泌乳高峰期娟犏牛和牦牛乳汁差異蛋白分析
2023-06-05 06:23:44潘陽陽王立斌邱山桐余四九徐庚全
核農學報 2023年7期
關鍵詞:分析
高 娜 潘陽陽 王 萌 王立斌 邱山桐 余 震 余四九 徐庚全
(甘肅農業大學動物醫學院,甘肅 蘭州 730070)
牦牛(Bos grunniens)是高海拔高寒地區重要的生物物種[1]。牦牛奶具有天然濃縮乳之稱,易消化吸收,奶中蛋白質具有提高機體免疫力、預防心腦血管疾病和促進嬰幼兒生長發育等功效[2]。隨著我國國民生活水平的提高,人們對奶及奶制品的需求急劇增加,但牦牛生產性能較差,品種原始,泌乳量較其他牛低,不能滿足人們對高品質牦牛奶日益增長的需求。為提高牦牛奶產量,甘肅農業大學余四九團隊成功利用娟姍牛及其凍精雜交改良甘南牦牛,繁育生產出優質娟犏牛(Bos taurus)[3],其耐粗飼料、生長發育快,對環境的適應性強。對娟犏牛乳的營養成分進行測定,發現其乳蛋白質、乳脂率和非乳脂固體等乳成分高于當地黃犏牛乳,接近優質牦牛乳品質[4],且娟犏牛初胎的擠奶量是牦牛的4~5 倍[5]。然而,娟犏牛乳品質與牦牛乳是否有差異還需進一步研究。
牛奶均能夠為機體提供酶、激素和免疫成分等,這些營養成分均對嬰幼兒的生長發育有著極其重要的功能。近年來,隨著蛋白質組學技術的不斷發展,在牦牛[6]、人[7]、山羊[8]、驢[9]和奶牛[10]等物種的乳汁中鑒定到了多種蛋白質。Li 等[11]采用串聯質譜(tandem mass tag,TMT)標記蛋白質組學方法在Murrah、Nili Ravi 和Mediterranean 水牛乳清蛋白中共鑒定到580 種蛋白質。Yang 等[6]基于同位素標記相對和絕對定量(isobaric tags for relative and absolute quantification,iTRAQ)技術,采用蛋白質組學方法分析了牛、牦牛、水牛、山羊和駱駝的乳汁中的乳清蛋白質組。Jin 等[7]利用數據獨立采集(data-independent acquisition,DIA)方法確定了母乳乳清蛋白組的差異,并評估了哺乳期對母乳的影響。但鮮見利用TMT 標記蛋白質組學方法分析泌乳高峰期娟犏牛奶和牦牛奶乳清與乳脂球膜(milk fat globule membrane,MFGM)蛋白的研究報道。
鑒于此,本試驗運用TMT 標記蛋白質組學的方法,通過基因本體(gene ontology,GO)分析差異表達蛋白(differentially expressed proreins,DEPs)的細胞組成、分子功能及其參與的生物過程,通過京都基因與基因組百科全書(kyoto encyclopedia of genes and genomes,KEGG)分析DEPs 參與的主要信號通路,通過蛋白質-蛋白質相互作用網絡(protein-protein interaction,PPI)探究DEPs間的功能相關性,比較泌乳高峰期娟犏牛奶與牦牛奶乳清和MFGM 蛋白表達的差異,旨在為高原奶制品的開發提供幫助,為生產接近母乳的功能性嬰幼兒配方乳粉提供科學依據。
1 材料與方法
1.1 試驗材料
1.1.1 樣品采集 于2022 年8 月采集處于泌乳高峰期娟犏牛奶和牦牛奶各30 份,均采自甘肅省甘南州夏河縣阿木去乎鎮牙力吉鄉。娟犏牛和牦牛兩者屬于草原散養。采集樣本每份均來自不同奶牛,每只牛采集200 mL 奶樣。所采泌乳高峰期娟犏牛奶樣與牦牛奶樣各隨機平均分為3 組,每組10 份樣本,3 組間互為重復。所有樣本低溫運回實驗室,-80 ℃超低溫冰箱保存備用。具體采樣信息見電子附表1。
