phillips R P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools[J]. Soil Biology and Biochemistry, 2017, 106: 119–128.
[25] Yin H J, Wheeler E, Phillips R P. Root-induced changes in nutrient cycling in forests depend on exudation rates[J]. Soil Biology and Biochemistry, 2014, 78: 213–221.
[26] Lu Y F, Zhang X N, Jiang J F, et al. Effects of the biological nitrification inhibitor 1, 9-decanediol on nitrification and ammonia oxidizers in three agricultural soils[J]. Soil Biology and Biochemistry, 2019, 129: 48–59.
[27] Strickland M S, Wickings K, Bradford M A. The fate of glucose, a low molecular weight compound of root exudates, in the belowground foodweb of forests and pastures[J]. Soil Biology and Biochemistry, 2012, 49: 23–29.
[28] Hao C K, Dungait J A J, Wei X M, et al. Maize root exudate composition alters rhizosphere bacterial community to control hotspots of hydrolase activity in response to nitrogen supply[J]. Soil Biology and Biochemistry, 2022, 170: 108717.
[29] Liu Y, Evans S E, Friesen M L, et al. Root exudates shift how N mineralization and N fixation contribute to the plant-available N supply in low fertility soils[J]. Soil Biology and Biochemistry, 2022, 165: 108541.
[30] Wardle D A, Walker L R, Bardgett R D. Ecosystem properties and forest decline in contrasting long-term chronosequences[J]. Science, 2004, 305(5683): 509–513.
[31] Spohn M, Ermak A, Kuzyakov Y. Microbial gross organic phosphorus mineralization can be stimulated by root exudates–A33P isotopic dilution study[J]. Soil Biology and Biochemistry, 2013, 65: 254–263.
[32] Zhang L, Feng G, Declerck S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium[J]. The ISME Journal, 2018, 12(10): 2339–2351.
[33] Dinkelaker B, Romheld V, Marschner H. Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (L.)[J]. Plant, Cell and Environment, 1989, 12(3): 285–292.
[34] Zhu H, Bing H J, Wu Y H, et al. Low molecular weight organic acids regulate soil phosphorus availability in the soils of subalpine forests, eastern Tibetan Plateau[J]. CATENA, 2021, 203: 105328.
[35] van Hees P A W, Jones D L, Finlay R, et al. The carbon we do not see—The impact of low molecular weight compounds on carbon dynamics and respiration in forest soils: A review[J]. Soil Biology and Biochemistry, 2005, 37(1): 1–13.
[36] Boddy E, Hill P W, Farrar J, et al. Fast turnover of low molecular weight components of the dissolved organic carbon pool of temperate grassland field soils[J]. Soil Biology and Biochemistry, 2007, 39(4): 827–835.
[37] Laanbroek H J. Methane emission from natural wetlands: Interplay between emergent macrophytes and soil microbial processes. A mini-review[J]. Annals of Botany, 2010, 105(1): 141–153.
[38] Girkin N T, Turner B L, Ostle N, et al. Root exudate analogues accelerate CO2and CH4production in tropical peat[J]. Soil Biology and Biochemistry, 2018, 117: 48–55.
[39] Liu Y H, Shahbaz M, Ge T D, et al. Effects of root exudate stoichiometry on CO2emission from paddy soil[J]. European Journal of Soil Biology, 2020, 101: 103247.
[40] 莊姍, 林偉, 丁軍軍, 等. 不同根系分泌物對土壤N2O排放及同位素特征值的影響[J]. 中國農業科學, 2020, 53(9): 1860–1873.
[41] Urbanová M, ?najdr J, Baldrian P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees[J]. Soil Biology and Biochemistry, 2015, 84: 53–64.
[42] Gartzia-Bengoetxea N, Kandeler E, Martínez de Arano I, et al. Soil microbial functional activity is governed by a combination of tree species composition and soil properties in temperate forests[J]. Applied Soil Ecology, 2016, 100: 57–64.
[43] Shi S J, Richardson A E, O'Callaghan M, et al. Effects of selected root exudate components on soil bacterial communities[J]. FEMS Microbiology Ecology, 2011, 77(3): 600–610.
[44] Kong C H, Chen L C, Xu X H, et al. Allelochemicals and activities in a replanted Chinese fir ((Lamb.) Hook) tree ecosystem[J]. Journal of Agricultural and Food Chemistry, 2008, 56(24): 11734–11739.
[45] Fernandez M, Malagoli P, Gallet C, et al. Investigating the role of root exudates in the interaction between oak seedlings and purple moor grass in temperate forest[J]. Forest Ecology and Management, 2021, 491: 119175.
[46] Xia Z C, Yu L, He Y, et al. Broadleaf trees mediate chemically the growth of Chinese fir through root exudates[J]. Biology and Fertility of Soils, 2019, 55(7): 737–749.
