細菌非編碼小RNA的生物學特征及作用機制研究進展
2023-06-21 09:20:39王海燕賀婷停李昊陽
四川師范大學學報(自然科學版) 2023年5期
王海燕 賀婷停 李昊陽
摘要:細菌非編碼小RNA(sRNA)是一類長度為50~500 nt、一般不編碼蛋白質的具有調控功能的小RNA,主要通過與靶mRNA堿基配對或與靶蛋白質結合,在轉錄后水平發揮調控作用.sRNA參與細菌氨基酸代謝和轉運、環境脅迫響應以及毒力作用等眾多重要生理過程的調控,與傳統的轉錄因子等構成多層級的調控網絡,協同作用使細菌對環境變化做出快速且精細的響應.由于sRNA在細菌調控網絡中的重要作用,發現新的sRNA并闡明它們的調控功能成為原核生物基因表達調控研究的一個熱點.本綜述對細菌sRNA的篩選、鑒定、作用機制以及生物學功能等方面取得的進展進行總結,希望給相關領域的研究者提供一個較為全面的視角.
關鍵詞:細菌; 非編碼sRNA; 轉錄后調控
中圖分類號:Q939.1 文獻標志碼: A 文章編號:1001-8395(2023)05-0581-10
細菌常常暴露在惡劣的自然環境中,為了應對外界環境的變化,細菌已進化形成一套精密的應答機制來適應各種環境變化.長期以來,普遍認為細菌基因的表達調控主要存在于轉錄水平,現在發現細菌中同樣存在由非編碼RNA介導的轉錄后調控.細菌非編碼RNA是一大類具有調控功能的RNA分子,通過各種機制調控多種生理過程,這些RNA包括核開關(riboswitch)、CRISPR RNA、非編碼小RNA(sRNA)等[1],其中sRNA是研究最多的一類轉錄后調控因子.細菌sRNA通常不進行翻譯(但是也有例外[2-3]),主要通過與被調控的靶mRNA進行堿基配對來調控靶標的翻譯或穩定性[4],少數sRNA通過與蛋白質結合發揮功能[5].和蛋白類調控因子相比,sRNA分子小,合成及周轉所需能量和時間少,sRNA結構可以在不同構象之間快速切換,確保了對環境壓力信號的迅速反應.因此,sRNA介導的轉錄后基因表達調控在細菌中扮演重要角色[6-7].
近年來,高通量測序特別是RNA-seq已經成為發現新sRNA的主流方法,研究者相繼從大腸桿菌、沙門氏菌、結核桿菌、霍亂弧菌、枯草芽孢桿菌、地衣芽孢桿菌、金黃色葡萄球菌、李斯特菌等多種細菌中預測到大量sRNA,每種細菌的sRNA數目約在幾十到200 個之間[8].然而,對sRNA的實驗鑒定及功能研究還比較有限,已有的研究主要集中于大腸桿菌和沙門氏菌等幾種革蘭氏陰性細菌,對于革蘭氏陽性菌,即使在模式菌枯草芽孢桿菌中經過功能鑒定的sRNA也只有十來個.細菌sRNA作為基因表達的轉錄后調控因子,參與細胞生理的多個層面,包括碳源、氨基酸和金屬離子的利用,細胞分裂,DNA水平轉移,群體感應,生物膜形成和毒力基因表達等,對細菌應對環境變化產生快速響應至關重要[9-13].本文對細菌sRNA的發現方法、功能機制以及研究策略進行簡要總結,以期能對細菌sRNA有一個更加全面和清晰的認識.
1細菌sRNA的分類
細菌sRNA長度多在50~500 nt,絕大部分不編碼蛋白質,轉錄后可形成穩定的二級結構,通過各種作用機制調控細菌的多種生理過程,影響遍及細菌的整個生命周期.目前已鑒定的細菌sRNA,根據其功能及作用方式可分為3大類:具有看家功能的sRNA,與靶蛋白結合的sRNA,與靶mRNA堿基配對結合的sRNA.已發現的細菌sRNA絕大多數屬于第三種類型,是sRNA發揮調控功能最普遍的一種方式,即調節mRNA的翻譯和穩定性.這類調控RNA根據其編碼基因的染色體位置進一步被分為兩類:順式編碼(cis-encoded)sRNA和反式編碼(trans-encoded)sRNA.順式編碼sRNA(又叫antisense sRNA)由其調控靶基因的反義鏈轉錄產生,通過與靶mRNA完全互補配對影響其穩定性.反式編碼sRNA通常由染色體的基因間隔區獨立轉錄產生,還有一部分由mRNA的5或3UTR加工產生,這類sRNA通過不完全堿基配對方式結合靶mRNA調控其穩定性或翻譯.雖然細菌中sRNA的數量并不多,但同一個sRNA可以調控多個靶基因,同一個靶基因也可以被多個sRNA所調控,使sRNA能夠參與對調控網絡不同節點、不同嚴謹性的協同控制.
2細菌sRNA的發現
由于sRNA是一類序列較短、通常不編碼蛋白的轉錄后調控因子,不受無義或移碼突變的影響,且很多只在特定環境下(如生長狀態、溫度、pH、離子濃度等)誘導表達,這給細菌sRNA的發現帶來一定的困難.