1.1.2 主要試劑 TMTpro 16 標記試劑盒、二喹啉甲酸(bicinchonininc acid,BCA)試劑盒、質譜級與色譜級乙腈(acetonitrile,ACN)、甲酸(formic acid,FA)、未染色蛋白分子量標記和質譜(mass spectrum,MS)級水,上海ThermoScientific 公司;氯化鈉、三(羥甲基)氨基甲烷鹽酸鹽(tris hydrochloride,Tris-HCl)、苯甲基磺酰氟(phenylmethyl sulfonyl fluoride,PMSF)、二硫蘇糖醇(dithiothreitol,DTT)、甘油、3-吲哚乙酸(3-indole acetic acid,IAA),上海生工生物工程股份有限公司;十二烷基磺酸鈉(sodium dodecyl sulfate,SDS)裂解液,上海碧云天生物技術有限公司;預制膠,南京金斯瑞生物科技股份有限公司提供;胰酶,上海華利世科技有限公司;四乙基溴化銨(tetraethylammonium bromide,TEAB),美國Sigma公司;丙酮(色譜純),北京沃凱生物科技有限公司。
1.1.3 儀器與設備 Q Exactive HF 質譜儀、EasynLC 1000 液相系統,上海ThermoFisher 公司;TGL-16A臺式冷凍離心機,上海盧湘儀離心機儀器有限公司;SDS-PAGE 凝膠電泳儀,北京市六一儀器廠;ST-360酶標儀,上海科華實驗系統有限公司;JXH-100恒溫混勻儀,上海凈信實業發展有限公司;Tanon-1600 數碼凝膠圖像分析儀器,上海天能科技有限公司;SCIENTZ-10N 凍干機,寧波新芝生物科技有限公司;高pH 分離液相色譜儀,上海Agilent 公司;MCCW V1乳成分分析儀,鄭州兆為儀器設備有限公司。
1.2 奶品TMT定量蛋白質組學試驗方法
參照邱山桐等[12]營養成分測定、乳清與乳脂的分離、乳清蛋白和MFGM 蛋白的提取、胰蛋白酶酶解與肽段標記、MS分析、數據庫檢索方法。
1.3 泌乳高峰期娟犏牛奶與牦牛奶乳清和MFGM 蛋白質差異分析
在定量結果的顯著性差異分析中,本試驗篩選樣本組內3 次重復數據進行統計分析,在1.2 差異倍數(fold change,FC)且P<0.05 條件下進行DEPs 篩選(即FC≥1.2 為上調,FC≤0.8 為下調,0.8<FC<1.2 無明顯變化)。采用兩組樣本間的蛋白質表達差異倍數和t-檢驗(t-test)得到的P兩個因素,利用ggplot2 繪圖軟件共同繪制火山圖。
1.4 泌乳高峰期娟犏牛奶與牦牛奶乳清和MFGM 蛋白生物信息學分析
利用數據庫檢索得到原始數據后,按照Score Sequest HT>0且unique peptide≥1,并去除空白值的標準篩選得到可信蛋白。利用可信蛋白的表達量進行主成分分析(principal component analysis,PCA)。用Cluster 3.0軟件對已鑒定的蛋白進行分層聚類。運用生物信息學在線資源[13](http://www.genome.jp)對DEPs進行GO[14]注釋,篩選生物過程、分子功能和細胞組成3種分類中對應差異蛋白數目大于1的GO條目。利用在線資源(http://www.genome.jp/kegg/pathway.html)對差異蛋白進行KEGG[15]路徑分析,以深入了解蛋白質生物學功能,從而找到富集的代謝通路條目。將乳清和MFGM DEPs提交到STRING(https://cn.string-db.org/)在線數據庫進行分析,獲得PPI[16]數據。利用ggplot2繪圖軟件繪制聚類圖、GO富集圖和KEGG通路圖。