[47] Vyvyan J R. Allelochemicals as leads for new herbicides and agrochemicals[J]. Tetrahedron, 2002, 58(9): 1631–1646.
[48] Schmidt J E, Bowles T M, Gaudin A C M. Using ancient traits to convert soil health into crop yield: Impact of selection on maize root and rhizosphere function[J]. Frontiers in Plant Science, 2016, 7: 373.
[49] Xun W B, Shao J H, Shen Q R, et al. Rhizosphere microbiome: Functional compensatory assembly for plant fitness[J]. Computational and Structural Biotechnology Journal, 2021, 19: 5487–5493.
[50] Chaparro J M, Badri D V, Vivanco J M. Rhizosphere microbiome assemblage is affected by plant development[J]. The ISME Journal, 2014, 8(4): 790–803.
[51] Schreiter S, Ding G C, Heuer H, et al. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce[J]. Frontiers in Microbiology, 2014, 5: 144.
[52] Liu J L, Ha V N, Shen Z, et al. Response of the rhizosphere microbial community to fine root and soil parameters followingL. afforestation[J]. Applied Soil Ecology, 2018, 132: 11–19.
[53] Mehmood A, Hussain A, Irshad M, et al. Cinnamic acid as an inhibitor of growth, flavonoids exudation and endophytic fungus colonization in maize root[J]. Plant Physiology and Biochemistry, 2019, 135: 61–68.
[54] Yuan J, Wu Y C, Zhao M L, et al. Effect of phenolic acids from banana root exudates on root colonization and pathogen suppressive properties ofNJN-6[J]. Biological Control, 2018, 125: 131–137.
[55] Shi S J, Herman D J, He Z L, et al. Plant roots alter microbial functional genes supporting root litter decomposition[J]. Soil Biology and Biochemistry, 2018, 127: 90–99.
[56] Huang A C, Jiang T, Liu Y X, et al. A specialized metabolic network selectively modulatesroot microbiota[J]. Science, 2019, 364(6440): eaau6389.
[57] Wen T, Yuan J, He X M, et al. Enrichment of beneficial cucumber rhizosphere microbes mediated by organic acid secretion[J]. Horticulture Research, 2020, 7: 154.
[58] Sasse J, Martinoia E, Northen T. Feed your friends: Do plant exudates shape the root microbiome?[J]. Trends in Plant Science, 2018, 23(1): 25–41.
[59] Wang H W, Ma C Y, Xu F J, et al. Root endophyte- enhanced peanut-rhizobia interaction is associated with regulation of root exudates[J]. Microbiological Research, 2021, 250: 126765.
[60] Kalu C M, Oduor Ogola H J, Selvarajan R, et al. Fungal and metabolome diversity of the rhizosphere and endosphere ofin an AMD-polluted environment[J]. Heliyon, 2021, 7(3): e06399.
[61] 潘聲旺, 雷志華, 吳云霄, 等. 根系分泌物在有機氯農藥殘留降解過程中的作用[J]. 云南大學學報(自然科學版), 2017, 39(4): 669–676.
[62] Cusack D F, Silver W L, Torn M S, et al. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests[J]. Ecology, 2011, 92(3): 621–632.
[63] Burns R G, DeForest J L, Marxsen J, et al. Soil enzymes in a changing environment: Current knowledge and future directions[J]. Soil Biology and Biochemistry, 2013, 58: 216–234.
[64] Dennis P G, Miller A J, Hirsch P R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities?[J]. FEMS Microbiology Ecology, 2010, 72(3): 313–327.
[65] Oburger E, Jones D L. Sampling root exudates–mission impossible?[J]. Rhizosphere, 2018, 6: 116–133.
[66] Zhang X C, Dippold M A, Kuzyakov Y, et al. Spatial pattern of enzyme activities depends on root exudate composition[J]. Soil Biology and Biochemistry, 2019, 133: 83–93.
[67] 司鵬, 邵微, 于會麗, 等. 小分子有機物對土壤酶活性及微生物多樣性的影響[J]. 中國土壤與肥料, 2019(2): 75–82.
[68] 張亮, 夏文靜, 周惠, 等. 半夏塊莖提取液對土壤微生物和酶活性的影響[J]. 貴州師范大學學報(自然科學版), 2019, 37(2): 87–92.
[69] Wang C Q, Xue L, Dong Y H, et al. Effects of stand density on soil microbial community composition and enzyme activities in subtropical(Lamb.) Hook plantations[J]. Forest Ecology and Management, 2021, 479: 118559.
[70] 王小平, 肖肖, 唐天文, 等. 連香樹人工林根系分泌物輸入季節性變化及其驅動的根際微生物特性研究[J]. 植物研究, 2018, 38(1): 47–55.
[71] 李軍, 趙彩平. 不同林分下樟子松根系分泌物對土壤環境及微生物的影響[J]. 山東農業大學學報(自然科學版), 2022, 53(3): 380–385.