早期發現的sRNA主要是通過實驗分離得到,包括放射性標記法、鳥槍克隆法(RNA組學)以及免疫共沉淀法[14-17].這些方法主要適合高豐度、組成型sRNA的篩選,例如4.5S、tmRNA、RNase P以及Spot 42等[18-20].隨著高通量測序技術的發展及相應的生物信息學分析工具不斷開發,利用基因組或轉錄組測序數據,大量sRNA被預測出來[21],成為發現細菌sRNA的主流方法.生物信息學預測軟件主要是參考細菌sRNA的序列保守性、二級結構和熱動力學穩定性以及轉錄信號等特征,包括基于比較基因組學開發的QRNA、ERPIN、MSARI、INFERNAL、ISI,基于二級結構與熱穩定性開發的RNAZ和CARNAC,以及基于孤兒啟動子轉錄信號特征開發的sRNAscanner、sRNAPredict3/SIPHT等[22].相比于實驗方法,生物信息學方法通過對高通量測序數據的分析,可在短期內獲得大量sRNA的相關信息,具有成本低、時間快等優點.但該方法的一個缺點是預測具有一定的不可靠性,因此,需結合生物學實驗進行驗證,方法包括qRT-PCR和Northern雜交等,其中Northern雜交是驗證非編碼sRNA存在的金標準.
3sRNA靶基因的高通量篩選及實驗鑒定
細菌非編碼sRNA主要通過堿基互補配對的方式對靶基因mRNA進行調控,如何有效找出靶基因是研究sRNA功能的關鍵因素.由于sRNA與靶mRNA是不完全堿基配對,配對核心區域一般只有7個堿基左右,因此,靶基因的預測及篩選具有相當大的挑戰性.迄今為止,已發展多種方法來篩選sRNA所調控的靶基因,包括生物信息學預測、組學篩選、實驗與高通量測序相結合的方法等.
生物信息學軟件主要根據RNA序列保守性、sRNA-mRNA堿基配對及結合自由能、RNA二級結構等因素預測sRNA的靶基因,是最簡單、快速的靶基因預測方法.迄今為止,有TargetRNA2[23]、CopraRNA/IntaRNA[24]和sTarPicker[25]等軟件被開發用于預測sRNA的靶mRNA,不同軟件預測方式不同,預測出的靶基因有時差異較大.此外,由于大部分預測軟件都是基于大腸桿菌及沙門氏菌中已經鑒定的sRNA及靶基因進行學習的,因此,用于其他非模式菌以及革蘭氏陽性菌也可能出現較多假陽性結果,給后續的實驗驗證造成干擾.
轉錄組測序(RNA-seq)是目前使用最多的靶基因篩選方法,其一般流程是:1) 在宿主菌中轉入一個誘導型sRNA表達質粒,對照組轉入不含sRNA的空載體;2) 在細菌生長對數期誘導sRNA表達,短暫誘導后收集菌體提取總RNA;3) 將sRNA誘導表達菌株和對照菌株的RNA進行轉錄組測序,比較找出誘導后上調或下調的基因,即為候選靶基因.不過,這種方法篩選到的是sRNA誘導表達后的差異基因,但無法區分這些差異基因來自于sRNA的直接調控還是間接調控.一般來說,采用較短的誘導時間(15~30 min)可以盡可能減少間接調控基因的RNA水平變化.此外,RNA-seq只能檢測mRNA水平發生變化的靶基因,無法獲得翻譯水平受到影響的靶基因.因此,該方法仍然具有一定的局限性.
實驗與高通量測序相結合篩選靶基因的方法主要分為兩大類.一類是基于革蘭氏陰性菌中sRNA-mRNA結合通常需要伴侶蛋白(如RNA結合蛋白Hfq和ProQ)輔助、或結合后會招募核酸酶(如RNase E)參與mRNA降解等特性,通過免疫沉淀特定蛋白來捕獲sRNA-mRNA-蛋白復合體,再用RNA連接酶將配對的sRNA-mRNA首尾相連,最后對連接RNA進行高通量測序.RIL-seq(RNA interaction by ligation and sequencing)[26]、CLASH(UV-crosslinking, ligation and sequencing of hybrids)等就是基于該原理設計的靶基因篩選策略,通過測序數據分析,理論上可以捕獲細胞特定時期天然狀態下所有sRNA的體內靶標.另一類是基于革蘭氏陽性菌中sRNA-mRNA結合無固定結合蛋白這一特點,主要方法如MAPS(MS2-affinity purification coupled with RNA-Seq)[27].MAPS的原理是在sRNA上融合一段來自于噬菌體的RNA序列MS2,然后利用可結合MS2 RNA的蛋白進行免疫共沉淀捕獲sRNA及其結合的靶mRNA或靶蛋白,最后通過RNA-seq或蛋白質譜分析獲得靶基因.