2 結果與分析
2.1 泌乳高峰期娟犏牛與牦牛乳汁成分分析
2.1.1 泌乳高峰期娟犏牛奶與牦牛奶的主要乳成分含量 由表1 可知,娟犏牛乳和牦牛乳的乳脂、乳蛋白和乳糖含量均存在顯著差異。
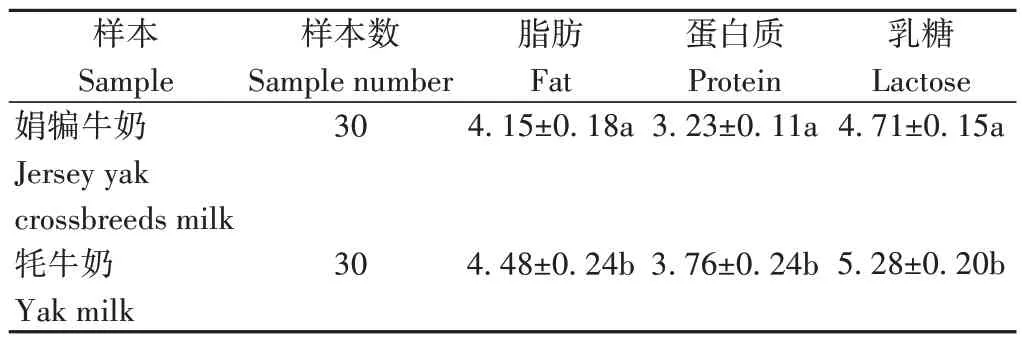
表1 娟犏牛奶與牦牛奶的主要乳成分含量Table 1 Comparison of main milk components between jersey yak crossbreeds milk and yak milk/%
2.1.2 娟犏牛奶與牦牛奶乳清和MFGM 蛋白質鑒定結果 泌乳高峰期娟犏牛奶與牦牛奶乳清蛋白中共鑒定出肽段2 991 個,MFGM 蛋白中共鑒定出肽段4 470個。娟犏牛奶與牦牛奶中共鑒定到651個乳清蛋白,獲得613個乳清可信蛋白,根據FC≥1.2為上調,FC≤0.8 為下調,從可信蛋白中篩選出145 個乳清DEPs,相比于牦牛奶乳清DEPs,娟犏牛奶乳清DEPs 中71 個上調,74 個下調。泌乳高峰期娟犏牛奶和牦牛奶中共鑒定到990 個MFGM 蛋白,獲得956 個MFGM 可信蛋白,根據FC≥1.2 為上調,FC≤0.8 為下調,從可信蛋白中篩選出160 個DEPs,相比于牦牛奶MFGM DEPs,娟犏牛奶MFGM DEPs中78個上調,82個下調。
2.1.3 泌乳高峰期娟犏牛奶與牦牛奶定量乳清和MFGM可信白PCA分析 在泌乳高峰期娟犏牛奶與牦牛奶中共篩選出613個乳清可信蛋白,956個MFGM 可信蛋白。從圖1-A 可知,乳清中第1主坐標(PC1)對于區分樣品的貢獻率為61.11%,第2 主坐標(PC2)對于區分樣品的貢獻率為14.48%;從圖1-B可知,MFGM中第1 主坐標(PC1)對于區分樣品的貢獻率為59.37%,第2 主坐標(PC2)對于區分樣品的貢獻率為16.64%。綜合上述結果可知,兩組奶樣同組間的分離度較低,重復性較好;同時兩種奶樣間分離度較高,說明兩種奶樣蛋白質間具有一定差異性。

圖1 娟犏牛奶與牦牛奶樣本可信蛋白PCA分析圖Fig.1 PCA analysis diagram of credible protein in jersey yak crossbreeds milk and yak milk samples
2.1.4 泌乳高峰期娟犏牛奶與牦牛奶主要乳清和MFGM DEPs
2.1.4.1 泌乳高峰期娟犏牛奶與牦牛奶主要乳清DEPs 相比于牦牛奶,娟犏牛奶乳清中上調的高豐度DEPs主要包括骨橋蛋白(osteopontin,OPN)、纖溶酶原(plasminogen,PLG)、補體C7(complement component 7,C7)、甘露糖結合蛋白C(mannose-binding protein C,MBP-C)和補體C4A(complement component 4A,C4A)等;乳清中下調的高豐度DEPs 主要包括熱休克同源71 kDa 蛋白(heat shock cognate 71 kDa protein,HSP7C)、CD59 糖蛋白(CD59 glycoprotein,CD59)和黃嘌呤脫氫酶(xanthine dehydrogenase,XDH)等,詳細信息見表2。