[72] Proctor C, He Y H. Quantifying root extracts and exudates of sedge and shrub in relation to root morphology[J]. Soil Biology and Biochemistry, 2017, 114: 168–180.
[73] Williams A, Langridge H, Straathof A L, et al. Comparing root exudate collection techniques: An improved hybrid method[J]. Soil Biology and Biochemistry, 2021, 161: 108391.
[74] Lu H N, Sun J T, Zhu L Z. The role of artificial root exudate components in facilitating the degradation of pyrene in soil[J]. Scientific Reports, 2017, 7(1): 1–10.
[75] Bi B Y, Tong Q, Wan C Y, et al.var.mediates interspecific belowground chemical interactions through root exudates[J]. Forest Ecology and Management, 2022, 511: 120158.
[76] Li S L, Zhu N L, Tang C P, et al. Differential distribution of characteristic constituents in root, stem and leaf tissues ofusing MALDI mass spectrometry imaging[J]. Fitoterapia, 2020, 146: 104679.
[77] Li B, Neumann E K, Ge J Y, et al. Interrogation of spatial metabolome ofwith high-resolution matrix- assisted laser desorption/ionization and laser desorption/ ionization mass spectrometry imaging[J]. Plant, Cell & Environment, 2018, 41(11): 2693–2703.
[78] 張騰. 馬尾松和杉木對植物的化感作用研究[D]. 杭州: 浙江大學, 2016.
[79] 俞元春, 余健, 房莉, 等. 缺磷脅迫下馬尾松和杉木苗根系有機酸的分泌[J]. 南京林業大學學報(自然科學版), 2007, 31(2): 9–12.
[80] 朱旭恒. 不同逆境脅迫對杉木幼苗生理指標及根系分泌物的影響[D]. 福州: 福建農林大學, 2009.
[81] 熊德成, 黃錦學, 楊智杰, 等. 增溫對杉木幼樹細根分泌物的影響研究初報[J]. 亞熱帶資源與環境學報, 2016, 11(4): 85–88, 92.
[82] Xia Z C, Kong C H, Chen L C, et al. Allelochemical- mediated soil microbial community in long-term monospecific Chinese fir forest plantations[J]. Applied Soil Ecology, 2015, 96: 52–59.
[83] Chen L C, Wang S L, Wang P, et al. Autoinhibition and soil allelochemical (cyclic dipeptide) levels in replanted Chinese fir () plantations[J]. Plant and Soil, 2014, 374(1): 793–801.
[84] 黃龍, 包維楷, 李芳蘭, 等. 土壤結構和植被對土壤微生物群落的影響[J]. 應用與環境生物學報, 2021, 27(6): 1725–1731.
Root Exudates’ Roles and Analytical Techniques Progress
CHEN Hong1, 2, TANG Haoye2, GUO Jiahuan1, PAN Chang1, WANG Ruhai2, WU Yonghong2, YU Yuanchun1*
(1 Co-Innovation Center for the Sustainable Forestry in Southern China, College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China; 2 Soil and Environment Analysis Center, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China)
Considerable plant root exudates are the key factor to maintain the vitality of rhizosphere microecosystem and an important part of rhizosphere material cycle, which plays an important driving role in material cycle in soil ecological environment. Plants release root exudates that stimulate microbial growth, activity, and turnover to accelerate nutrient cycling in the rhizosphere, increase nutrient utilization in soil ecosystem, and cause changes in CO2and CH4fluxes in small-scale space. Different plants release various root secretion for long-term survival nutrients between species, but some can secrete harmful allelochemicals to kill other sensitive plants, even which are poison to themselves or causing continuous cropping obstacle, etc. They could also affect soil microbial community through root exudates to exclude other plants, and finally realize self-survival.Plants’ healthy growth depend on their interactions with the complex dynamic community of soil microorganisms, but the rhizosphere microbial community structure and composition is affected by the plant species and growth phases, soil properties, function genes, etc., the dynamic changes of these factors may lead to the diversification of root exudates, thus forming a complex and variable relationship between root exudates and plants, and affecting the healthy growth of plants. Therefore, the research on plant root exudates is a hot topic in the fields of soil ecology, plant nutrition and metabolism. With the continuous development of analytical techniques, the research on plant root exudates is getting more and more detailed, which has certain guiding significance for the production of agriculture, forestry and other industries.
Root exudates; Analytical technique; Soil; Microorganism
S714;O65
A
10.13758/j.cnki.tr.2023.02.001
陳虹, 唐昊冶, 郭家歡, 等. 根系分泌物主要作用及解析技術進展. 土壤, 2023, 55(2): 225–233.
國家重點研發計劃項目(2021YFD2201304)資助。
(ycyu@njfu.edu.cn)
陳虹(1977—),女,江蘇沭陽人,博士研究生,高級工程師,研究方向為分析化學、土壤生態學。E-mail: chenhong@issas.ac.cn
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