這些通過高通量測序篩選獲得的靶基因,還需要進行體外、體內相互作用的實驗驗證,以確認其可靠性.體外相互作用的檢測方法主要包括凝膠阻滯實驗EMSA、足跡法footprinting[28]以及趾紋法toeprinting[29]等,用于檢測RNA-RNA以及RNA-蛋白質的相互作用.體內相互作用實驗主要是利用報告基因(GFP或lacZ)來檢測sRNA-mRNA的相互作用,是驗證sRNA與靶mRNA相互作用最常用的方法.首先利用copraRNA/IntaRNA軟件預測二者的相互作用位點,由于大部分sRNA與靶mRNA的結合位于靶mRNA的5UTR或起始密碼子附近的區域,因此,可以構建靶基因5UTR及起始密碼子附近區域與報告基因編碼區融合的表達質粒,sRNA誘導表達單元可以與報告基因構建在同一個載體上,也可以構建在不同的載體上.將含有sRNA以及融合報告基因的質粒轉入同一個細胞,通過sRNA誘導前后報告基因表達水平的變化來判斷sRNA是否調控該mRNA.體內相互作用實驗不僅可以驗證sRNA對靶基因的調控,還可以通過堿基突變方式尋找sRNA-mRNA的結合位點,因此成為檢驗sRNA-mRNA相互作用最可靠的方法.
4細菌sRNA的作用機制及生物學功能
細菌sRNA的表達通常受不同生長時期或各種脅迫因素的誘導,sRNA水平在某種特定條件下達到最高.當sRNA被誘導產生后,可通過不同方式調控靶基因的表達.通常情況下,少數sRNA直接與蛋白質結合發揮調控功能,大多數sRNA通過堿基配對與靶基因mRNA結合對其進行調控,最新的研究顯示,sRNA還可以通過結合其他sRNA發揮調控功能.
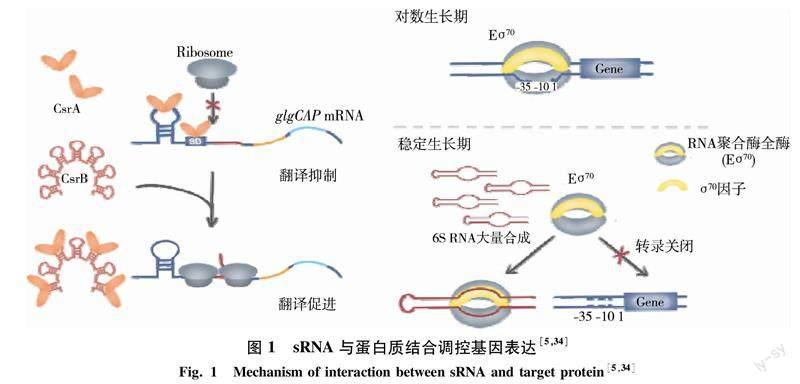
4.1sRNA與蛋白質結合 這類sRNA在體內通過模擬靶蛋白的天然底物結構,與底物競爭性結合靶蛋白從而影響靶蛋白的功能.例如大腸桿菌全局性調控蛋白CsrA是一種RNA結合蛋白,可結合在mRNA分子上進而影響其穩定性或翻譯,CsrA的結合嚴格依賴于RNA上的GGA基序.非編碼sRNA CsrB上含有多個GGA基序可同時結合超過18個CsrA,從而與mRNA競爭性結合CsrA,釋放CsrA對靶mRNA的調控.例如大腸桿菌glgCAP操縱子參與糖原合成,CsrA與glgC的SD序列結合,抑制glgC mRNA的翻譯并加速glgCAP mRNA降解,導致糖原合成減少.當CsrB大量表達時,CsrB螯合CsrA釋放glgCAP mRNA的核糖體結合位點,進而促進mRNA翻譯,使糖原合成增加[5,30](圖1).與CsrB和CsrA類似的sRNA和RNA結合蛋白廣泛存在于多種細菌的基因組中,例如熒光假單胞菌有3個CsrB類似sRNA(RsmX/Y/Z)和兩個CsrA類似蛋白(RsmA/E),表明這種sRNA與蛋白結合的調控機制在細菌中是常見的[31].此外,6S RNA也是一個研究比較清楚的與蛋白結合的sRNA,其二級結構與基因轉錄起始區的DNA構象類似,可以競爭結合σ70 RNA聚合酶(Eσ70),從而影響相關基因轉錄[32-34].大腸桿菌的Eσ70主要負責對數生長期相關基因的轉錄,當細菌生長進入穩定期,6S RNA大量產生并螯合Eσ70,使依賴于Eσ70的對數期基因轉錄關閉,而依賴于其他σ因子的穩定期或脅迫基因轉錄得以開啟(圖1).6S RNA在原核生物中高度保守,枯草芽孢桿菌有兩個6S RNA同源RNA,分別為6S-1和6S-2[35-36],二者均可以與RNA聚合酶結合抑制下游基因的轉錄.
4.2sRNA與mRNA結合 順式編碼sRNA通常由蛋白基因的反義鏈轉錄產生,大多是通過高通量測序被發現的,研究最多的是細菌的毒素-抗毒素系統.細菌毒素蛋白基因反義鏈編碼的抗毒素實質是一個非編碼sRNA,抗毒素sRNA與毒素mRNA完全配對,影響mRNA的穩定性或翻譯,從而中和毒素[37].例如大腸桿菌I型毒素-抗毒素系統TisAB/IstR-1,在正常生長條件下,抗毒素sRNA IstR-1(70 nt)表達,通過其5端的25個堿基與毒蛋白tisAB mRNA的5UTR互補配對,抑制翻譯并招募RNase III加速其降解,使毒蛋白低表達細菌得以正常生長.在SOS應激條件下,IstR-1表達水平下降,tisAB mRNA持續翻譯促進毒蛋白積累,導致細菌生長減緩[38-39].枯草芽孢桿菌染色體上的原噬菌體區域編碼的I型毒素-抗毒素系統BsrG/SR4也具有類似的調控機制[40].除了毒素-抗毒素系統外,地衣芽孢桿菌的枯草菌素家族蛋白酶基因apr的反義鏈可以轉錄產生sRNA AprA,它與蛋白酶基因mRNA相互作用抑制其翻譯,從而降低蛋白酶水平[41].