2.1.4.2 泌乳高峰期娟犏牛奶與牦牛奶主要MFGM DEPs 相比于牦牛奶,娟犏牛奶MFGM 中上調的高豐度DEPs 主要包括OPN、C7 和α1-酸性糖蛋白(alpha-1-acid glycoprotein,AGP),MFGM 中下調的高豐度DEPs 主要包括HSP7C、XDH 和E1A 激活基因阻遏子1(cellular repressor of E1A-stimulated genes 1,CREG1),詳細信息見表3。
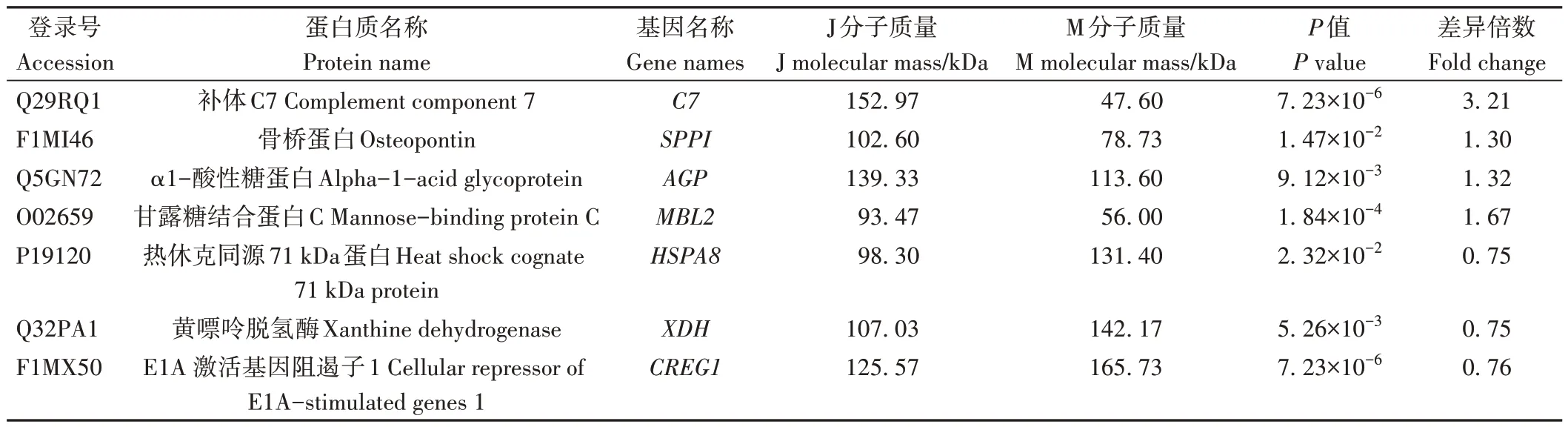
表3 娟犏牛奶與牦牛奶主要MFGM DEPs鑒定信息Table 3 Identification information of main MFGM DEPs in jersey yak crossbreeds milk and yak milk
2.2 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs 生物信息學分析
2.2.1 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs GO功能注釋 對泌乳高峰期娟犏牛奶與牦牛奶中的145種乳清DEPs進行富集分析,發現只有119種參與GO功能注釋,乳清DEPs共富集482個生物過程,上調與下調DEPs參與最多的生物過程是免疫反應;共富集132個細胞成分,上調與下調DEPs參與最多的細胞成分是細胞外區域與細胞外間隙;共富集179 個分子功能,上調與下調DEPs 參與最多的分子功能是蛋白質同二聚體活性。由圖2 可知,生物過程功能分類中,泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs在固有免疫應答、補體激活,經典途徑和革蘭氏陽性菌的防御反應中富集度較高,各有9、4和4個娟犏牛奶乳清差異表達上調蛋白富集,2、1和1個下調DEPs。細胞組分功能分類中,細胞外間隙和細胞外區域DEPs占比最多。分子功能分類中,蛋白質同源二聚體活性和絲氨酸型內肽酶抑制劑活性DEPs富集度較高。本研究分析發現娟犏牛奶中的上調乳清核心DEPs主要集中在細胞外間隙的固有免疫應答。
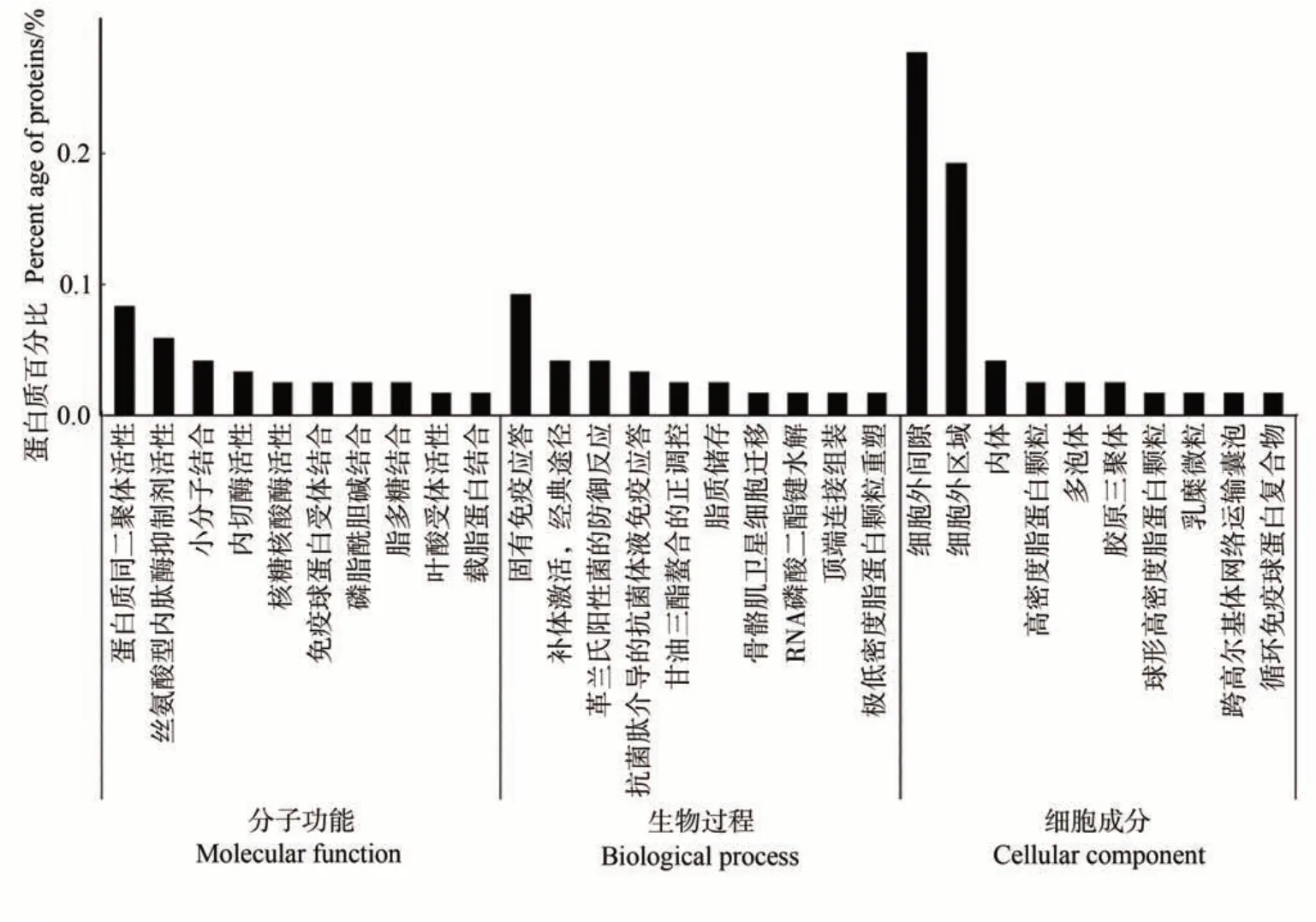
圖2 娟犏牛奶與牦牛奶乳清DEPs的GO注釋Fig.2 Go annotation of DEPs whey between jersey yak crossbreeds milk and yak milk
GO富集結果說明,泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs具有不同免疫功能的原因,可能是由于娟犏牛奶與牦牛奶在細胞外間隙和細胞外區域等細胞組分的結合能力不同,通過催化蛋白質同源二聚體活性和絲氨酸型內肽酶抑制劑活性等,造成對固有免疫應答、補體激活,經典途徑和革蘭氏陽性菌的防御反應等途徑的不同反應,從而導致免疫功能不同。