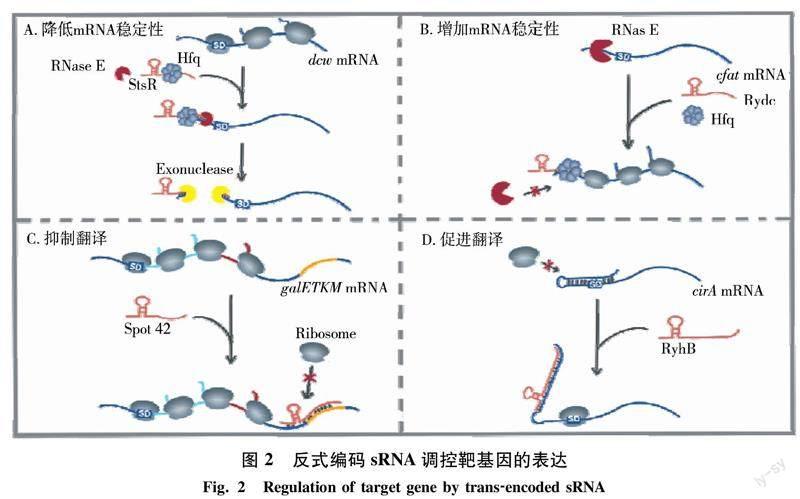
反式編碼sRNA在細菌中數量最多,通過部分堿基配對調控靶mRNA的表達,其作用機制主要包括以下4種方式(圖2):1)sRNA與mRNA結合后會招募核糖核酸酶對sRNA-mRNA復合體進行降解,促進靶mRNA的降解.例如在α變形桿菌中,細胞生長到穩定期以及處于脅迫條件時,sRNA StsR高表達,然后與細胞分裂/細胞壁基因簇dcw mRNA的5UTR結合,招募內切核酸酶RNase E切割dcw mRNA,進而加速外切核酸酶對mRNA的降解,最終導致細胞分裂速度減慢[42].2)sRNA結合封閉靶mRNA的核酸酶切割位點,增加mRNA的穩定性.如在腸沙門氏菌中,細菌通過改變細胞膜性質來抵抗外界環境的變化,基因cfa1(編碼環丙烷脂肪酸合酶)參與細胞膜重塑過程.在正常生長過程中,sRNA RydC可與Hfq共同作用結合在cfa1 mRNA的RNase E識別位點,保護cfa1 mRNA免遭RNase E切割,從而維持細胞在生長過程中細胞膜的特性[43].3)sRNA結合于靶mRNA的核糖體結合位點及附近,阻止核糖體與mRNA結合,從而抑制靶mRNA的翻譯.該方式是sRNA最主要的作用方式,也是生物信息學預測sRNA靶基因的一個重要參數.如大腸桿菌的sRNA Spot 42,長度為109 nt,特異性結合在操縱子galETKM mRNA的galK(編碼半乳糖激酶)的翻譯起始區域,阻斷核糖體結合抑制galK翻譯,使半乳糖激酶與UDP-半乳糖差向異構酶(由galE編碼)的比率發生改變,從而適應不同的生存環境[44].4)sRNA與mRNA的5UTR結合,改變5UTR區的構象使核糖體結合位點得以暴露,促進靶基因的翻譯.如在鐵離子不足的條件下,大腸桿菌sRNA RyhB高表達,與大腸桿菌素(colicin)受體基因cirA mRNA的5UTR結合暴露核糖體結合位點,促進cirA mRNA的翻譯,使細菌對大腸桿菌素更加敏感[45].
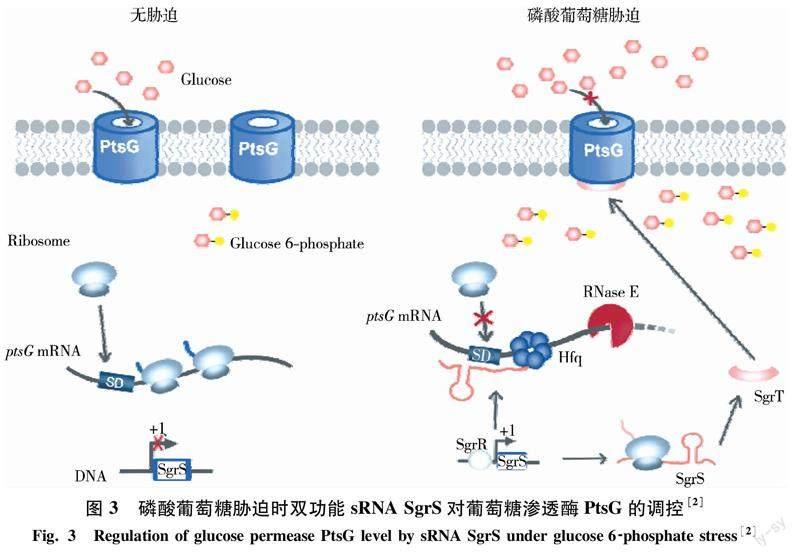
4.3雙功能sRNA 傳統意義上,sRNA為不編碼蛋白的非編碼RNA,但隨著研究的深入,發現個別sRNA也可以作為mRNA編碼小肽行使功能.迄今為止,共發現5個雙功能sRNA,即大腸桿菌的SgrS[46]、金黃色葡萄球菌的RNA III[47]和Psm-mec[48]、膿性鏈球菌的Pel RNA[49]以及枯草芽孢桿菌的SR1[50],其中SgrS的特性及功能在大腸桿菌和沙門氏菌中研究得較清楚.細菌利用細胞膜上的PTS系統將葡萄糖運輸到胞內,同時磷酸化生成6-磷酸葡萄糖(G6P),當糖的轉運和代謝不能協調匹配,就可能導致胞內G6P積累,產生細胞毒性.在G6P脅迫下,轉錄因子SgrR大量產生并激活sRNA SgrS的轉錄,SgrS可與PTS系統的葡萄糖滲透酶基因ptsG mRNA的SD序列結合阻止核糖體結合,并招募RNase E加速mRNA降解,從而抑制新的PtsG蛋白合成.此外,sRNA SgrS還編碼一個長度僅為43個氨基酸的小肽SgrT,SgrT可與細胞膜上已經存在的PtsG蛋白結合抑制其轉運活性[2](圖3).SgrS的這兩個功能似乎是互斥的,即一個SgrS分子只能以一種方式發揮作用,表現出對G6P脅迫響應的時間順序:即SgrS首先以sRNA形式與靶mRNA配對快速響應脅迫,而SgrT蛋白要在脅迫后30 min才產生,進一步發揮抑制PtsG蛋白活性的功能.
4.4作為sRNA“海綿”間接調控靶基因的表達 sRNA除了結合靶蛋白和靶mRNA,還可以作為RNA“海綿”,通過結合具有調控功能的sRNA,間接調控靶mRNA的表達.例如沙門氏菌sRNA SroC,由Glu/Asp轉運蛋白操縱子gltIJKL的gltI mRNA的3端加工產生,可以競爭結合具有全局性調控功能的sRNA GcvB,形成sRNA-sRNA配對,進而招募RNase E對GcvB進行降解,以緩解GcvB對氨基酸轉運及代謝相關基因(也包括gltIJKL mRNA)的抑制作用[51](圖4).此外,SroC還可以結合sRNA MgrR,釋放其對eptB mRNA的抑制作用,影響細菌對多黏菌素B的敏感性[52].枯草芽孢桿菌中一個獨立轉錄產生的sRNA RosA也可作為RNA海綿,結合至少兩種sRNA,即參與氧化脅迫調節的RoxS以及參與鐵脅迫調節的FsrA.RosA的轉錄受到碳代謝關鍵調控因子CcpA的抑制,在以阿拉伯糖為碳源時,CcpA表達受到抑制,使RosA的轉錄增加,RosA競爭性結合RoxS,從而緩解RoxS對TCA循環以及細胞呼吸鏈相關基因的抑制作用,由此將RoxS對氧化脅迫的調控作用與碳代謝又聯系起來[53].
4.5sRNA參與多個代謝通路的協同控制 反式編碼sRNA通過短的不完全堿基配對與靶mRNA結合,因此,同一個sRNA可以調節相關通路的多個基因,協同控制相應的生理過程.sRNA參與細菌鐵穩態調控是研究得比較深入的案例.鐵元素是絕大多數生命必需的營養物質,通常以蛋白輔因子形式存在,大腸桿菌一般擁有上百種含鐵蛋白,包括代謝酶、電子傳遞蛋白、感應與調控蛋白等,參與一系列重要的生物過程.但鐵過量又會給生命帶來嚴重威脅,因此,細菌必須隨時維持細胞內鐵元素水平的相對穩定,即鐵穩態.轉錄調控因子Fur是最關鍵的鐵穩態調控因子,此外,還有多個sRNA參與細菌鐵穩態的調控,最著名的是sRNA RyhB.研究發現,當胞內游離Fe2+濃度較低時,RyhB參與調控100多個基因的表達以維持胞內鐵離子穩態:1) 抑制非必需含Fe2+蛋白的表達,包括約50個Fe-S蛋白基因以及21個Fe2+結合蛋白基因,以使細胞內有限的鐵用于必需的生命過程;2) 抑制Fe-S簇合成的“管家”Isc系統,促進脅迫響應的Suf系統,節約鐵離子以滿足最需要的Fe-S蛋白合成;3) 促進鐵攝取相關蛋白CirA以及ShiA的表達,增加細胞外鐵離子的輸入[54].