2.2.2 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs KEGG 代謝通路分析 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs 有71 種參與KEGG 通路富集分析。KEGG富集分析將泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs分為細胞過程、環境信息處理、人類疾病、組織系統和新陳代謝5個部分,共富集到156條通路。細胞過程富集11 條、新陳代謝富集26 條、環境信息處理富集19條、人類疾病富集50 條和組織系統富集50 條,其中DEPs富集的50條組織系統中有17條是免疫系統。娟犏牛奶與牦牛奶乳清DEPs 富集最多的前20 條通路如圖3 所示。娟犏牛奶與牦牛奶乳清DEPs 主要富集的途徑包括金黃色葡萄球菌感染、PI3K-Akt 信號通路、肺結核和補體與凝血級聯,各有12、10、10 和9 個娟犏牛奶乳清DEPs富集。
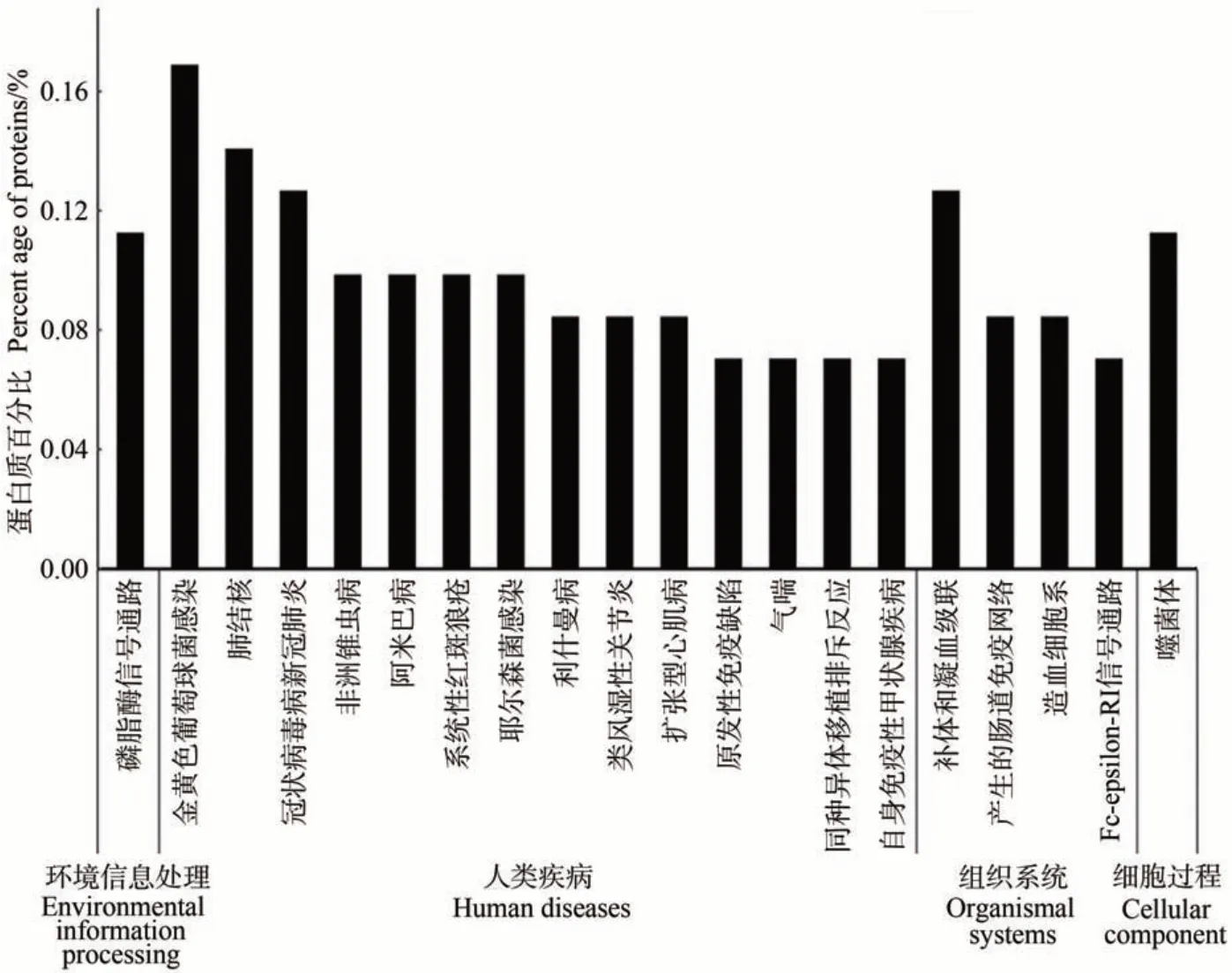
圖3 娟犏牛奶與牦牛奶乳清DEPs的KEGG途徑分析(前20條)Fig.3 KEGG pathway analysis of DEPs whey between jersey yak crossbreeds milk and yak milk (top 20)
娟犏牛奶乳清DEPs上調的29種里有16種富集在免疫系統,占55.2%,下調42 種里有7 種參與免疫系統,占16.7%,且免疫系統中上調DEPs 總豐度高于下調的,說明娟犏牛奶乳清比牦牛奶擁有更多參與免疫的DEPs。除S100A8 外,其余上調的17 種娟犏牛奶乳清DEPs 均極顯著表達;除GNG12 和PIGR 外,其余下調的7 種均極顯著表達(P<0.01)(表4)。17 條免疫系統中蛋白富集最多的通路是補體和凝血級聯(圖4),此途徑中上調的DEPs 有8 種,分別是C4A、MBP-C、C7、補體因子I(complement factor I,CFI)、從生蛋白(clusterin,CLU)、α2-巨球蛋白(alpha-2-macroglobulin,A2M)、纖維蛋白原α鏈(fibrinogen alpha chain,FGA)和PLG,占富集在免疫系統上調DEPs 的44.4%,下調的DEPs 是CD59,占富集在免疫系統下調DEPs 的11.1%,對娟犏牛奶中的乳清核心DEPs 進行富集分析,發現在補體和凝血級聯通路富集的上調乳清核心DEPs最多,主要是FGA、C4A、PLG、A2M和CLU。
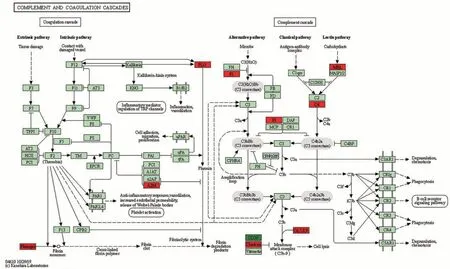
圖4 娟犏牛奶與牦牛奶乳清DEPs補體和凝血級聯代謝通路圖Fig.4 Metabolic pathway diagram of complement and coagulation cascade of jerseyyak crossbreeds milk and yak milk whey DEPs

表4 參與免疫系統的娟犏牛奶與牦牛奶乳清DEPs鑒定信息Table 4 Identification information of DEPs whey between jersey yak crossbreeds milk and yak milk involved in immune system