另一方面,同一個靶mRNA又可被不同的sRNA所調節,使細胞內不同的靶mRNA通過sRNA連接成復雜的網絡以協調細菌的生理和行為.在大腸桿菌和其他腸道細菌中,生物膜形成涉及對鞭毛合成和生物膜基質合成兩個過程的反向控制,不同sigma因子、一系列級聯調控轉錄因子以及大量sRNA協同作用共同調控這一過程.其中,開啟鞭毛基因表達最上層的轉錄激活因子FlhDC,負責生長穩定期基因轉錄的sigma因子σS,以及生物膜基質重要組分菌毛合成的關鍵調節因子CsgD,這3個基因的mRNA被證明是多個sRNA的作用中心.研究顯示,flhDC mRNA可被3種sRNA ArcZ、OmrA/OmrB以及OxyS結合而抑制表達,被McaS結合則促進翻譯.σSmRNA可被sRNA DsrA、RprA 和ArcZ正調控,促進菌毛及生物膜形成.csgD mRNA 可被sRNA McaS、RprA、GcvB、RydC、RybB和OmrA/OmrB負調控,抑制菌毛及生物膜形成[55](圖5).綜合來看,這些sRNA根據其對鞭毛及生物膜合成的調控效應可分為4類:1) 促進生物膜形成、下調鞭毛合成(如ArcZ)[56-57];2) 作用正好相反,即刺激鞭毛合成、干擾生物膜基質合成(如McaS)[58];3) 下調這兩種功能(如OmrA/OmrB)[59];4) 在鞭毛和生物膜功能的控制中具有多種更復雜的影響(如RprA)[60].不同sRNA共同作用的最終效應取決于sRNA和mRNA的表達時期、速率與水平、sRNA與mRNA結合力等多種因素[55].
5小結與展望
非編碼sRNA在細菌中廣泛存在,其調控作用遍及細菌的整個生命過程.在細菌基因表達調控系統中,sRNA與傳統的轉錄因子等構成多層級的調控網絡,協同作用使細菌對環境脅迫做出快速且精細的響應.目前,細菌sRNA的研究主要集中在大腸桿菌和沙門氏菌等革蘭氏陰性細菌,對革蘭氏陽性菌及非模式菌的sRNA研究還比較缺乏.對在實際應用中具有重要價值的非模式菌(如致病菌、工業生產菌等)開展研究,探索sRNA如何調控細菌的致病性、耐藥性及脅迫應答,不僅可以揭示更多參與不同生物學功能調控的sRNA,擴展sRNA的多樣性,也將為其實際應用提供理論基礎.
參考文獻
[1] WATERS L S, STORZ G. Regulatory RNAs in bacteria[J]. Cell,2009,136(4):615-628.
[2] RAINA M, KING A, BIANCO C, et al. Dual-function RNAs[J]. Microbiol Spectr,2018,6(5):471-485.
[3] VANDERPOOL C K, BALASUBRAMANIAN D, LLOYD C R. Dual-function RNA regulators in bacteria[J]. Biochimie,2011,93(11):1943-1949.
[4] H?R J, MATERA G, VOGEL J, et al. Trans-acting small RNAs and their effects on gene expression in Escherichia coli and Salmonella enterica[J/OL]. EcoSal Plus,2020,9(1):10.1128/ecosalplus.ESP-0030-2019.
[5] BAKER C S, MOROZOV I, SUZUKI K, et al. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli[J]. Mol Microbiol,2002,44(6):1599-1610.
[6] MARS R A, NICOLAS P, DENHAM E L, et al. Regulatory RNAs in Bacillus subtilis:a gram-positive perspective on bacterial RNA-mediated regulation of gene expression[J]. Microbiol Mol Biol Rev,2016,80(4):1029-1057.
[7] GOTTESMAN S, STORZ G. Bacterial small RNA regulators:versatile roles and rapidly evolving variations[J]. Cold Spring Harb Perspect Biol,2011,3(12):a003798.
[8] H?R J, GORSKI S A, VOGEL J. Bacterial RNA biology on a genome scale[J]. Mol Cell,2018,70(5):785-799.
[9] MORFELDT E, TAYLOR D, GABAIN A V, et al. Activation of alpha-toxin translation in Staphylococcus aureus by the trans-encoded antisense RNA, RNAIII[J]. Embo J,1995,14(18):4569-4577.
[10] ROMBY P, VANDENESCH F, WAGNER E G. The role of RNAs in the regulation of virulence-gene expression[J]. Curr Opin Microbiol,2006,9(2):229-236.
[11] PAPENFORT K, VOGEL J. Regulatory RNA in bacterial pathogens[J]. Cell Host Microbe,2010,8(1):116-127.
[12] DURAND S, GILET L, CONDON C. The essential function of B. subtilis RNase III is to silence foreign toxin genes[J]. PLoS Genet,2012,8(12):e1003181.
[13] HOE C H, RAABE C A, ROZHDESTVENSKY T S, et al. Bacterial sRNAs:regulation in stress[J]. Int J Med Microbiol,2013,303(5):217-229.
[14] H?TTENHOFER A, VOGEL J. Experimental approaches to identify non-coding RNAs[J]. Nucleic Acids Res,2006,34(2):635-646.
[15] LIVNY J, WALDOR M K. Identification of small RNAs in diverse bacterial species[J]. Curr Opin Microbiol,2007,10(2):96-101.
[16] VOGEL J, GERHART E, WAGNER H. Approaches to Identify Novel Non-messenger RNAs in Bacteria and to Investigate Their Biological Functions:RNA Mining[M]//Handbook of RNA Biochemistry. Weinheim:Wiley-VCH Verlag GmbH,2008:595-613.
[17] BILUSIC I, POPITSCH N, RESCHENEDER P, et al. Revisiting the coding potential of the E. coli genome through Hfq co-immunoprecipitation[J]. RNA Biol,2014,11(5):641-654.
[18] GRIFFIN B E. Separation of 32P-labelled ribonucleic acid components. The use of polyethylenimine-cellulose (TLC) as a second dimension in separating oligoribonucleotides of ‘4.5 S and 5 S from E. coli[J]. FEBS Letters,1971,15(3):165-168.
[19] HINDLEY J. Fractionation of 32P-labelled ribonucleic acids on polyacrylamide gels and their characterization by fingerprinting[J]. Journal of Molecular Biology,1967,30(1):125-136.
[20] IKEMURA T, DAHLBERG J E. Small ribonucleic acids of Escherichia coli:I. Characterization by polyacrylamide gel electrophoresis and fingerprint analysis[J]. Journal of Biological Chemistry,1973,248(14):5024-5032.
[21] WASSARMAN K M, REPOILA F, ROSENOW C, et al. Identification of novel small RNAs using comparative genomics and microarrays[J]. Genes Dev,2001,15(13):1637-1651.
[22] SRIDHAR J, GUNASEKARAN P. Computational small RNA prediction in bacteria[J]. Bioinform Biol Insights,2013,7(7):83-95.
[23] KERY M B, FELDMAN M, LIVNY J, et al. TargetRNA2:identifying targets of small regulatory RNAs in bacteria[J]. Nucleic Acids Res,2014,42(1):W124-129.
[24] WRIGHT P R, GEORG J, MANN M, et al. CopraRNA and IntaRNA:predicting small RNA targets, networks and interaction domains[J]. Nucleic Acids Res,2014,42(1):W119-123.
[25] YING X M, CAO Y, WU J Y, et al. sTarPicker: a method for efficient prediction of bacterial sRNA targets based on a two-step model for hybridization[J]. Plos One,2011,6(7):e22705.
[26] MELAMED S, FAIGENBAUM-ROMM R, PEER A, et al. Mapping the small RNA interactome in bacteria using RIL-seq[J]. Nat Protoc,2018,13(1):1-33.
[27] MERCIER N, PR?VOST K, MASS?E, et al. MS2-affinity purification coupled with RNA sequencing in gram-positive bacteria[J]. J Vis Exp,2021,168(6):1-13.
[28] PENG Y, SOPER T J, WOODSON S A. RNase footprinting of protein binding sites on an mRNA target of small RNAs[J]. Methods Mol Biol,2012,905:213-224.
[29] NILSEN T W. Toeprinting [J]. Cold Spring Harb Protoc,2013,2013(9):896-899.
[30] LIU M Y, GUI G J, WEI B D, et al. The RNA molecule CsrB binds to the global regulatory protein CsrA and antagonizes its activity in Escherichia coli[J]. J Biol Chem,1997,272(28):17502-17510.
[31] SONNLEITNER E, HAAS D. Small RNAs as regulators of primary and secondary metabolism in Pseudomonas species[J]. Appl Microbiol Biotechnol,2011,91(1):63-79.
[32] WASSARMAN K M. 6S RNA:a regulator of transcription[J]. Mol Microbiol,2007,65(6):1425-1431.
[33] WILLKOMM D K, HARTMANN R K. 6S RNA-an ancient regulator of bacterial RNA polymerase rediscovered[J]. Biol Chem,2005,386(12):1273-1277.
[34] WASSARMAN K M. 6S RNA, a global regulator of transcription[J/OL]. Microbiol Spectr,2018,6(3):10.1128/microbiolspec.RWR-0019-2018.
[35] TROTOCHAUD A E, WASSARMAN K M. A highly conserved 6S RNA structure is required for regulation of transcription[J]. Nat Struct Mol Biol,2005,12(4):313-319.
[36] BARRICK J E, SUDARSAN N, WEINBERG Z, et al. 6S RNA is a widespread regulator of eubacterial RNA polymerase that resembles an open promoter[J]. RNA,2005,11(5):774-784.
[37] SMIRNOV A, F?RSTNER K U, HOLMQVIST E, et al. Grad-seq guides the discovery of ProQ as a major small RNA-binding protein[J]. Proc Natl Acad Sci USA,2016,113(41):11591-11596.
[38] DARFEUILLE F, UNOSON C, VOGEL J, et al. An antisense RNA inhibits translation by competing with standby ribosomes[J]. Mol Cell,2007,26(3):381-392.
[39] VOGEL J, ARGAMAN L, WAGNER E G, et al. The small RNA IstR inhibits synthesis of an SOS-induced toxic peptide[J]. Curr Biol,2004,14(24):2271-2276.
[40] JAHN N, PREIS H, WIEDEMANN C, et al. BsrG/SR4 from Bacillus subtilis-the first temperature-dependent type I toxin-antitoxin system[J]. Mol Microbiol,2012,83(3):579-598.
[41] HERTEL R, MEYERJ?RGENS S, VOIGT B, et al. Small RNA mediated repression of subtilisin production in Bacillus licheniformis[J]. Sci Rep,2017,7(1):5699.