2.2.3 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs PPI分析 泌乳高峰期娟犏牛奶與牦牛奶乳清DEPs PPI分析得到145 個節點和882 個互作邊,連接度排名前25 的乳清DEPs 得到結果如圖5 所示。與牦牛奶乳清DEPs相比,娟犏牛奶乳清中連接度靠前且表達上調的CLU、PLG、A2M、FGA、C4A 和免疫球蛋白J 鏈(immunoglobulin J chain,JCHAIN)等DEPs 與多種蛋白相互關聯;連接度靠前且表達下調的HSP7C、轉化蛋白RhoA(transforming protein RhoA,RHOA)和CD59 等DEPs 與多種蛋白相互關聯,推測這些DEPs 可能是泌乳高峰期娟犏牛奶乳清的核心DEPs。
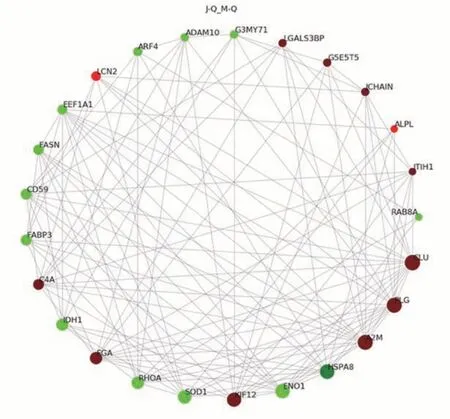
圖5 娟犏牛奶與牦牛奶乳清DEPs STRING相互作用網絡分析Fig.5 STRING interaction network analysis of DEPs whey between jersey yak crossbreeds milk and yak milk
2.3 泌乳高峰期娟犏牛奶與牦牛奶MFGM DEPs 生物信息學分析
2.3.1 泌乳高峰期娟犏牛奶與牦牛奶MFGM 顯著DEPs GO 功能注釋 對泌乳高峰期娟犏牛奶與牦牛奶中的160 種MFGM DEPs 進行富集分析,發現只有131種參與GO 功能注釋,MFGM DEPs 共富集507個生物過程,上調與下調DEPs 參與最多的生物過程主要是免疫反應;共富集132 個細胞成分,上調與下調DEPs 參與最多的細胞成分是細胞外區域與細胞外間隙;共富集207個分子功能,上調與下調DEPs參與最多的分子功能是鳥苷-5′-三磷酸酶(guanosine 5′-triphosphatase,GTPase)活性與GTP 結合。由圖6 可知,生物過程功能分類中,泌乳高峰期娟犏牛奶與牦牛奶MFGM DEPs在固有免疫應答、補體激活,經典途徑和革蘭氏陽性菌的防御反應中富集度較高,各有8、4和1個娟犏牛奶MFGM 差異表達上調蛋白富集,2、2和1個下調DEPs。細胞組分功能分類中,細胞外間隙和細胞外區域DEPs極顯著富集,且占比最高。分子功能分類中,GTPase活性、GTP結合和絲氨酸型內肽酶抑制劑活性DEPs富集度較高。綜上,娟犏牛奶中的上調MFGM核心DEPs主要集中在細胞外間隙的固有免疫應答。
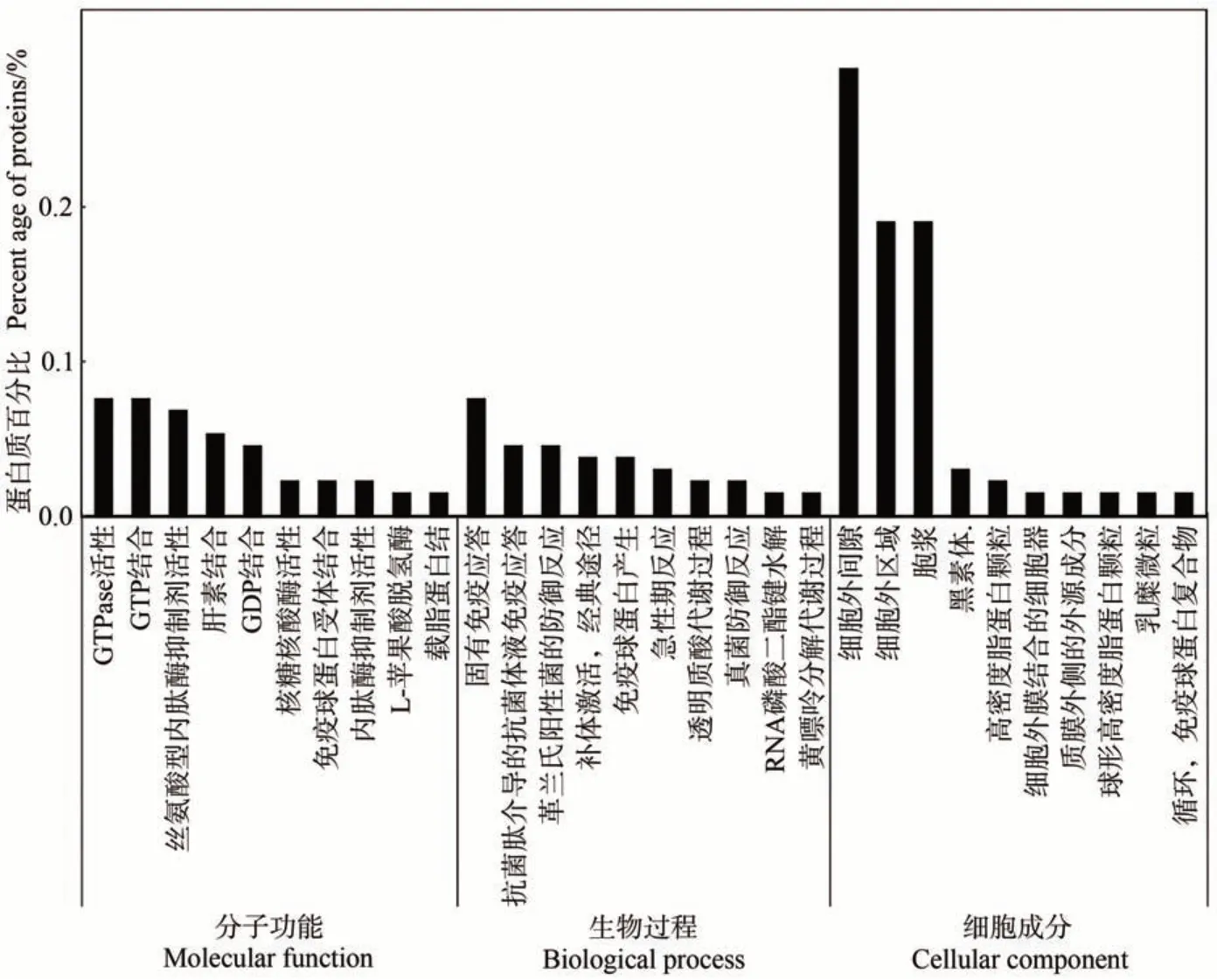
圖6 娟犏牛奶與牦牛奶的MFGM DEPs的GO注釋Fig.6 Go annotation of DEPs MFGM between jersey yak crossbreeds milk and yak milk
2.3.2 泌乳高峰期娟犏牛奶與牦牛奶MFGM DEPs KEGG 代謝通路分析 泌乳高峰期娟犏牛奶與牦牛奶MFGM DEPs 有74 種參與KEGG 通路富集分析。KEGG 途徑分析將娟犏牛奶與牦牛奶MFGM DEPs 分為細胞過程、環境信息處理、人類疾病、新陳代謝和組織系統5 個部分,共富集到181 條通路。細胞過程富集16 條、環境信息處理富集21 條、人類疾病富集55 條、新陳代謝富集32 條和組織系統富集57 條,其中富集到組織系統的57 條中有15 條是免疫系統。娟犏牛奶與牦牛奶MFGM DEPs 富集最多的前20 條通路結果如圖7 所示。娟犏牛奶與牦牛奶MFGM DEPs 富集的途徑主要是金黃色葡萄球菌感染、補體與凝血級聯反應、肌動蛋白細胞骨架的調節和肺結核,各有11、10、9和8個DEPs富集。