[42] GR?TZNER J, REMES B, EISENHARDT K M H, et al. sRNA-mediated RNA processing regulates bacterial cell division[J]. Nucleic Acids Res,2021,49(12):7035-7052.
[43] FR?HLICH K S, PAPENFORT K, FEKETE A, et al. A small RNA activates CFA synthase by isoform-specific mRNA stabilization[J]. Embo J,2013,32(22):2963-2979.
[44] M?LLER T, FRANCH T, UDESEN C, et al. Spot 42 RNA mediates discoordinate expression of the E. coli galactose operon[J]. Genes Dev,2002,16(13):1696-1706.
[45] SALVAIL H, CARON M P, B?LANGER J, et al. Antagonistic functions between the RNA chaperone Hfq and an sRNA regulate sensitivity to the antibiotic colicin[J]. EMBO J,2013,32(20):2764-2778.
[46] WADLER C S, VANDERPOOL C K. A dual function for a bacterial small RNA:SgrS performs base pairing-dependent regulation and encodes a functional polypeptide[J]. Proc Natl Acad Sci USA,2007,104(51):20454-20459.
[47] BRONESKY D, WU Z, MARZI S, et al. Staphylococcus aureus RNAIII and its regulon link quorum sensing, stress responses, metabolic adaptation, and regulation of virulence gene expression[J]. Annu Rev Microbiol,2016,70(1):299-316.
[48] KAITO C, SAITO Y, IKUO M, et al. Mobile genetic element SCCmec-encoded psm-mec RNA suppresses translation of agrA and attenuates MRSA virulence[J]. PLoS Pathog,2013,9(4):e1003269.
[49] MANGOLD M, SILLER M, ROPPENSER B, et al. Synthesis of group A streptococcal virulence factors is controlled by a regulatory RNA molecule[J]. Mol Microbiol,2004,53(5):1515-1527.
[50] GIMPEL M, BRANTL S. Dual-function sRNA encoded peptide SR1P modulates moonlighting activity of B. subtilis GapA[J]. RNA Biol,2016,13(9):916-926.
[51] MIYAKOSHI M, CHAO Y, VOGEL J. Cross talk between ABC transporter mRNAs via a target mRNA-derived sponge of the GcvB small RNA[J]. EMBO J,2015,34(11):1478-1492.
[52] ACU?A L G, BARROS M J, PE?ALOZA D, et al. A feed-forward loop between SroC and MgrR small RNAs modulates the expression of eptB and the susceptibility to polymyxin B in Salmonella Typhimurium[J]. Microbiology,2016,162(11):1996-2004.
[53] DURAND S, CALLAN-SIDAT A, MCKEOWN J, et al. Identification of an RNA sponge that controls the RoxS riboregulator of central metabolism in Bacillus subtilis[J]. Nucleic Acids Res,2021,49(11):6399-6419.
[54] CHAREYRE S, MANDIN P. Bacterial iron homeostasis regulation by sRNAs[J]. Microbiol Spectr,2018,6(2):1-11
[55] MIKA F, HENGGE R. Small regulatory RNAs in the control of motility and biofilm formation in E. coli and Salmonella[J]. Int J Mol Sci,2013,14(3):4560-4579.
[56] DE LAY N, GOTTESMAN S. A complex network of small non-coding RNAs regulate motility in Escherichia coli[J]. Mol Microbiol,2012,86(3):524-538.
[57] MANDIN P, GOTTESMAN S. Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA[J]. Embo J,2010,29(18):3094-3107.
[58] J?RGENSEN M G, NIELSEN J S, BOYSEN A, et al. Small regulatory RNAs control the multi-cellular adhesive lifestyle of Escherichia coli[J]. Mol Microbiol,2012,84(1):36-50.
[59] HOLMQVIST E, REIMEGRD J, STERK M, et al. Two antisense RNAs target the transcriptional regulator CsgD to inhibit curli synthesis[J]. Embo J,2010,29(11):1840-1850.
[60] MIKA F, BUSSE S, POSSLING A, et al. Targeting of csgD by the small regulatory RNA RprA links stationary phase, biofilm formation and cell envelope stress in Escherichia coli[J]. Mol Microbiol,2012,84(1):51-65.
Biological Characteristics and Regulation Mechanisms
of Bacterial Non-coding Small RNAWANG Haiyan,HE Tingting,LI Haoyang(College of Life Sciences, Sichuan University, Chengdu 610064, Sichuan)
AbstractBacterial non-coding small RNAs (sRNA) exist as relatively short transcripts (50-500 nt) that are transcribed but do not encode proteins. sRNA plays a post-transcriptional regulatory role on target genes mostly by imperfect base-pairing with target mRNA, and rarely by binding target protein. sRNA is involved in the regulation of many important physiological processes, such as amino acid metabolism and transport, stress response and virulence, etc., forming a multi-level regulatory network with traditional transcription factors to enable bacteria responding quickly to environmental stress. Due to the important role of sRNA in bacterial regulatory network, the discovery of new sRNAs and revealing their regulatory mechanisms have become a hotspot in prokaryotic gene expression regulation research. This review summarizes the advance in screening, identification, regulatory mechanism and biological function of bacterial sRNA, hoping to provide a more comprehensive perspective for researchers of related fields.
Keywords:bacteria; noncoding sRNA; post-transcriptional regulation
(編輯 鄭月蓉)