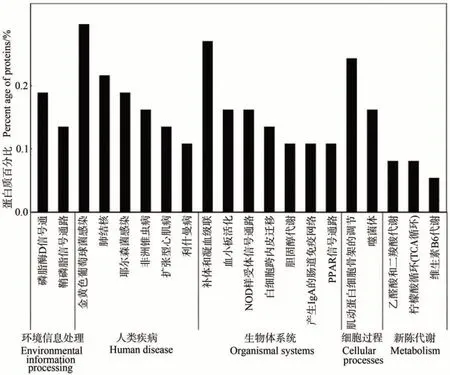
圖7 娟犏牛奶與牦牛奶的MFGM DEPs的KEGG途徑分析(前20條)Fig.7 KEGG pathway analysis of DEPs MFGM between jersey yak crossbreeds milk and yak milk (top 20)
娟犏牛奶MFGM DEPs 上調的31 種中,有14 種富集在免疫系統,占45.1%;下調的43種中,有11種參與免疫系統,占25.5%,且免疫系統中上調DEPs 總豐度高于下調DEPs(表5),說明娟犏牛奶MFGM 比牦牛奶擁有更多參與免疫的DEPs。16 條免疫系統中蛋白富集最多的通路是補體和凝血級聯,此途徑中上調的DEPs 有10 種,分別是C4A、MBP-C、C7、CFI、CLU、A2M、FGA、激肽原-1(kininogen-1,KNG1)、補體因子H(complement factor H,CFH)和PLG,占富集在免疫系統上調DEPs 的62.5%,對娟犏牛奶中的MFGM 核心DEPs 進行分析,發現在補體和凝血級聯通路富集的上調MFGM 核心DEPs 最多,主要是FGA、C4A、PLG、A2M和CLU。
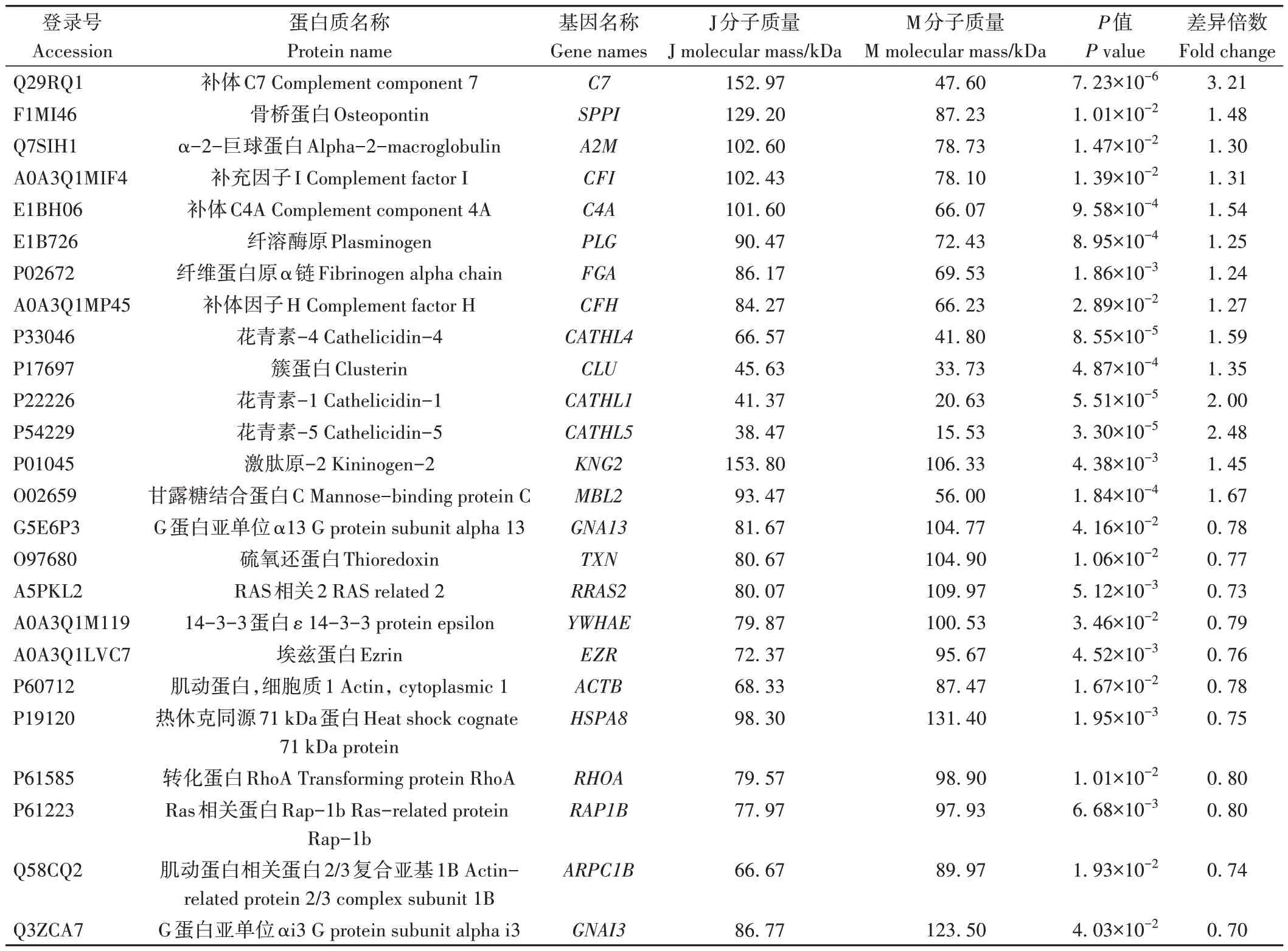
表5 參與免疫系統的娟犏牛奶與牦牛奶的MFGM DEPs鑒定信息Table 5 Identification information of DEPs MFGM between jersey yak crossbreeds milk and yak milk involved in immune system
2.3.3 泌乳高峰期娟犏牛奶與牦牛奶MFGM 顯著DEPs PPI 分析 泌乳高峰期娟犏牛奶與牦牛奶MFGM DEPs PPI 分析后得到160 個節點和1 232 個互作邊,連接度排名前25的DEPs如圖8所示。與牦牛奶MFGM DEPs 相比,娟犏牛奶MFGM 中連接度靠前且表達上調的結合珠蛋白(haptoglobin,HP)、A2M、CLU、PLG和AGP等DEPs與多種蛋白相互關聯;連接度靠前且表達下調的HSP7C 和RHOA 等DEPs 與多種蛋白相互關聯,推測這些DEPs 可能是泌乳高峰期娟犏牛奶MGFM的核心DEPs。
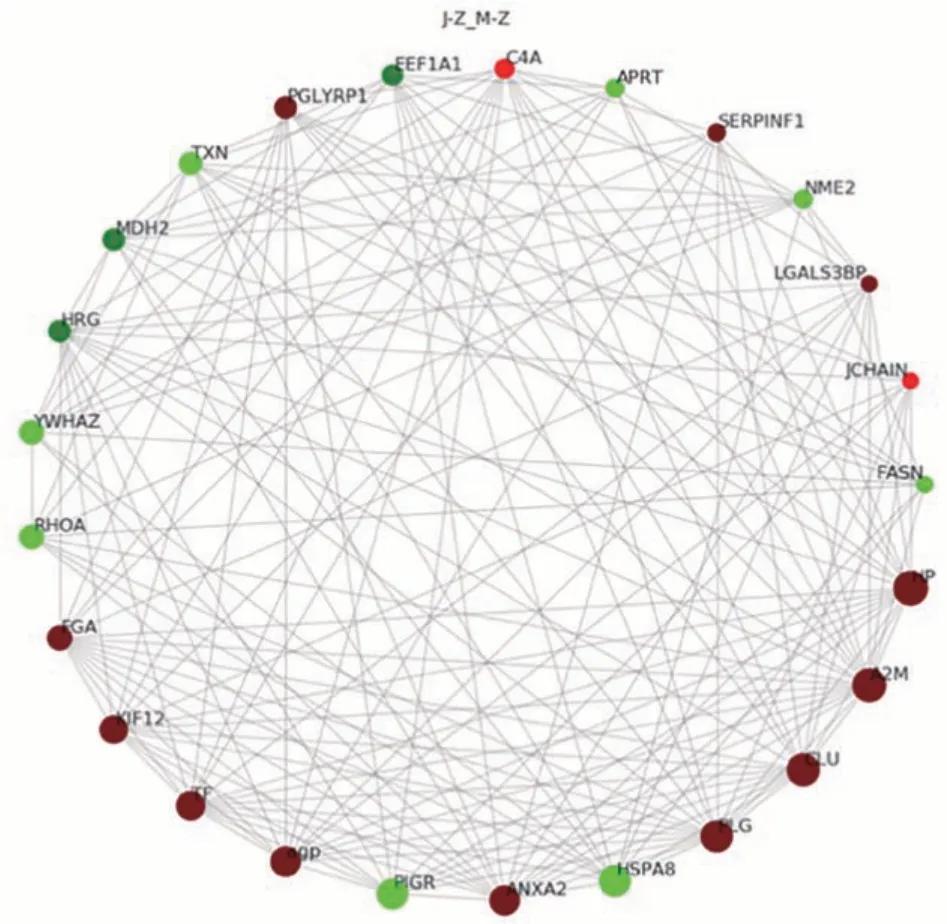
圖8 娟犏牛奶與牦牛奶的MFGM DEPs STRING 相互作用網絡分析Fig.8 STRING interaction network analysis of DEPs MFGM between jersey yak crossbreeds milk and yak milk
3 討論
對娟犏牛乳營養成分測定,發現乳成分接近優質牦牛乳品質[4],且娟犏雌牛的擠奶量7.5~10 kg,是當地牦雌牛(平均1.4 kg)的4~5倍[5],娟犏牛的產奶量遠高于牦牛。但泌乳高峰期娟犏牛奶與牦牛奶乳成分測定結果表明,娟犏牛奶的乳糖率、乳脂率和乳蛋白率均比牦牛奶低。這與吳富鑫等[17]研究的高產奶牛由于“稀釋作用”的影響,乳糖率、乳脂率和乳蛋白率可能更低的結果相一致。
目前,通過蛋白質組學方法對牦牛奶乳清蛋白[18]和MFGM 蛋白[19]已有相關報道,而對娟犏牛奶乳清蛋白和MFGM 蛋白鮮有研究。本研究中,娟犏牛奶與牦牛奶相比,主要乳清與MFGM 高豐度上調DEPs 包括MBP-C、C4A、C7、PLG、OPN 和AGP 等,娟犏牛奶乳清與MFGM 中的上調核心DEPs 包括HP、CLU、PLG、A2M、FGA、AGP 和C4A 等。對這些DEPs 分析發現MBP-C、C4A、C7、A2M 和PLG 都富集在補體與凝血級聯通路。C7 是補體與凝血級聯反應的終末產物,作為補體系統固有成分之一,是形成攻膜復合物(membrace attack complex,MAC)的主要限速因子[20]。MBP-C、C4A 和C7富集的甘露糖結合凝集素通道最終會聚合生成MAC,MAC 在抗炎中發揮重要作用,具有免疫保護功能,并在感染細胞膜內形成跨膜通道,分別導致腫瘤細胞及細菌溶解[21]。A2M 通過抑制侵入機體病原體釋放的多種蛋白酶與毒素,減少其對宿主的破壞,增強宿主的抗病力。纖溶系統在血栓、止血和傷口愈合中扮演重要角色,而PLG 是纖溶酶的酶原形式,能夠降解纖維蛋白,在纖溶系統中起關鍵作用[22]。OPN參與人體多種生理和病理過程,如骨重塑、傷口愈合和免疫調節等[23],也可能通過改變免疫反應來促進病原防御,可以抵御細菌和病毒感染[24]。CLU 參與免疫調節、細胞黏附、細胞分化及轉化調控等多項重要的機體活動。AGP 是免疫調節蛋白,通過調節細胞因子參與免疫調節功能[25]。本研究對主要DEPs 的分析發現,娟犏牛奶中豐度較高的主要乳清和MFGM DEPs在免疫保護和免疫調節等方面表現優異,且娟犏牛奶乳清和MFGM 中上調的免疫相關蛋白數量和總豐度都高于牦牛奶,說明娟犏牛奶較牦牛奶在免疫保護和免疫調節等生物過程中發揮更重要的作用。
娟犏牛奶與牦牛奶相比,主要乳清和MFGM 高豐度下調DEPs 包括HSP7C、CD59、XDH 和CREG1 等。娟犏牛奶乳清與MFGM 中的下調核心DEPs 包括HSP7C、PIGR、RHOA和CD59等。對這些DEPs分析發現熱休克蛋白家族是一組對于維持細胞功能至關重要的分子伴侶,HSP7C是熱休克蛋白家族的組成成員,也是高度保守的伴侶蛋白[26],還具有促凋亡和抗凋亡的特性,對細胞活力起保護作用[27]。CD59 分子可參與免疫反應的調節過程,誘導T淋巴細胞的激活,并且促進T 淋巴細胞增殖[28]。XDH 能夠抑制金黃色葡萄球菌、大腸桿菌和沙門氏菌的生長[29],在胃腸道內發揮抗菌作用,具有免疫保護功能[30]。CREG1是維持組織細胞成熟穩態的重要分子,具有對抗細胞缺血缺氧損傷、抗凋亡和抗增殖等多種作用[31],CREG1 也可能在發揮對抗細胞/組織損傷,保證細胞穩定性方面具有十分重要的臨床藥用價值。本研究對主要DEPs 的分析發現娟犏牛奶下調的主要乳清和MFGM 高豐度DEPs在保證細胞穩定性、促進T 淋巴細胞增殖和抗菌等方面發揮作用,說明牦牛奶中豐度較高的主要乳清和MFGM 差異蛋白在保證細胞穩定性、促進T 淋巴細胞增殖和抗菌等方面表現優異。
4 結論
本試驗利用TMT 定量蛋白組學技術成功獲得了娟犏牛與牦牛的乳清蛋白、MFGM 蛋白的差異表達譜。研究發現,娟犏牛乳較牦牛乳在免疫保護和免疫調節等生物過程中發揮更重要作用,推測娟犏牛奶更有助于新生兒建立抗微生物感染的免疫系統。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
